English
English文献解读|Cell Rep(9.995):胃混合腺神经内分泌癌的精确显微解剖分析其基因组景观和进化克隆起源
+ 论文ID 原名:Precise microdissection of gastric mixed adeno-neuroendocrine carcinoma dissects its genomic landscape and evolutionary clonal origins 译名:胃混合腺神经内分泌癌的精确显微解剖分析其基因组景观和进化克隆起源 期刊:Cell Reports 影响因子:9.995 发表时间:2023.6.6 DOI号:10.1016/j.celrep.2023.112576
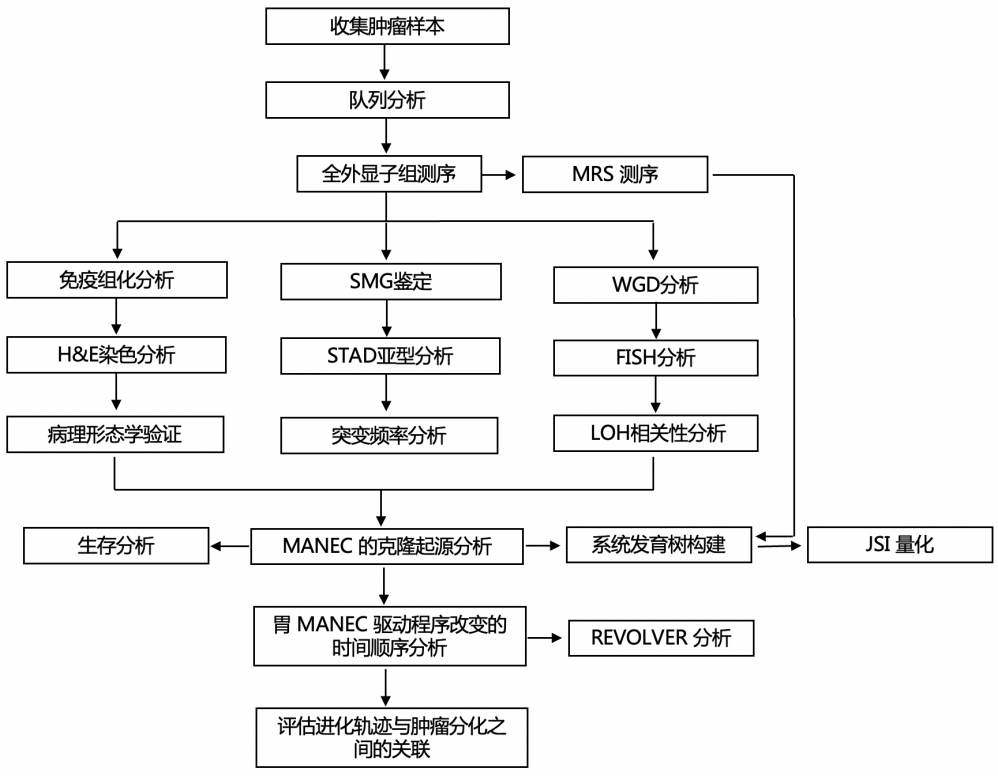
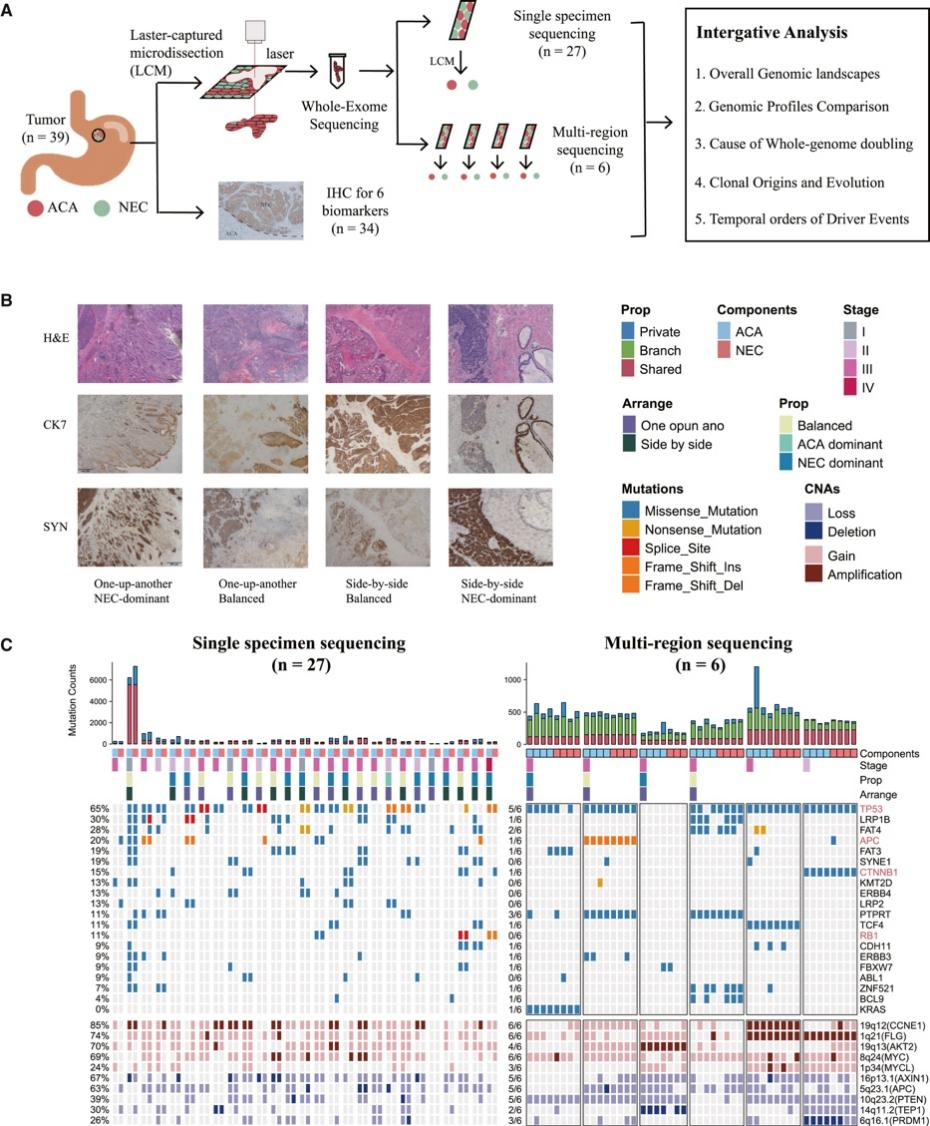
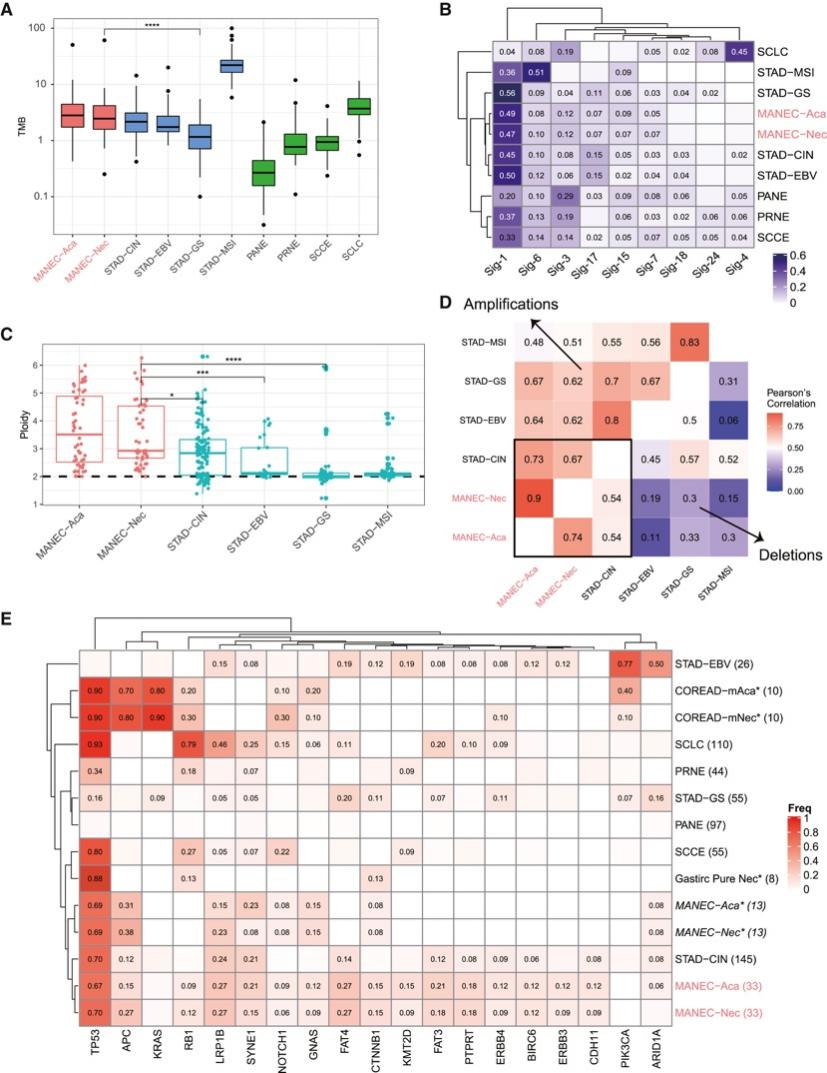
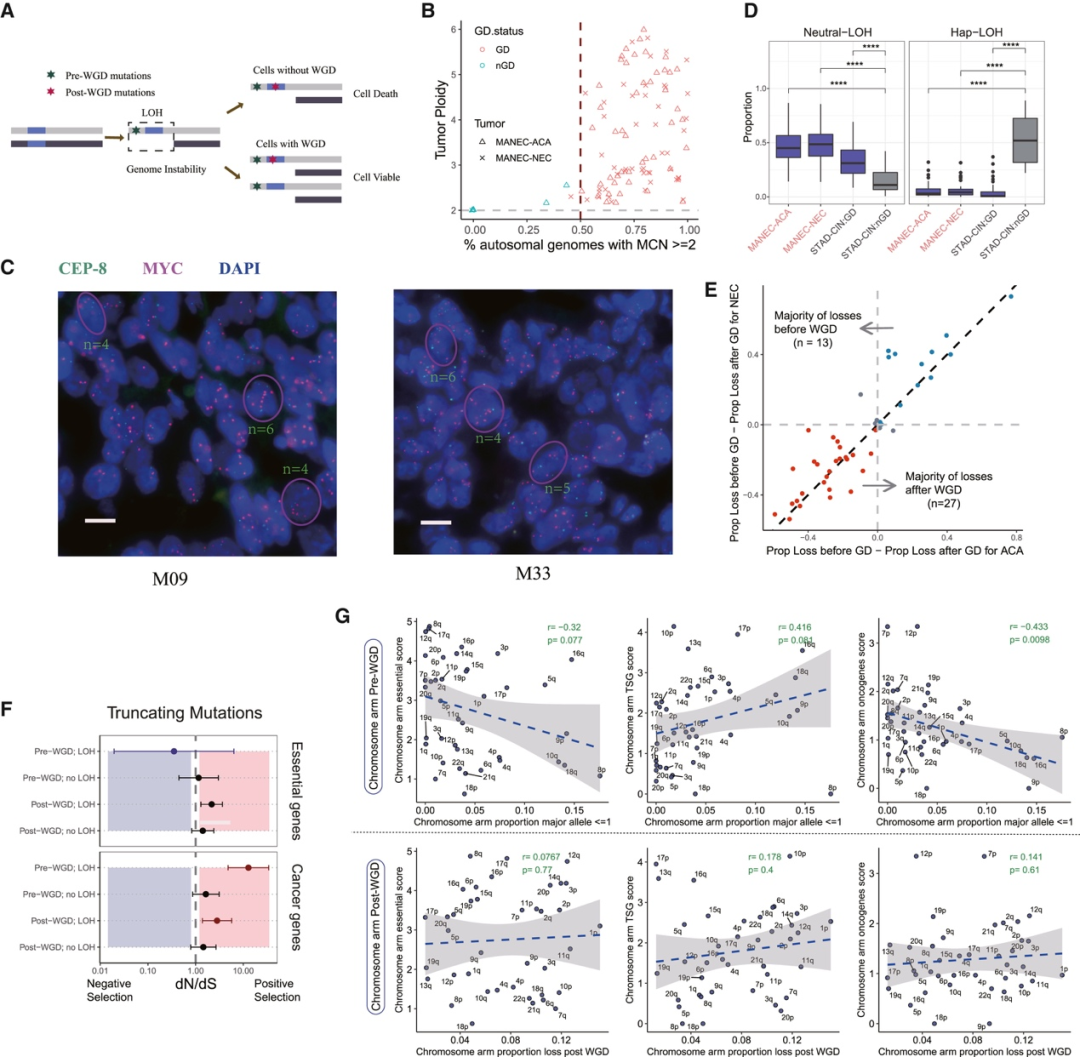
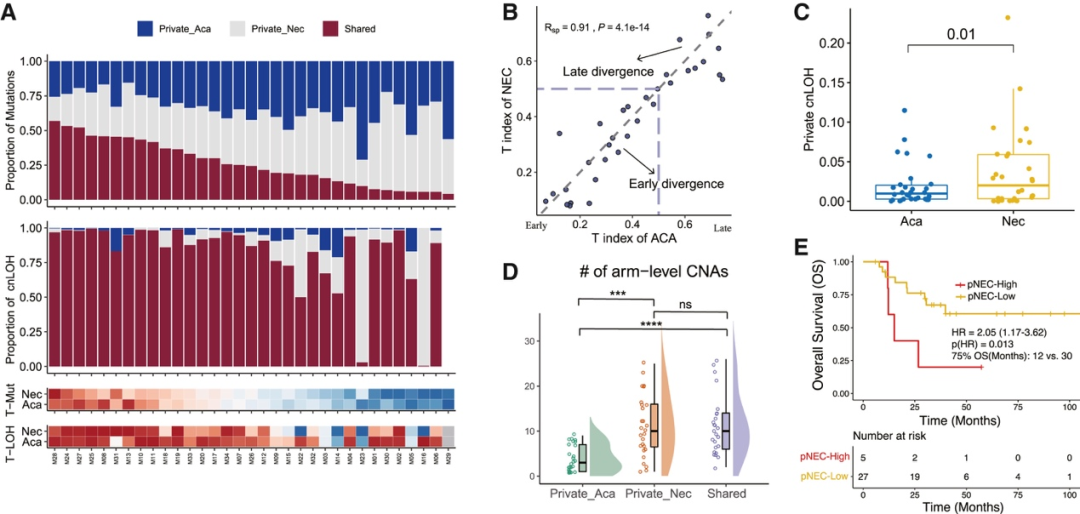
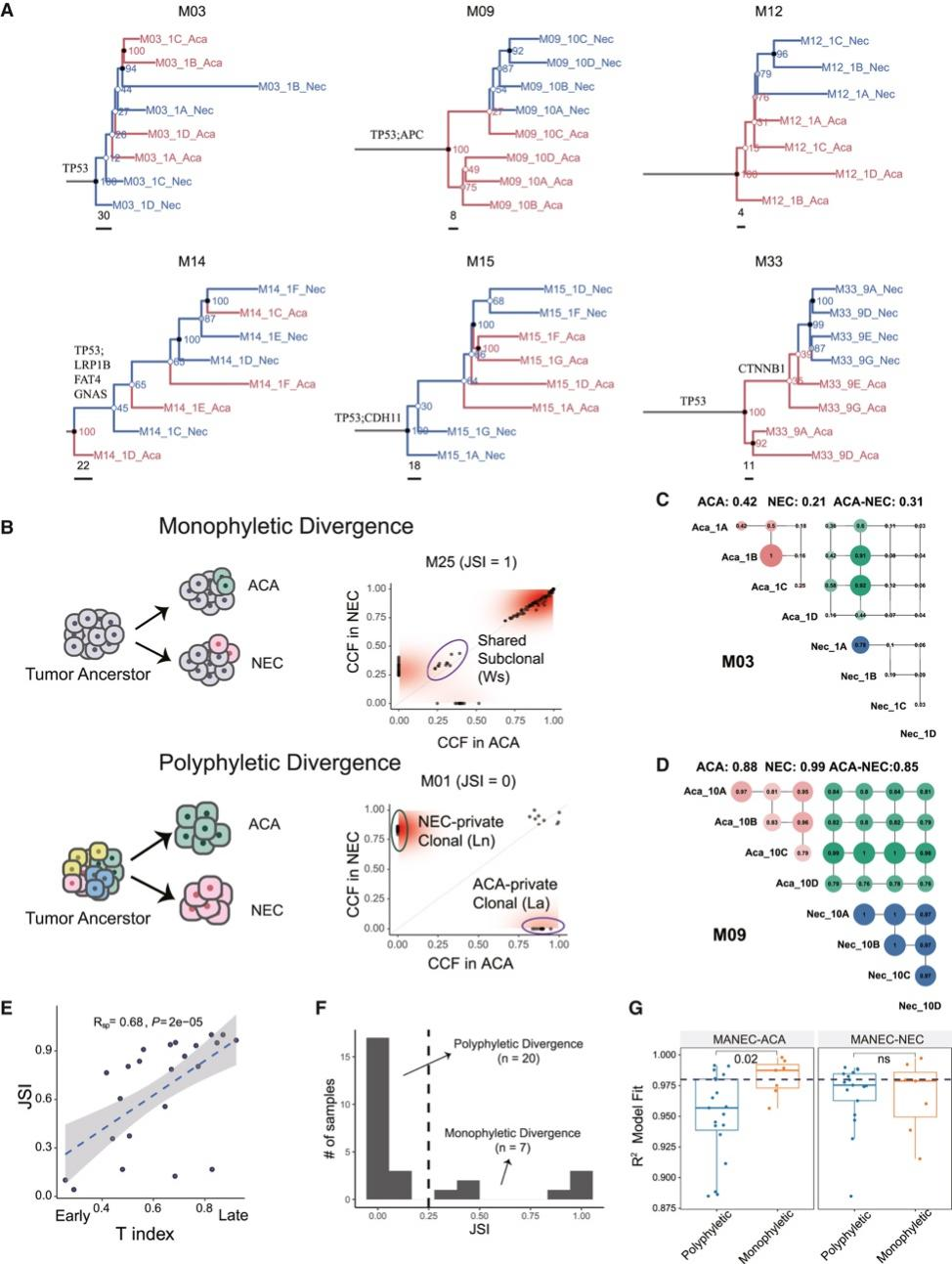
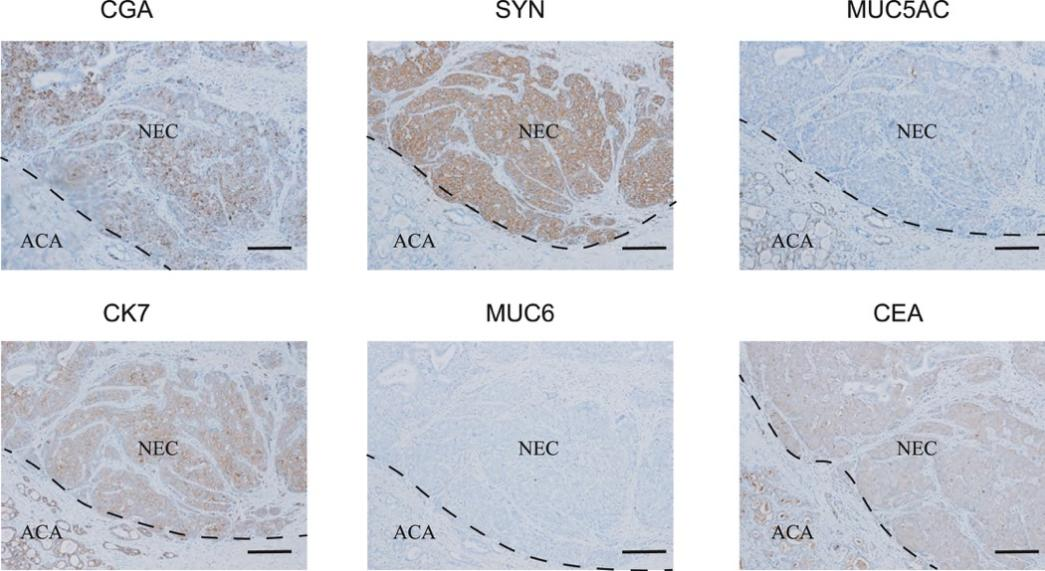
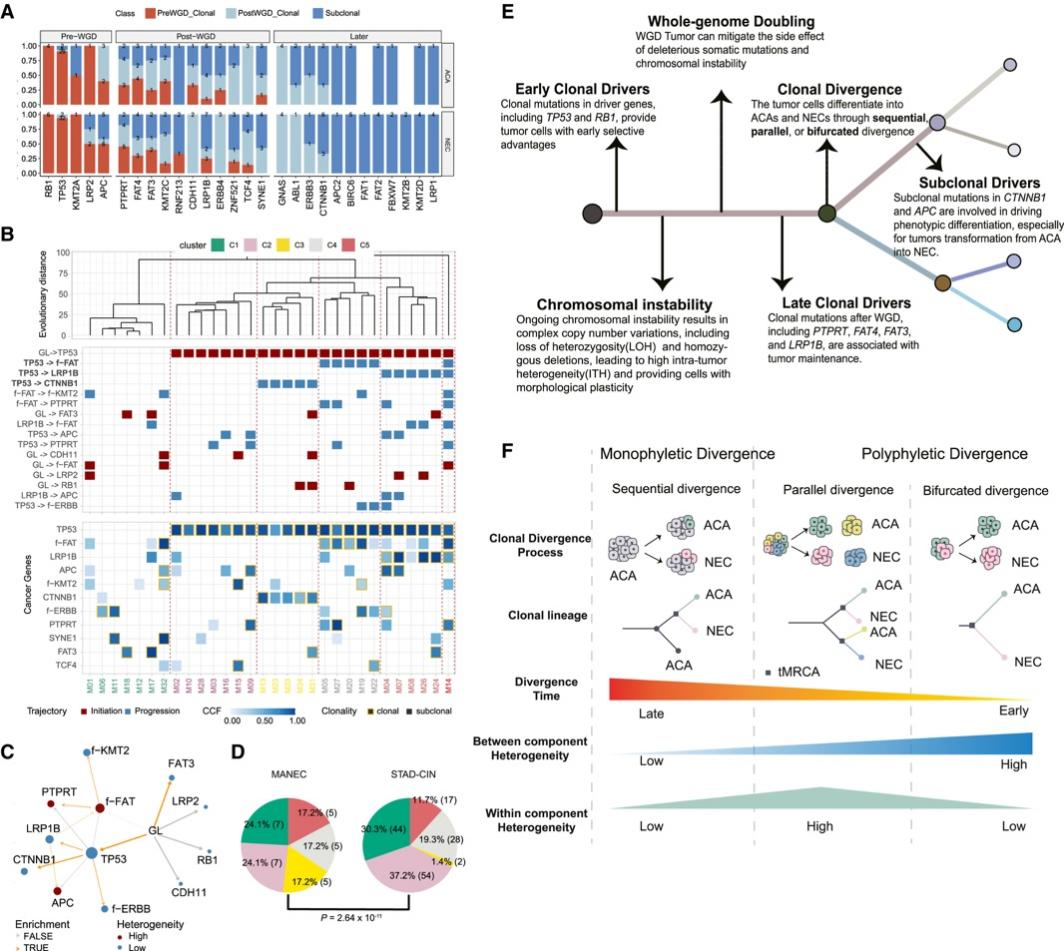
背 景 胃混合性腺神经内分泌癌 (MANEC) 是一种临床侵袭性和异质性肿瘤,由腺癌 (ACA) 和神经内分泌癌 (NEC) 组成,但目前MANEC 的基因组特性和进化克隆起源仍不清楚。 实验设计 结 果 01 胃 MANEC 的患者队列和临床病理特征以及基因组景观 在本研究中,研究者团队回顾性收集了39例胃MANEC病例样本,其中33例用于测序,6例用于验证研究(图1)。来自同一个体患者的两种肿瘤成分的细胞通过激光捕获显微切割 (LCM) 技术分别收集并进行全外显子组测序(图1A)。为了捕捉 MANEC 两个组成部分的进化过程,他们对 6 名患者的同一个体癌症的腺癌 (ACA) 和神经内分泌癌 (NEC) 组成部分进行了多区域测序 (MRS)。 根据两种成分的空间排列和肿瘤细胞组成对病例进行分类(图1B),如果肿瘤成分是分离的,则将病例定义并排;如果肿瘤成分混合,则将病例定义为连续。这些病例还根据肿瘤成分分为 ACA 显性、NEC 显性或平衡。再根据苏木精和伊红(H&E;肿瘤细胞形态学)、细胞角蛋白 7(CK7;ACA 标志物)和突触素(SYN;NEC 标志物)染色分类的 26 例样本中,有 14 例和 12 例相互连续和并排模式,分别有 12、1 和 13 例具有平衡、ACA 主导和 NEC 主导模式。[1] 总共在 ACA(中位数 385,范围 49-6219)和 NEC(中位数 381,范围 36-7279)样本中分别鉴定出 26004 和 26177 个突变,包括各自 11268 和 11231 个非同义突变,4459 和 4430 个同义突变等。此外,ACA 和 NEC 共有 62.1%的体细胞突变,ACA 和 NEC 中分别有 18.7%和 19.2% 的突变是各自的突变。ACA 和 NEC 中反复突变的癌基因的大多数体细胞突变也在患者中共享(图1C)。 他们鉴定出了四个显著突变基因 (SMG),即TP53、RB1、APC和CTNNB1,还鉴定了 MANEC 的显著突变的局灶性拷贝数改变 (CNA)(图1C)。 图1. 胃混合腺神经内分泌癌基因图谱 02 MANEC 在遗传上类似于 STAD-CIN 亚型 为了阐明胃 MANEC 的遗传特性,他们比较了胃 MANEC 与 TCGA STAD 以及来自其他器官的纯 NEC 和 MANEC 的突变特征(图2)。胃腺癌 (STAD) 进一步分为四种亚型,包括基因组稳定(GS)、染色体不稳定性 (CIN)、微卫星不稳定性 (MSI) 和Epstein-Barr病毒 (EBV) 亚型。 胃 MANEC 的肿瘤突变负荷 (TMB) 与 STAD-CIN 和 STAD-EBV 相当,显著低于 STAD-MSI,但高于 STAD-GS(图2A)。与 STAD-GS、STAD-MSI 和来自其他器官的纯 NEC 相比,胃 MANEC 的两个成分的突变特征与 STAD-CIN 和 STAD-EBV 的突变特征更相似(图2B)。MANEC ACA/NEC 的肿瘤倍性高于 STAD-CIN,并且显著高于其他 STAD 亚型(图2C)。CNA 的成对 GISTIC G 分数显示 MANEC 的两个组成部分中的扩增与 STAD-CIN 中的相似(图2D)。 接下来,他们将重点放在STAD和纯NEC中反复突变的癌症基因(图2E),胃 MANEC 中TP53突变的频率(胃 ACA/NEC 为 67% 和 70%)与 STAD-CIN (70%) 相当,高于 STAD-GS (16%) 和 STAD-EBV (<5%)。总的来说,胃 MANEC 的分子特性在很大程度上类似于 STAD-CIN 亚型。 图2. STAD 和来自其他器官的纯 NEC 的基因组图谱比较分析 03 导致 CIN 的 WGD 引起胃 MANEC 胃 MANEC 的倍性增加突出了 CIN 在 MANEC 癌变中的重要作用。肿瘤的多倍体可能是由非整倍体的独立积累引起的,或者更可能是由于具有随机拷贝数变化 (CAN) 的 WGD 事件(图 3A)。对于大多数 WGD 样本,超过 50% 的常染色体的主要拷贝数 (MCN) ≥ 2,这与晚期癌症肿瘤中的 WGD 一致(图3B)。使用MYC(8 号染色体)、CCNE1(19 号染色体)和PTEN(10 号染色体)探针,通过荧光原位杂交 (FISH) 在 3 名患者的 4 个样本中进一步验证肿瘤倍性状态(图3C)。MYC和 8 号染色体的着丝粒在两名患者中普遍扩增,支持 MANEC 中拷贝数的增加(图3C)。 值得注意的是,与 STAD-CIN WGD 肿瘤相比,胃 MANEC ACA/NEC 中有更高比例的基因组受到基因的杂合性缺失(LOH)(图3D)。相比之下,平均而言,胃 ACA/NEC 中只有 2% 和 1% 的基因组是单倍体 LOH,而 STAD-CIN 非 WGD 肿瘤中为 52%(图 3D)。对于大多数样本,WGD 发生的时间早于大多数拷贝数丢失(图3E)。单倍体基因组区域的体细胞突变在加倍之前对肿瘤存活具有有害影响,而 WGD 可以通过提供额外的基因组拷贝来减轻这些影响。相比之下,对癌症基因的分析表明,位于 LOH 区域的突变在单倍体基因组环境中比在重复基因组中受到更强的正向选择(图3F)。 必要基因密度与单倍体每条染色体臂LOH发生率呈弱负相关,表明负选择可能会减少必要基因富集程度高的基因组区域的拷贝数损失。相反,单倍体LOH的发生也受到癌基因的影响,致癌基因富集程度高的染色体拷贝数损失减少。同时,肿瘤抑制基因 (TSG) 丰度较高的基因组片段拷贝数丢失发生率较高。然而,在单倍体区域的重复上没有观察到这些相关性(图3G)。 图3. 胃 MANEC 进化过程中的全基因组加倍 (WGD) 事件 04 MANEC的克隆起源 ACA 和 NEC 的克隆起源对于理解 MANEC 的发病机制特别重要。他们检测了 ACA 和 NEC 之间共有的基因组改变,共有体细胞突变的中位数比例为 0.24,共有拷贝数中性LOH (cnLOH) 的中位数比例为 0.36。这些共有的体细胞突变和 cnLOH 支持所有 MANEC 肿瘤都是单克隆起源的假设(图4A)。大多数 MANEC 肿瘤经历了早期分化(图4B),ACA 和 NEC 的单独突变数量相当,而 NEC 中单独 cnLOH 的比例显著高于 ACA (图4C)。然后他们在染色体臂水平上探索了 ACA 和 NEC 的 CAN,通过比较共有和单独染色体臂的分布,发现 NEC 获得的臂级 CNA 明显多于 ACA(图4D),并且 NEC-单独 CNA 表现出与共有 CNA 相似的基因组分布。高水平的NEC-单独 cnLOH患者生存预后不良的风险增加,而ACA-单独cnLOH水平对预后无显著影响(图4E)。因此,NEC成分在肿瘤分化过程中积累更多的CNA,促进表型分化,决定了MANEC的恶性程度。 图4. MANEC 的克隆起源 05 MRS定义的MANEC的演化 为了系统地研究肿瘤成分之间的进化关系,他们构建了基于体细胞突变和等位基因特异性 CNA的系统发育树,以及来自六个 MRS 肿瘤的肿瘤克隆树(图 5 A)。他们比较了六个 MRS 肿瘤的每个 ACA/NEC 对中突变的癌细胞分数 (CCF)。ACA 和 NEC 对之间的突变相似性由 Jaccard 相似性指数 (JSI) 量化(图5B)。较高的JSI对应于 ACA 和 NEC 之间较高的遗传相似性,他们使用JSI来量化 MRS 肿瘤中样本对之间的 ITH(图5C-D)。具有连续散度的肿瘤(包括 M09、M12 和 M33)表现出比具有平行散度的肿瘤(包括 M03、M14 和 M15)更高的JSI水平(图5C-D)。JSI与分化时间T呈正相关,说明分化较晚的肿瘤在ACA和NEC之间具有较高的遗传相似性(图5E)。根据JSI,平行发散肿瘤具有相对较晚的分化并表现出多系发散。他们检测了单样本测序患者的JSI值,发现大多数肿瘤的JSI接近 0,表明这些肿瘤处于平行分化和多系分化(图5F)。 他们推测不同发散模式下的肿瘤可能处于不同的肿瘤演化模式。事实上,多系分化下的肿瘤在 ACA 成分中表现出更多的非中性进化。正如预期的那样,在平行进化模型中,肿瘤在肿瘤起始过程中具有高度的遗传异质性(图5G)。 图5. MANEC克隆分解过程 为了描绘 NEC-ACA 转变的全貌,他们通过免疫组织化学 (IHC) 分析了 NEC 区域中的 6 种生物标志物,包括 2 种 NEC 特异性(CGA 和 SYN)和 4 种 ACA 特异性生物标志物(MUC5AC、CK7、MUC6、和 CEA)在 34 名 MANEC 患者中(包括 6 名新诊断的 MANEC 患者)(图6)。在NEC成分中,34例患者均有强表达的SYN或CGA,证实了NEC成分的诊断。此外,他们还发现 NEC 成分中 ACA 特异性生物标志物的表达,MUC5AC、CK7、MUC6 和 CEA 的阳性率分别为 76.5%、47.1%、23.5% 和 70.6%。他们发现 34 名患者中有 33 名至少表达了 4 种 ACA 特异性标记物中的一种。然而,在 ACA 成分中,没有患者表达 SYN 或 CGA,上述结果强烈支持 ACA 到 NEC 的转变。 图6. 通过免疫组织化学验证肿瘤成分转换 06 胃 MANEC 驱动程序改变的时间顺序 驱动突变的时间顺序可以提供有关癌症进展所涉及的分子过程的信息。他们根据 WGD 事件的时间及其克隆状态将癌症基因突变分为 WGD 前克隆、WGD 后克隆和亚克隆突变(图7A)。五个基因(RB1、TP53、KMT2A、LRP2和APC)中的大多数突变是克隆性的并且发生在 WGD 之前,表明它们在肿瘤起始中的作用。相反,其他驱动程序事件,包括PTPRT、FAT4、FAT3和LRP1B中的突变,虽然主要是克隆,但通常发生在基因组复制之后,表明它们参与了肿瘤维持。其余的癌症基因,包括GNAS、ABL1、ERBB3和CTNNB1,大多是亚克隆事件并且单独发生在 ACA 或 NEC 中,表明它们在表型分化中的作用。 驱动突变的时间顺序可以从进化树中推断出来,他们使用了一种迁移学习方法 (REVOLVER) 来捕获不同患者之间反复发生的基因组改变的重复进化轨迹,同时合并单等位基因和双等位基因事件(图 7 B)。REVOLVER 确定了几个重复的进化转变,这些转变以 5 个聚类为特征,即 TP53 野生型 (C1)、TP53仅突变 (C2)、TP53到CTNNB1轨迹 (C3)、TP53到FAT家族轨迹 (C4) 和TP53 -到- LRP1B轨迹 (C5)(图7 B)。当所有的轨迹组合成一个基因网络时,发现有4个基因发生了独立于TP53的突变,即RB1、LRP2、FAT3和CDH11(图7C)。三个进化轨迹涉及初始TP53突变,然后扩展到下游基因的突变,包括TP53到CTNNB1 (C3),TP53到 F-FAT 到PTPRT(或 F-KMT2)(C4),以及TP53到LRP1B到APC (C5),这些突变聚类的患者分布在胃 MANEC 和 STAD-CIN 之间存在显著差异(图7D)。 他们从肿瘤进化的角度对 WGD 如何对 MANEC 发挥作用进行了详尽的解释(图 7E),同时提出了三种模型来描述肿瘤的发散过程,包括连续发散、平行发散和多系发散(图7F)。 图7. 胃MANEC癌基因突变的时间顺序 + + + + + + + + + + + 结 论 本项研究对来自 33 名患者的 101 个样本进行了全外显子组和多区域测序,以阐明它们的进化路径,确定了四个显著突变的基因,TP53、RB1、APC和CTNNB1。MANEC 类似于染色体不稳定的胃腺癌,因为 MANEC 中的全基因组加倍占主导地位,并且发生时间早于大多数拷贝数丢失。所有肿瘤都是单克隆起源的,NEC 成分显示出比 ACA 成分更具侵袭性的基因组特性。系统发育树显示出两种肿瘤发散模式,包括连续发散和平行发散。此外,ACA和NEC优势区域的6种生物标志物的免疫组化证实了ACA到NEC的转变,而不是NEC到ACA的转变。这些数据提供了对 MANEC 的克隆起源和肿瘤分化新的见解。