

 English
English文献解读|J Hepatol(33):多组学分析揭示肝母细胞瘤的连续分化谱和 Wnt-MDK 驱动的免疫逃避
✦ +
+
论文ID
原名:Multiomic analysis uncovers a continuous spectrum of differentiation and Wnt-MDK-driven immune evasion in hepatoblastoma
译名:多组学分析揭示肝母细胞瘤的连续分化谱和 Wnt-MDK 驱动的免疫逃避
期刊:Journal of Hepatology
影响因子:33
发表时间:2025.02.01
DOI号:10.1016/j.jhep.2025.01.031.
背 景
肝母细胞瘤 (HB) 是儿童肝脏最常见的原发性恶性肿瘤,其发病率目前估计约为每百万 1.5 例,近几十年来一直在上升。平均每个肿瘤有 3.9 个突变,与成人癌症相比,甚至与其他儿童癌症相比,HB 的突变负担较低。HB 中的大多数复发性基因突变会导致 Wnt/β-catenin 信号通路的异常激活,其中CTNNB1改变导致 80% 的病例中 β-catenin 降解减少。对肿瘤微环境复杂机制的理解已为成人癌症带来了有希望的新疗法,但儿童癌症的类似疗法仍然缺乏。本研究旨在对肝母细胞瘤的肿瘤微环境进行全面分析,揭示其空间结构和关键信号传导机制。
实验设计
结 果
01

肝母细胞瘤肿瘤细胞表现出肿瘤间和肿瘤内的异质性
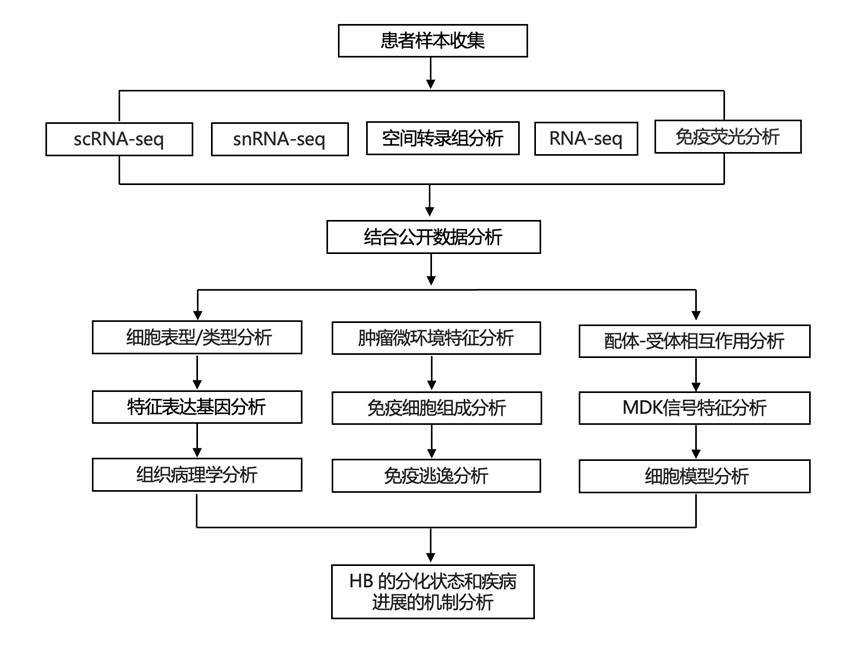
为了阐明HB中的肿瘤异质性和TME景观,研究团队对15份人类HB样本进行了单细胞转录组分析(scRNA-seq)和单细胞核转录组分析(snRNA-seq),并分别对22份和7份样本进行了空间转录组学(ST)分析和多重免疫荧光(mIF)染色。此外,他们还分析了先前发表的包含99份HB和11份非肿瘤及胎儿肝脏样本的转录组分析(RNA-seq)数据,以及包含健康肝脏灌注中免疫细胞的scRNA-seq数据(图1 A)。
本研究的HB scRNA-seq 队列包含来自 5 名患者的 10 个样本,其中包括 3 个未接受化疗的样本、4 个接受化疗的样本和 3 个肺转移样本,共得到 43441 个细胞,对这些细胞进行了进一步分析。整合所有 10 个样本、均匀流形近似和投影降维(UMAP)以及聚类分析,形成了 27 个具有不同基因表达谱的聚类,这种聚类在不同样本之间有很大重叠。结合典型标记基因表达、GO 分析和推断的拷贝数变异,他们鉴定出肿瘤、淋巴细胞、髓样细胞、内皮细胞和肝星状细胞群(图1B)。
总共 16978 个肿瘤细胞聚集成六组(图 1C)。其中四组(此处称为“发育 1”、“发育 2”、“发育 3”和“发育循环”)显示PDE4D、AXIN2、SOX11和PTCH1等基因上调(图 1D)。 GO 分析表明与细胞外基质 (ECM) 和早期发育相关的生物过程活性升高,例如Wnt 信号通路的调节、干细胞增殖和间充质发育;因此,他们将这组肿瘤细胞分配到称为“发育”的程序中(图 1E)。其中,“发育循环”除发育基因外,还显示与有丝分裂相关的基因上调,表明处于活跃增殖状态。相反,另一个聚类的特点是载脂蛋白和细胞色素 P450 基因家族基因高表达,与脂质和肝脏特异性代谢有关(图 1D-E)。这组分化程度更高的肿瘤细胞可归类为“代谢”。“代谢”和“发育”这两个程序在某种程度上对应于之前描述的 HB 分类,例如 公开研究中的 C1/C2 分类,其中 C1 组的特点是发育阶段较晚、肝脏代谢和更有利的结果,而 C2 组显示出不同的表型和不利的结果。来自“代谢”程序的细胞显示出 C1 特异性基因的更高表达,而 C2 特异性基因在“循环”细胞中具有更高的表达。此外,公开数据中定义的“肝细胞”组与本研究的“代谢”组有相似之处,而的“肝脏祖细胞”和“间充质”组都反映在本研究的“发育”程序中。有趣的是,除了这些程序之外,本研究还确定了另一个聚类为“中间”,其中细胞表现出早期发育和 ECM 的转录模式,同时还表现出肝脏代谢特征(图 1D-E)。在本研究的大多数样本中发现,“中间”细胞似乎代表过渡状态。同样,他们分析的大多数样本中也有来自其他程序的细胞,表明肿瘤内异质性程度高。虽然“循环”程序主要由来自一个未接受化疗的原代 HB 样本的细胞组成,但“发育”和“中间”程序的细胞趋于在化疗处理的样本中富集。他们在单独的五个 HB 样本的 snRNA-seq 队列中验证了这些结果,从而验证了所识别程序的稳健性和一致性。
为了证实其存在并进一步探索其相关性,他们为“发育”、“中间”、“代谢”和“循环”肿瘤程序创建了基因特征,以分析分数并比较 公开 RNA-seq数据集内的不同条件和不同的患者结果。他们发现“代谢”程序所代表的转录环境并不特定于肿瘤,也存在非肿瘤肝脏和胎儿肝脏样本(图 1 F)。与未经治疗的原发性 HB 样本相比,该程序在复发或转移等后期条件下表达较低。与此相反,“发育”程序对肿瘤具有高度特异性,在后期条件下表达增加。然而,他们没有看到“发育”评分与患者结果之间的联系。从“循环”程序来看,肿瘤中的富集程度明显高于非肿瘤肝脏,而在胎儿肝脏样本中的富集程度甚至更高。此外,“循环”评分从未治疗的原发性肿瘤到治疗、复发和转移性肿瘤逐渐增加。“中间”程序显示出对肿瘤样本高度特异性的模式,并且在患者预后较差的肿瘤样本中显示评分增加。另一个引人注目的观察结果是复发样本中“循环”和“发育”程序的活性升高,同时“代谢”程序的活性降低。这意味着复发性肝癌的侵袭性体现在更未分化和增殖的状态中(图 1 F)。
为了阐明肿瘤的空间组成以及scRNA-seq鉴定出的肿瘤程序,他们分析了22个人类HB样本的ST数据。结果显示,多个程序可以在一个样本中共存(图1 G)。虽然他们确实在某些样本中观察到“循环”或“中间”程序的明显富集,但与本研究的scRNA-seq数据类似,每个样本的大部分内容都反映在“发育”和“代谢”转录程序中。
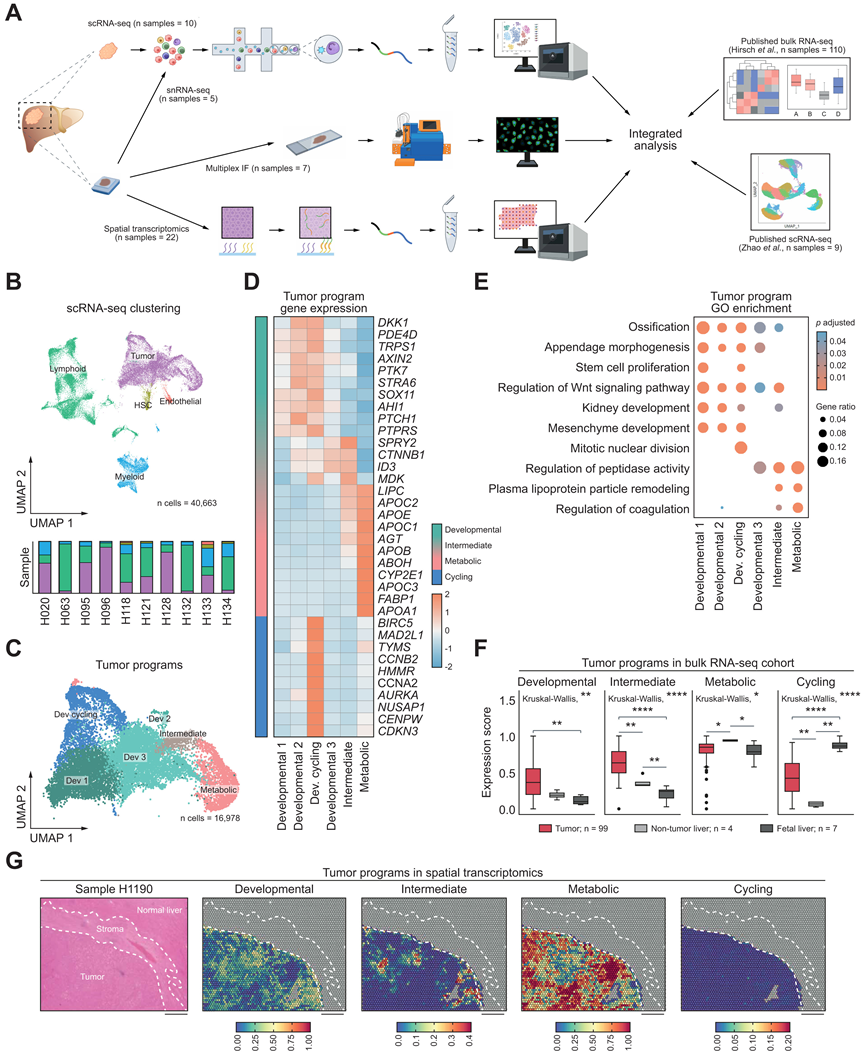
图1. 肝母细胞瘤肿瘤细胞表现出肿瘤内异质性。
(A)实验设计。(B) 来自 10 个肝母细胞瘤肿瘤样本的细胞的 UMAP 表示,按主要细胞类型着色,包括显示每个样本中细胞类型代表性的条形图。(C) 仅肿瘤细胞的 UMAP 表示,按肿瘤程序着色。(D) 显示代表每个肿瘤程序的标记基因的热图。(E)GO分析。(F) 每个肿瘤程序的基因特征表达分数的箱线图。(G) 组织病理学注释的肝母细胞瘤切片的H&E染色以及相应的空间转录组学图,显示了肿瘤点处各肿瘤程序的丰度估计值。
02
肝母细胞瘤微环境的特点是免疫细胞多样性高
鉴于 scRNA-seq 队列中非肿瘤细胞的比例很大,更具体地说,免疫细胞的比例也很大,他们对这个部分进行了更详细的注释,发现淋巴细胞类型高度多样化,尤其是 T 细胞(图 2A)。他们鉴定了 CD8+细胞毒性 T 细胞、CD8+滤泡细胞毒性 T 细胞、CD8+ T 记忆细胞、粘膜相关不变 T 细胞、幼稚 T 细胞以及 CD4+ T 记忆细胞和 CD4+T 调节细胞 (Treg)。此外,他们发现了常规和组织驻留自然杀伤 (NK) 细胞、先天淋巴细胞、B 细胞、浆细胞样树突状细胞和一个独特的循环淋巴细胞聚类。
髓系区室约为淋巴区室大小的三分之一,由 CD14+和 CD16+单核细胞、肿瘤相关巨噬细胞 (TAM) 和两种类型的树突状细胞(常规树突状细胞 1 型和 2 型)以及肥大细胞组成(图 2B)。这些细胞类型的分布在样本中存在相对和绝对差异。考虑到不同细胞类型数量众多,值得注意的是,所有细胞类型在大多数样本、患者和疾病条件下均有体现。
为了更仔细地检查不同免疫细胞类型的丰度,特别是与正常肝脏相关的丰度,他们公开的 RNA -seq数据中肿瘤和非肿瘤样本之间细胞类型特异性标记的表达,与非肿瘤和胎儿肝脏样本相比,肿瘤样本中 B 细胞富集的评分以及细胞毒性淋巴细胞与 Treg 之间的比率显著降低(图 2C)。此外,与原发性 HB 相比,B 细胞的估计值在复发或转移等后期阶段呈负相关趋势,并且从单次手术治愈的 HB 到复发性 HB 和以死亡告终的 HB,B 细胞的估计值显著下降。这表明 HB 具有独特的免疫细胞特征,与较差的预后相关。
他们将本研究的HB scRNA-seq队列与公共scRNA-seq数据整合在一起,该数据包含来自健康肝灌注、脾脏和外周血的免疫细胞(图2 D)。两个数据集均显示出相似的髓系细胞以及T细胞和NK细胞的相对数量。然而,HB样本中的Treg细胞数量明显较高,而B细胞和浆细胞的数量较低(图2 E)。在髓系区室中,与公共数据集中的基因特征相比,HB 巨噬细胞中的几个基因显著上调(图 2 F),包括MSR1(一种 TAM 标志物,已证明可以预测不同肿瘤实体的较差预后)和EGR1(一种可以抑制炎症基因并驱动肝单核细胞衍生的巨噬细胞从促炎状态转变为抗炎状态的转录因子)。这些基因还包括S100A10,它下调 Toll 样受体功能CXCL8(可以抑制不同胃肠道癌症中的 T 细胞浸润)和髓系检查点分子VSIR。

图2. 肝母细胞瘤中的免疫区室表现出改变的细胞类型组成。
(A) 10 个肝母细胞瘤 scRNA-seq 样本中淋巴细胞的 UMAP 表示,按细胞类型着色。(B) 10 个肝母细胞瘤 scRNA-seq 样本中髓系细胞的 UMAP 表示,按细胞类型着色。(C) 肿瘤和非肿瘤样本的 B 细胞系(左)和细胞毒性淋巴细胞与 Treg 评分之间的比率(右)。(D) UMAP 图整合了免疫细胞数据。(E) 条形图显示细胞类型比例。(F) 巨噬细胞的基因表达。
03
空间解析分析揭示肝母细胞瘤的免疫逃逸
接下来,他们整合了本研究的 ST 和 scRNA-seq 数据集,以使用 Seurat 中实现的基于锚点的方法获得每个 ST 点的细胞类型丰度估计值,揭示了丰富的淋巴细胞和不同数量的髓样细胞,它们似乎通常位于肿瘤边界。然而,最引人注目的结果是明显缺乏免疫细胞浸润到肿瘤细胞区域(图 3 A)。通过定量肿瘤细胞、基质和邻近正常肝区域中的免疫细胞丰度证实,与正常肝脏相比,肿瘤中的淋巴细胞和髓样细胞减少(图 3 B)。支持这一观察的现象是,肿瘤接近度以及淋巴和髓样邻域评分呈负相关。比较未治疗和化疗治疗的样本表明,与治疗样本相比,一些治疗样本在邻近正常肝区域中的淋巴丰度略低,而一些未治疗样本在肿瘤区域中的髓样丰度略高。然而,在未处理和处理过的样本中,免疫逃避的总体观察结果是一致的。
为了更好地从功能上理解空间组成,他们通过对每个样本执行 NMF 生成元程序 (MP),将结果压缩为稳健的 NMF 模块,并将其聚合为 7 个 MP。GO 分析显示,MP 代表 ECM 组织、肝脏代谢、免疫过程、染色质重塑和细胞周期等一般过程,以及不同的发育过程、细胞凋亡和激素反应。当将代表免疫过程的 MP 3 与肿瘤邻域评分关联时,他们再次看到负相关性。
之后,他们对 ST 中分析的七个相同 HB 样本进行了 mIF 染色,在蛋白质水平上验证了免疫细胞,尤其是淋巴细胞的排除,这些细胞在肿瘤细胞区域中基本上不存在(图 3C)。非肿瘤区域的淋巴细胞密度及其比例(相对于所有细胞)明显高于肿瘤区域(图 3D-E)。他们观察到髓系浸润程度仅略高。有趣的是,许多确实浸润到肿瘤细胞区域的巨噬细胞的CD204蛋白(由MSR1编码)呈阳性。为了发现这种免疫细胞排斥的潜在机制,他们通过定义肿瘤核心和肿瘤边界区域进一步探索了 ST 数据(图 3 F)。大约一半的斑点指定为非肿瘤,而 45% 和 7% 分别定义为肿瘤核心和肿瘤边界。对这两个区域之间的GO 分析表明,肿瘤核心中与肝脏代谢相关的过程上调,而肿瘤边界区域中与 ECM 沉积和包膜形成相关的过程增高(图 3 G)。由于这是已知的免疫逃避因素,它可能是导致 HB 肿瘤区域缺乏免疫细胞浸润的原因之一。
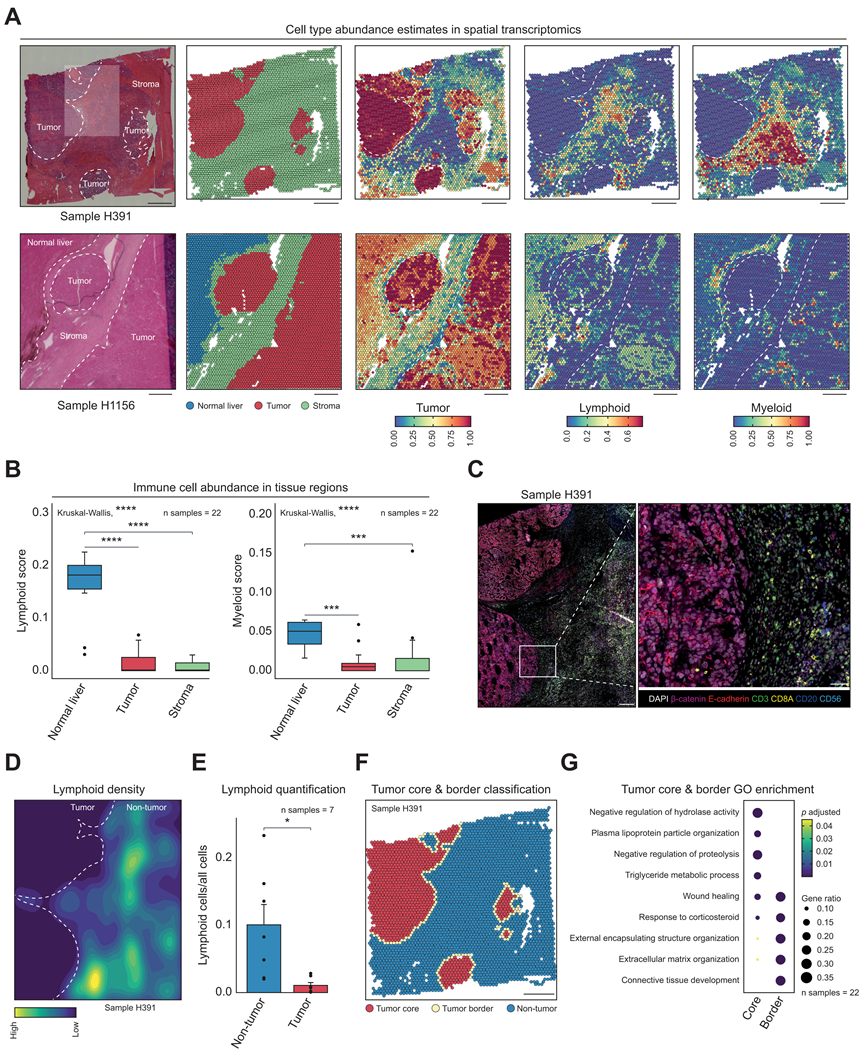
图3. 肿瘤微环境的空间解析分析揭示了肝母细胞瘤的免疫逃逸。
(A) 两份经组织病理学注释的肝母细胞瘤切片的 H&E 染色,以及相应的 ST 图,显示样本内各个点的区域注释和细胞类型丰度估计值。(B) 箱线图比较了 22 份 ST 肝母细胞瘤样本的正常肝脏、肿瘤和基质中淋巴细胞和髓系细胞丰度估计值的中位数。(C) 一份肝母细胞瘤样本的多重免疫荧光。(D) 基于多重免疫荧光细胞分割数据中淋巴细胞蛋白表达评分的淋巴细胞密度图。(E) 七个多重免疫荧光样本中淋巴细胞的平均比例。(F) 样本 H391 的空间图,按肿瘤核心、肿瘤边界或非肿瘤分类着色。(G) GO分析。
04
肿瘤特异性表达促进肝母细胞瘤中中期因子信号升高
为了更好地了解信号环境和 HB 调节其免疫微环境的潜在机制,他们利用 CellChat探索了主要细胞类型之间的配体-受体相互作用,特别是从肿瘤细胞传出的相互作用。最强的传出信号模式之一是中期因子 (MDK) 信号(图 4 A),近年来它作为不同癌症中的关键参与者而受到关注。在 MDK 的受体中,NCL在所有细胞类型中普遍表达,LRP1在髓系细胞和肝星状细胞中特异性表达,ITGA4仅在髓系和淋巴细胞中表达(图 4 B),除了肿瘤细胞本身之外,它在接收和影响 MDK 信号方面发挥着最重要的作用(图 4 C)。
最近描述 MDK免疫调节作用的研究与本研究关于免疫逃避以及促炎和抗炎淋巴细胞之间比例变化的关键观察结果很好地吻合。例如,已发现 MDK-NCL 与抑制免疫活动相关,而 MDK-LRP1 则已证明可诱导抑制性巨噬细胞表型,进而导致 Treg 募集增强。根据这些数据,他们发现MDK的表达与 HB 的 RNA-seq数据中细胞毒性淋巴细胞、NK、B 和单核细胞的估计丰度之间存在显著的负相关性(图 4D)。
虽然之前没有关于 HB 的描述,但他们发现MDK基因在肿瘤细胞中高度特异性表达。在单细胞分辨率下(图 4B)以及在 ST 数据(图 4E-F)中观察到了这种模式。此外,他们在 HB 的免疫荧光染色中发现高水平的 MDK(图 4G)。RNA-seq数据显示,与非肿瘤和胎儿肝脏样本相比,肿瘤中MDK表达升高,此外,与未接受化疗的 HB 相比,接受化疗和转移性的 HB 中的表达显著更高(图 4H)。值得注意的是,与单次治愈性手术等预后较好的患者的样本相比,因癌症死亡的患者的样本表现出显著更高的MDK表达(图 4H),强调了 MDK 与 HB 的相关性。
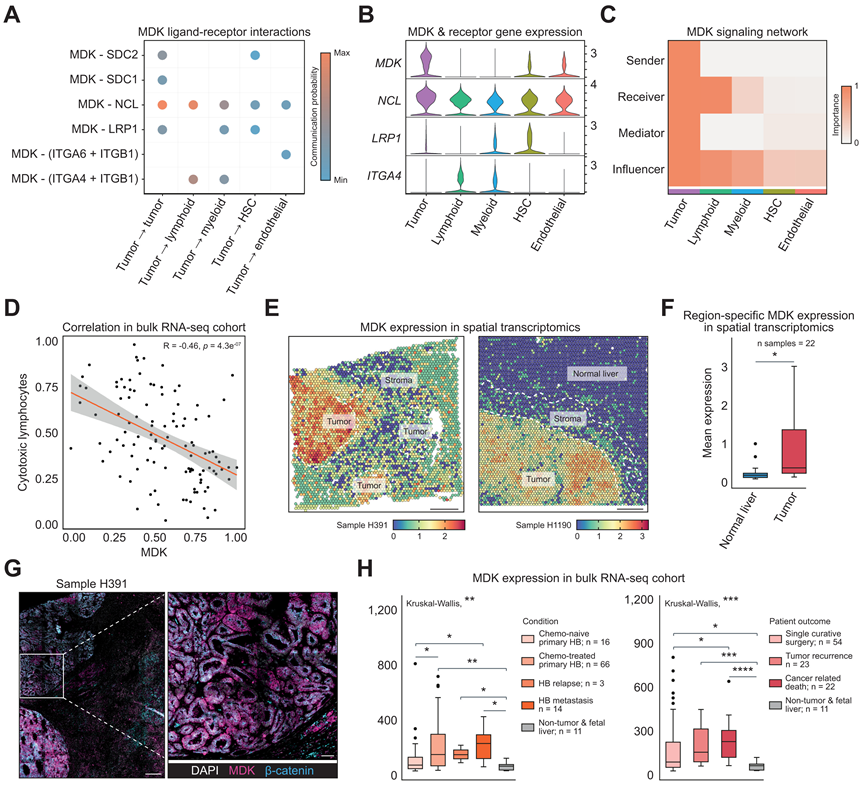
图4. 肿瘤特异性表达促进肝母细胞瘤中中期因子信号传导增强。
(A) 基于 scRNA-seq 数据预测肝母细胞瘤主要细胞类型中涉及 MDK 的配体-受体对显著富集。(B) 小提琴图显示肝母细胞瘤 scRNA-seq 数据中MDK及其受体的表达水平。(C) 热图显示 CellChat 预测的肝母细胞瘤中 MDK 信号的富集情况以及主要细胞类型的相对重要性。(D) MDK表达与细胞毒性淋巴细胞评分之间的相关散点图。(E) 空间图显示两例肝母细胞瘤 ST 样本中肿瘤特异性MDK 的表达情况。(F) 箱线图显示22 例肝母细胞瘤 ST 样本正常肝脏和肿瘤区域的平均MDK表达情况。(G) 肝母细胞瘤样本的多重免疫荧光分析。(H) MDK表达的箱线图。
05
β-catenin表达增加介导中期因子上调
先前的研究表明,Wnt/β-catenin信号转导可导致神经胶质瘤细胞中的MDK上调,这可能是通过β-catenin与MDK启动子区的TCF/LEF结合位点结合实现的。鉴于Wnt/β-catenin信号转导在HB中发挥的关键作用,他们旨在探究这种与HB中MDK的关联。
他们发现RNA-seq 数据中KEGG Wnt 信号通路特征和MDK的表达之间存在正相关(图 5A),而ST 数据中CTNNB1和MDK的表达之间存在正相关。为了进一步检验 Wnt 激活和MDK表达之间的因果关系,他们用 Wnt 信号抑制剂 PKF118-310 (PFK) 或 MDK 抑制剂 iMDK 处理 HepG2 和 Huh6 HB 细胞系,并通过免疫印迹和 RNA-seq 分析验证其效果。与未处理的细胞相比,观察到用 PKF 处理后 MDK 水平适度但显著降低。此外,抑制 MDK 后 β-catenin 下调,暗示 MDK 和 β-catenin 之间存在正反馈回路。
在转录水平上评估 Wnt 和 MDK 抑制对 HB 肿瘤细胞的其他下游效应,发现相关生物过程的富集发生了一些变化(图 5B)。抑制 Wnt 和 MDK 后,与细胞凋亡相关的过程上调,细胞生长受到负调节。PKF 治疗后的其他变化包括髓系白细胞活化上调,细胞外基质组织和与肝脏代谢相关的通路(如磷脂酰胆碱代谢过程)下调。抑制 MDK 后,一些最显著的额外变化是与白细胞迁移和趋化性、髓系细胞分化或白细胞介素(IL)-2 产生相关的过程上调,而Wnt 信号通路和骨化以及其他发育过程的调节则下调。
为了进一步探索 MDK 在 HB 免疫调节中的作用,他们将 THP-1 衍生的巨噬细胞与 HB 细胞系共培养,HB 细胞中既有使用也有不使用 MDK 抑制(图 5C)。随后对分选出的巨噬细胞进行 RNA-seq,结果显示,与巨噬细胞单一培养物(图 5D)以及与与 MDK 抑制的 HB 细胞共培养的巨噬细胞(图 5E)相比,未经处理的共培养衍生的巨噬细胞中与 ECM 沉积和胶囊形成相关的通路上调。这表明 HB 肿瘤细胞的紧密接近促进了巨噬细胞潜在的促纤维化转变,并且如果肿瘤细胞中的 MDK 信号传导受到抑制,这种影响甚至会逆转。值得注意的是,这些生物学过程与本研究发现在肿瘤边界区域富集的过程非常相似(图 3G),许多髓系细胞位于该区域。
他们鉴定出几个参与 ECM 形成的单个基因,例如COL18A1、TGFBI以及TMPRSS6,它们表现出相似的表达模式:与未处理的 HB 细胞共培养后上调,但与 MDK 抑制的 HB 细胞共培养后不上调(图 5F)。具有相同模式的其他基因包括抗炎基因EGR1和S100A10,他们已在 scRNA-seq 数据中检测到它们在 HB TAM 中上调(图 2F),AMOTL2,作为机械传感途径的一部分参与抑制炎症;PDLIM1,NF-κB 信号抑制因子;IL18BP,可抑制IL-18 诱导的干扰素γ产生。
这些结果表明,MDK信号通路在Wnt活性增强的刺激下,参与塑造HB的免疫环境,其机制很可能是通过改变巨噬细胞的转录程序实现的。虽然这些变化可能具有直接的抗炎作用,但共培养巨噬细胞和肿瘤边界具有相似性,尤其是在ECM和包膜形成方面,表明存在其他间接效应,例如促进免疫逃避的ECM重塑(图5G)。

图5. Wnt 反应中期因子信号支持髓系驱动的免疫逃避。
(A) Wnt 信号通路评分与MDK表达之间的相关散点图。(B)GO分析。(C)实验设计。(D-E)GO分析。(F) 箱线图显示实验条件之间 THP-1 基因表达差异。(G) 肝母细胞瘤中Wnt和MDK信号传导之间可能存在的相互作用及其潜在下游效应的图形总结。
+ + + + + + + + + + +
结 论
在已鉴定的四个转录肿瘤程序中,“发育”和“代谢”反映了不同的肝细胞分化阶段,“循环”在未分化细胞和复发样本中富集,“中间”在预后不良患者的样本中表现出高活性。本研究发现抗炎免疫细胞与促炎免疫细胞的比例增加,并有证据表明肿瘤区域存在免疫排斥。肝母细胞瘤细胞中免疫调节剂中期因子的Wnt反应性上调与巨噬细胞表型的改变相关,而这种改变可以通过抑制中期因子部分逆转。HB细胞存在于连续的肝脏分化谱中,并位于改变的免疫环境中。Wnt信号增强中期因子的表达,而中期因子似乎通过修饰巨噬细胞使其能够逃避免疫反应来塑造免疫环境,从而为治疗提供了一个潜在的靶点。
+ + + + +



