

 English
English文献解读|Nature(48.5):创伤后应激障碍患者脑单细胞转录组和染色质动力学
✦ +
+
论文ID
原名:Single-cell transcriptomic and chromatin dynamics of the human brain in PTSD
译名:创伤后应激障碍患者脑单细胞转录组和染色质动力学
期刊:Nature
影响因子:48.5
发表时间:2025.06.18
DOI号:10.1038/s41586-025-09083-y
背 景
创伤后应激障碍 (PTSD) 是一种常见的精神健康障碍,发生在严重创伤之后。PTSD 在普通人群中的发病率约为 6-8%,具有中等遗传性(从 24% 到 40% 不等)并且是多基因的。最近有研究开始阐明 PTSD 患者死后大脑的分子生物学。这些研究主要集中在前额皮质 (PFC)和杏仁核,并发现了几种基因通路的分子变化,包括 GABA 能信号传导、免疫和神经炎症以及糖皮质激素信号传导。鉴于受影响的生物过程的高度多样性,某种特定的细胞类型不太可能导致 PTSD 的病理生理。人类大脑中的基因表达由顺式调控元件 (CRE)、染色质修饰和转录因子与基因增强子和启动子的结合共同调控。CRE 在特定细胞类型中的作用对于理解疾病风险变异如何调控基因表达至关重要。先前的研究已经发现了 PTSD 患者 DNA 甲基化的显著变化;然而,目前还没有研究全面分析 PTSD 患者大脑的单细胞染色质景观。基因组技术的最新进展现在可以在单个细胞中检测染色质组装,这些检测可以与基因表达分析相结合,以提供所需的分辨率,以确定风险变异如何调控单个细胞内的转录。
实验设计
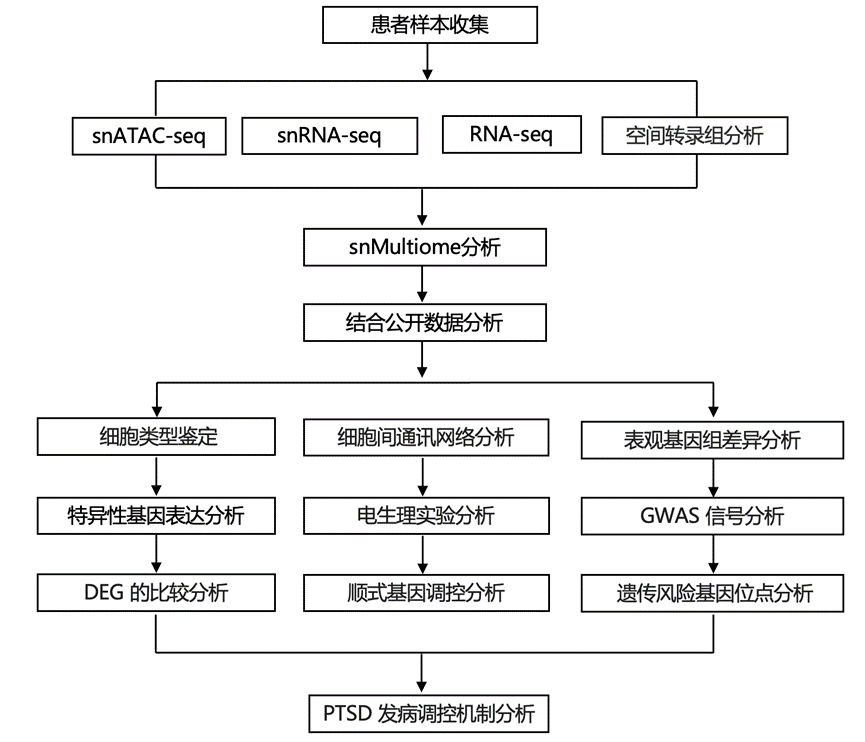
结 果
01

PTSD 患者 PFC 的单细胞多组学分析
为了分析PTSD中的单细胞类型基因表达和调控变化(图1a),研究团队对 111 名捐献者的背外侧前额叶皮层(DLPFC)进行了单细胞核转录组分析(snRNA-seq)和单细胞染色质转座酶可及性测序(snATAC-seq),对 36 名患有 PTSD 的捐献者、36 名患有 MDD(精神疾病对照)的捐献者以及 39 名均无 PTSD 的捐献者(CON)进行了分析,报告了 935371 个细胞核的 snRNA-seq 结果(图1b)、473033 个细胞核的 snATAC-seq 结果(图1c)以及 119431 个细胞核的单核多组学(snMultiome)结果(图1d)。
他们鉴定了兴奋性神经元(EXN;四种亚型:CUX2、RORB、FEZF2和OPRK1)、抑制性神经元(IN;五种亚型:LAMP5、KCNG1、VIP、SST和PVALB)、少突胶质细胞(OLG)、少突胶质细胞祖细胞(OPC)、内皮细胞(END)、星形胶质细胞(AST)和小胶质细胞(MG)(图1b),它们具有细胞类型特异性标记基因的表达谱。基于跨模态分离的细胞核数量,分析细胞类型的差异丰度,发现 PTSD 中 OLG 的数量与 CON 和 MDD 相比有所减少(图1b)。他们使用 snATAC-seq数据以及每个标记的染色质可及性概况和 snMultiome数据鉴定了所有七种典型脑细胞类型。为了鉴定更具体的转录组细胞亚型,他们使用之前的注释18鉴定了 61 种亚型:18 种 EXN 和 30 种 IN 和 13 种非神经元亚型,包括两种 OLG、三种 AST、巨噬细胞、T 细胞(T SKAP1 CD247)、平滑肌细胞和周细胞。
他们使用两种常见的基于细胞的差异表达基因(DEG)分析方法对所有 14 种细胞类型聚类进行了研究。此外,他们使用了基因表达的混合线性模型,该模型可以校正基于样本的偏差。他们将 snDEG 定义为 MAST 和 Wilcoxon 之间重叠的显著DEG。他们分析了典型的细胞类型和亚型(图1e),在 EXN 中发现了 322 个 DEG,在 IN 中发现了 66 个,其中最多(557 个)出现在CUX2 EXN 中。在七种细胞类型中,他们从MAST中鉴定出3431个DEG,从Wilcoxon中鉴定出2989个DEG。其中,1376个DEG同向重叠,包含1184个独特基因(PTSD snDEG)(图1f)。他们发现9.92%的DEG在EXN和IN中均发生显著改变(图1f),并且没有DEG在所有细胞类型中均受到调控。
对来自部分重叠队列的组织转录组分析(RNA-seq)显示,排名靠前的 PTSD snDEG 的方向和幅度一致(图1g),包括之前发现的基因FKBP5、YBX3和HDAC9。整体而言,他们发现两个数据集之间存在高度相关性,他们还鉴定出了许多新的亚型特异性基因,包括LAMP5 IN中的GRM5和CUX2以及OPRK1 EXN中的CLU,这些基因之前已证明与自杀和阿尔茨海默病有关。他们发现END和 OLG中FKBP5表达增加(图1g)。据报道, FKBP5转录本在 PTSD的死后额叶皮质中上调,并且与蘑菇型树突棘密度呈负相关。他们将 DEG 结果与部分重叠的较小数据集进行了比较,发现显著的 DEG与PTSD(61.4%)和 MDD(72.7%)队列在不同细胞类型中存在中度重叠。
他们纳入了一个精神病对照组,该组患者的一种疾病 (MDD) 在 PTSD 确诊患者中高度共病(超过 50%)。他们系统地比较了 PTSD 与 CON 以及 MDD 与 CON 中上调和下调的基因,以确定共同和不同的分子机制。他们以与 PTSD 队列相同的方式分析了 MDD 队列,并确定了 1918 个 MDD snDEG。总体而言,发现 MDD 和 PTSD 之间的 snDEG 具有高度重叠(57.6%)(图1h),其中 502 个是 PTSD 特异性的 snDEG,1236 个是 MDD 特异性的 snDEG。对PTSD特异性snDEG的分析显示,与钙调蛋白信号传导、钙粘蛋白和泛素结合相关的通路富集(图1i)。他们发现snRNA-seq与MDD数据集之间的整体转录水平具有高度相关性,且前几个MDD DEG之间存在中等程度的相关性,他们观察到非神经元细胞类型中MDD和PTSD之间的DEG重叠程度最高。
为了表征 PTSD 和 MDD 之间的差异,他们通过无阈值秩超几何重叠检测了表达不同的 DEG(即在 PTSD 中上升而在 MDD 中下降,反之亦然),发现P值和优势比显示一致基因的富集显著较高(87.8%),不一致基因的富集较低。12 个基因在不同细胞类型中存在显著不一致(在 PTSD 中下降而在 MDD 中上升)。值得注意的是,CTNNA3(编码 catenin α3)在 PTSD 的九种神经元亚型中的七种中下调,在 MDD 中上调,HSPA1A(编码热休克蛋白 A1A)在 PTSD 中下调,但在 MDD 内皮细胞中上调。
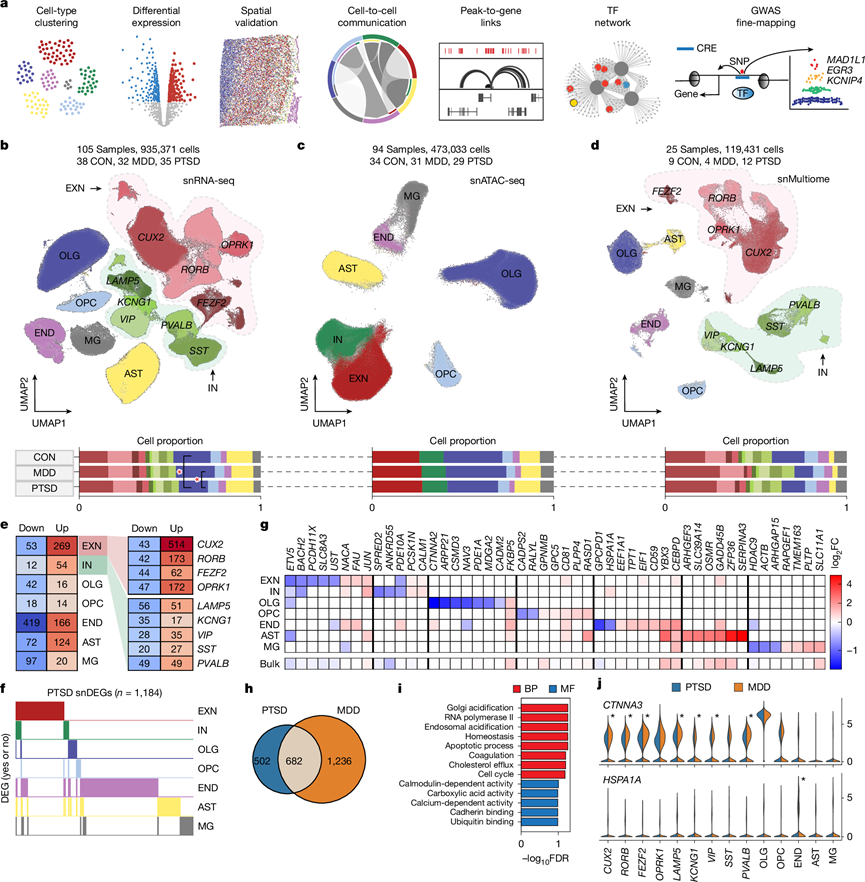
图1. 人类 PFC 中细胞类型的多模态基因组分类和细胞类型特异性基因表达变化。
(a) 分析示意图。(b-d) UMAP可视化。(e) 每种主要细胞类型(左)、EXN 亚型(右上)和 IN 亚型(右下)在两个方向上的显著 DEG 计数,按 DEG 数量着色。(f) PTSD snDEG 在各种细胞类型中的出现情况。(g) 每种细胞类型的DEG 的 log2 FC 值。(h) PTSD snDEG 和MDD snDEG之间的重叠。(i) GO分析。(j) 每种亚型中显著不一致的基因的标准化平均表达。
02
PTSD基因表达变化的空间验证
他们在 DLPFC 的所有细胞类型中都鉴定出了许多转录本表达变化。为了证实这些变化,他们使用 10X Xenium 平台进行空间转录组分析(scXenium),针对基于细胞类型标记和 snRNA-seq 分析中的表达变化而筛选出的一组基因(366 个)(图2a)。他们分析了由体细胞和细胞核分析的空间数据,随后使用 Squidpy 和典型细胞类型注释(图2a)从 snRNA-seq 结果(图2b)中对转录本进行聚类。他们观察到总共 550520 个细胞(scXenium),每个细胞的中位数为 347 个总转录本和 100 个独特转录本。同样,他们观察到总共 523314 个细胞核(snXenium),每个细胞核的中位数为 87 个总转录本和 42 个独特转录本。他们发现软脑膜表面附近的 END 标记(假定的血管)和定位于白质的 OLG 标记显著富集。对细胞类型注释的细胞核和单个转录本的可视化揭示了 DLPFC 的高分辨率详细结构。在 Xenium 后苏木精和伊红染色后,他们没有发现样本之间主要的解剖学差异。
为了验证 snRNA-seq 数据集,他们使用 MAST 探索了病例与对照组之间 DLPFC 所有细胞类型的表达变化(图2c),观察到神经元中大多数基因表达变化。他们将这些结果与snRNA-seq MAST 结果进行了比较,发现几种DEG 重叠最高,包括神经元(73% EXN 和 42% IN)、OPC(64%)和 END(62%)(图2d)。总体而言,他们发现 snRNA-seq 与 snXenium和 scXenium的细胞核转录本均具有高度相关性。 snRNA-seq 和 scXenium 数据之间的高度相关性表明体细胞转录水平准确反映在细胞核中。
他们观察到 END 中 88 个 DEG 发生了显著变化(图2e),其中 31 个与 snRNA-seq DEG 重叠。例如,他们发现FKBP5在 PTSD 非神经元细胞中高表达,并且在 END 中的表达显著增加(图2f)。他们观察到整个 DLPFC 中FKBP5表达量很高(图2g-h),并且在血管附近的水平明显更高(图2g-h)。他们发现上调最多的转录本之一是CXCL14,它是一种在整个大脑的中间神经元中表达的趋化因子,可以下调神经元之间的 GABA 能传递。
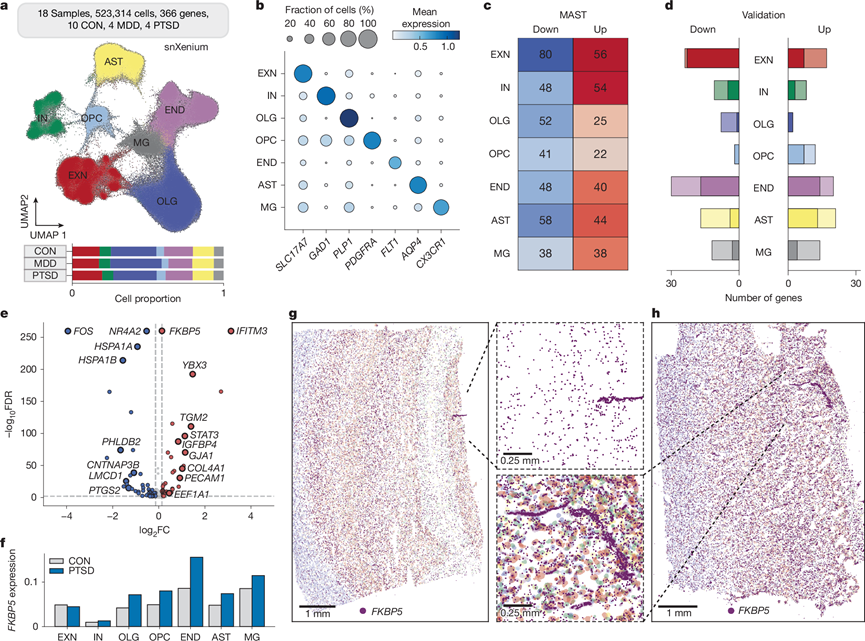
图2. PTSD PFC 的空间转录组分析。
(a) UMAP可视化。(b) 用于细胞类型注释的典型标记的表达。(c) 使用 MAST 在两个方向上对 PTSD snXenium DEG 进行计数。(d) PTSD snRNA-seq 和 snXenium MAST DEG 之间的重叠。(e) 火山图显示 snXenium 在 END 中与 PTSD 和 CON MAST DEG 的比较。(f) FKBP5表达分析。(g) CON 载玻片上的FKBP5转录本以深紫色表示,每个细胞核以其相应的细胞类型颜色表示(END 为浅紫色)。(h) PTSD 载玻片上的FKBP5转录本以深紫色表示,每个细胞核以其相应的细胞类型颜色表示(END 为浅紫色)。
03
PTSD改变中间神经元和小胶质细胞的通讯
为了理解PTSD转录组中表达变化的功能,他们利用经验获得的配体-受体对,基于转录本表达水平构建配体-受体通讯网络。PTSD患者小胶质细胞发送模式减弱,而MDD患者小胶质细胞发送模式增强,这是由小胶质细胞中SPP1转录本的上调以及SPP1(骨桥蛋白)通路中整合素受体的神经元表达所驱动的。与MDD患者(下)相比,骨桥蛋白受体亚基整合素α4在PTSD患者(上)的几种神经元亚型中存在差异调节。
研究结果还表明 PTSD 中的神经元通讯存在差异,这可能是由突触传递的改变引起的。因为神经递质水平无法通过基因表达来测量,他们使用 NeuronChat 通过合成和运输转录物的表达水平来近似神经递质水平。他们分析了 PTSD 和 CON 样本之间神经递质发送细胞信号传导表达的差异。在 PTSD 中,与其他神经元细胞类型相比, SST IN的发送者通讯信号显著减少(图3a),这证实了之前的报告。他们还发现从SST IN到所有其他神经元细胞类型的通讯强度差异在 PTSD 中下调,AST、END 和 OPC 进一步降低(图3b)。此外,他们观察到整个 DLPFC 的 IN中SST转录本显著下调(图3c-e)。值得注意的是,下调最多的通讯强度发生在SST IN与KCNG1 IN之间。与其他抑制性细胞类型相比,SST IN对GABA的利用不足,尤其是GABA转运转录本SLC32A1,导致GABA向GABRA5、GABBR1、GABRB1和GABRG1受体的输出减少(图3f)。
为了验证SST IN通过 GABRA5 向 EXNs传递信息减少的发现,他们使用了啮齿动物创伤应激模型、单次长时间应激 (SPS),并使用膜片钳切片电生理学评估了小鼠 PFC (mPFC) 中的突触功能。他们在SST IN中选择性表达通道视紫红质 2 (ChR2),并记录了对照组和 SPS 小鼠 EXN中光诱发的SST IN 介导的抑制性突触后电流 (eIPSC)(图3g)。在孵育了 GABA 受体 α5 亚基 (GABRAα5) 特异性拮抗剂 MRK 016 后,他们观察到 eIPSC 幅度显著降低(图3h-i),证实含有 GABRAα5 的 GABA 受体是SST IN 到 EXN eIPSC 的主要贡献者。相比之下,SPS 小鼠在 MRK 016 之后平均振幅略有下降,但变化明显小于对照组,表明 GABRA5 介导的SST IN 到 EXN 突触传递显著减少(图3i)。此外,他们还评估了对照组和 SPS 组的紧张性 GABA 电流水平(图3j)。紧张性抑制由 GABA B受体(含有 GABAB R1 亚基;由GABABR1编码)直接介导,以及由GABAB受体调节的突触外 GABAA受体介导。SST IN 已证实可通过紧张性激活 GABAB受体来沉默兴奋性突触传递。他们发现用 CGP 55845 阻断 GABAB受体可导致对照组 EXN 中紧张性电流水平降低幅度明显大于 SPS 小鼠(图3k-l)。总之,这些结果支持以下结论:SPS 小鼠中SST IN 突触传递至 EXN 减少。
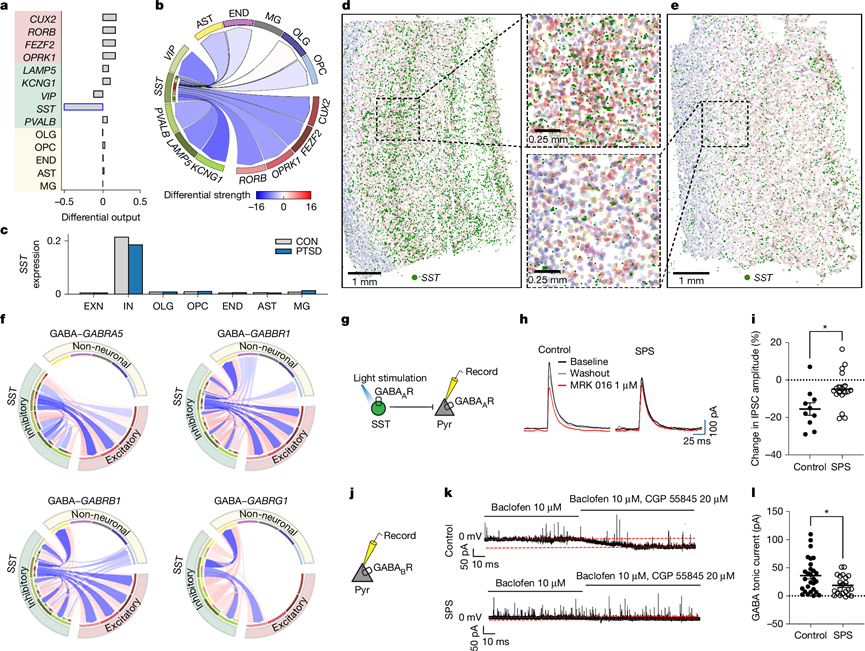
图3. PTSD 中的配体-受体通讯改变。
(a) 所有发送细胞类型的对数差异输出的条形图。(b) Circos 图显示 PTSD 和 CON 之间来自SST IN的所有细胞间相互作用的差异强度。(c) 比较 10 个 CON 和 4 个 PTSD 样本按细胞类型的平均SST对数归一化计数。(d) CON 分析。(e) PTSD分析。(f) Circos 图显示 PTSD 患者的SST IN所有细胞间相互作用的差异输出强度下降。(g) 切片电生理实验装置示意图。(h) 对照组和 SPS 小鼠记录的代表性 eIPSC。(i) 与基线相比,GABARα5 拮抗剂 MRX 016 中 IPSC 幅度变化的百分比定量。(j) 实验装置示意图。 (k) 对照组(上)和 SPS 组(下)的代表性曲线。(l)定量对照组和 SPS 组中 GABA 紧张性电流的大小。
04
PTSD改变基因表达的顺式调控
他们使用 snATAC-seq 分析了染色质的可及性,以探究PTSD 中细胞类型特异性的顺式基因调控机制,在所有细胞类型中发现了 913717 个染色质峰,主要位于基因组的非编码区域内,例如内含子(51.1%)和基因间区域(26.2%)(图4a)。将近一半的峰是细胞类型特异性的(图4b),其中 EXN 和 IN 之间(57.1%)以及 OLG 和 OPC 之间的重叠最多(57.2%)(图4b)。与来自 DLPFC 的主体组织脑开放染色质区域(bCRE) 的比较证实了他们的发现,大多数(88.6%)峰在所有七种细胞类型中平均重叠。将 snATAC-seq 与 6 个高质量 snRNA-seq 样本子集整合,使用并集峰发现了 1433145 个峰与基因之间的联系(图4c)。之前的研究表明,并非所有峰都控制基因表达,因此他们将 CRE 定义为具有峰与基因连锁的峰,并发现 89.18% 的峰是跨细胞类型的 CRE(图4d)。
为了鉴定疾病基因调控机制,他们将 CRE 连锁基因 (CLG) 与每种细胞类型的差异表达基因 (DEG) (CLG-DEG) 进行了交叉分析,发现 END (544) 和 EXN (313)中的 CLG 重叠度最高(图4e)。在 EXN 和 AST 中,大多数 CLG-DEG 与上调基因相对应(84% 的 EXN 和 66% 的 AST),并且不同细胞类型重叠的 CLG 靶标 DEG 大多与不同的 CRE 相关。例如,YBX3在两种不同的细胞类型(END 和 AST)中上调,每种细胞类型都具有独特的、细胞类型特异性的 CRE(图1g)。类似地,FKBP5在多种细胞类型中高表达(图4f),并在非神经元细胞中显著上调(图1g)。他们发现与FKBP5转录起始位点 (TSS) 高度相关的峰到基因链接均为 END 和 MG CRE,可能影响这些细胞类型中的基因表达(图4g)。
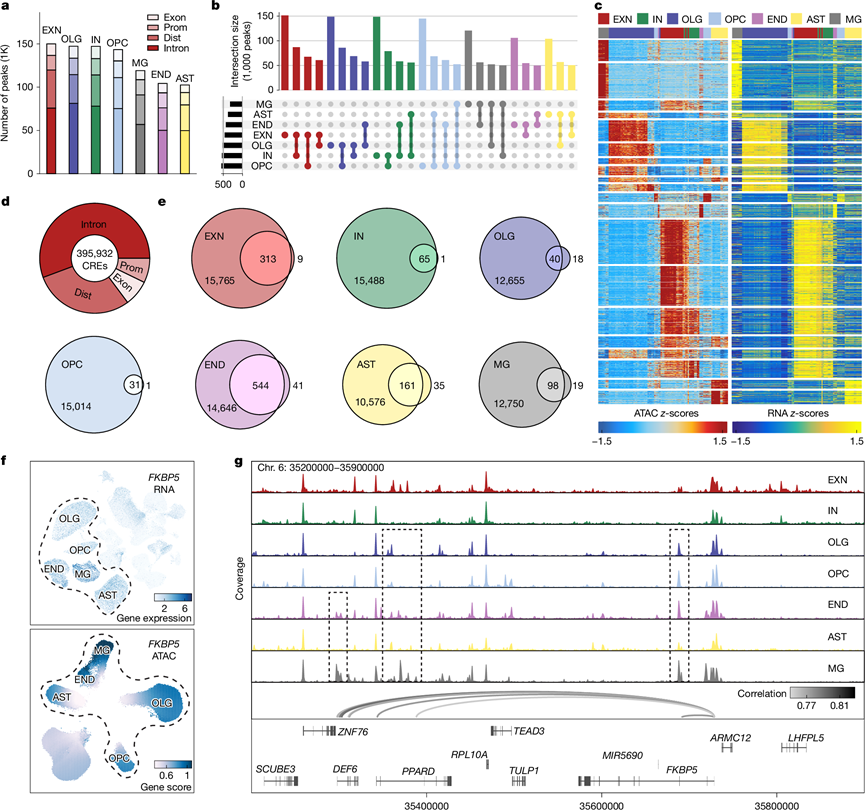
图4. PTSD 改变不同细胞类型基因表达的顺式调控。
(a) snATAC-seq 中主要细胞类型按基因组类别分隔的染色质峰数。(b) 翻转图显示峰数和不同细胞类型之间峰的重叠。(c) 并排的 ATAC(左)和基因(右)区域链接的热图。 (d) 环形图显示按基因组类别分隔的具有显著链接基因的 CRE 数量。(e) 显示每种细胞类型的 CLG 和 PTSD DEG 交集的维恩图。(f) FKBP5的 UMAP 图,以snRNA-seq 对数归一化基因表达(上)和 snATAC-seq 基因评分(下)突出显示。(g) 染色质可及性信号轨迹突出显示6 号染色体(Chr. 6):35200000–35900000 区域内不同细胞类型的FKBP5峰到基因链接。
05
PTSD风险基因位点的细胞类型特异性顺式调控
为了评估遗传风险变异在大脑特定细胞中的作用,他们对 PTSD、其临床症状(过度唤醒、回避和重复体验)、MDD、其他精神疾病以及其他临床和非临床性状 GWAS进行了连锁不平衡评分回归 (LDSC) (图5a),发现 PTSD(总 PCL)和 PTSD 病例对照组的 EXN 和 IN 的 ATAC 峰中 PTSD GWAS 信号显著富集。这些发现也反映在症状 GWAS 中。他们还发现在这些相同的细胞类型中 MDD 风险显著富集。他们将这些发现与 bCRE(灰点)进行了比较,发现所有性状的 LDSC 富集率更高。为了证实这一点,他们使用snMultiome进行了相同的分析,以分析神经元亚型中PTSD风险基因的富集情况。将snMultiome与相同的bCRE(灰色)进行比较,发现LDSC的富集率更高(图5b)。他们在总体PCL GWAS中鉴定出一个EXN亚型(OPRK1),在PTSD病例对照中鉴定出一个IN亚型(KCNG1),这表明两个群体之间可能存在差异。
接下来,他们在 EXN 和 IN 中对细胞类型特异性 CRE 内的 PTSD 相关多态性进行了精细映射,分析了来自之前两项 全基因组关联分析研究(GWAS) 的所有风险基因座,并确定了八个属于细胞类型特异性 CRE 的基因座,重点关注这些基因座进行精细映射(图5c-f)。他们评估了 PTSD 风险单核苷酸多态性(SNP)的后验包含概率 (PIP),发现对于 8 个基因中的 7 个,位于 CRE 内的 SNP,PIP 值与负对数转换的 GWAS P值(平均相关系数 = 0.769)具有高度相关性,这表明精细映射捕获了具有强 GWAS 关联的显著领先 SNP。对于MAD1L1基因,他们发现导致总 PCL 的主要致病变异是百万退伍军人计划 (MVP) 在 IN 中发现的 rs10235664(图5c)。然而,在纳入 IN CRE 后,他们检测到了其他 PIP 得分更高的变异。值得注意的是,变异 rs62444919与 MVP 显著变异 rs10235664表现出高度连锁不平衡。峰到基因环路具有较高的相关值,分别为0.87和0.67,表明ELFN1转录具有较强的增强子活性。他们还使用NeuN + Hi-C数据集轨迹在该区域发现了一个拓扑相关结构域(TAD),证实了ELFN1和MAD1L1之间存在染色体环路。此外,他们还证实了先前在EXN中发现的与KCNIP4基因致病的SNP rs4697248(图5d)。他们重点介绍了六个PTSD基因(EGR3、LRFN5、OPCML、CAMKV、TCF4和CRHR1)在EXN和IN中的精细映射风险SNP,以及它们对应的CRE(图5e-f)。通过将细胞类型特异性的ATAC数据纳入PIP评分,并识别这些基因位点的TAD,鉴定出新的推定因果风险变异,并重点突出了需要进一步进行功能基因组验证的候选变异。
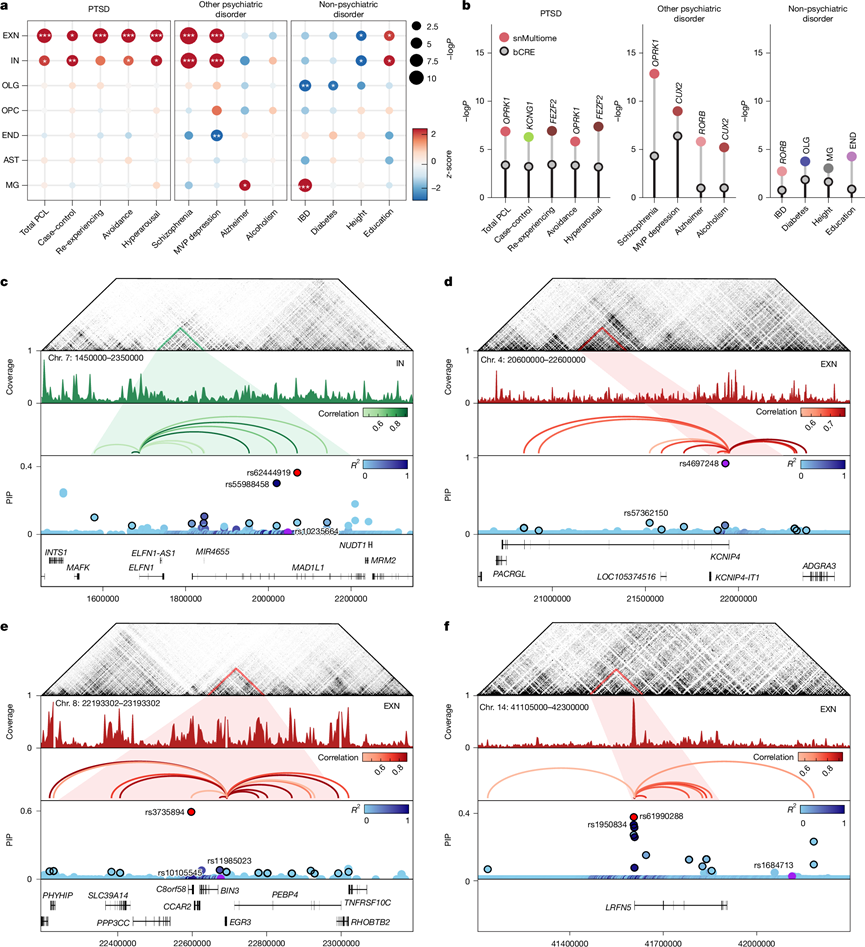
图5. PTSD 疾病遗传风险基因位点的细胞类型特异性顺式调控。
(a) snATAC-seq 细胞类型中 LDSC 对各种 GWAS 特征的富集。(b) 棒棒糖图显示 LDSC 对 GWAS 特征的富集。(c-f) GWAS基因座和细胞类型的顺式调控架构。
+ + + + + + + + + + +
结 论
本研究分析了 111 例人类大脑背外侧前额叶皮层超过 200 万个细胞核的分子反应,这些大脑组织取自患有和不患有 PTSD 及重度抑郁症的个体,并在死后进行组织学分析。本研究鉴定了神经元和非神经元细胞类型的细胞聚类、基因表达变化和转录调控因子,并以细胞类型特异性的方式绘制了 PTSD 的表观基因组调控组图,揭示了抑制性神经元、内皮细胞和小胶质细胞中 PTSD 相关基因的改变,并发现了与糖皮质激素信号传导、GABA 能传递和神经炎症相关的基因和通路。本研究利用细胞类型特异性空间转录组学进一步验证了这些发现,证实了SST和FKBP5等关键基因的破坏。通过整合遗传、转录组和表观遗传数据,本研究揭示了在细胞类型特异性背景下破坏PTSD基因(包括ELFN1、MAD1L1和KCNIP4)的可靠变异的调控机制。总之,这些发现全面表征了创伤应激反应对人类前额叶皮质持续影响的细胞特异性分子调控机制。
+ + + + +



