

 English
English文献解读|Cell Metab(29.0):延长禁食时间与成年雌性小鼠的热量限制相结合可产生更大的 保护作用
✦ +
+
论文ID
原名:Prolonged fasting times reap greater geroprotective effects when combined with caloric restriction in adult female mice
译名:延长禁食时间与成年雌性小鼠的热量限制相结合可产生更大的衰老保护作用
期刊:Cell Metabolism
影响因子:29.0
发表时间:2023.07.11
DOI号:10.1016/j.cmet.2023.05.003
背 景
饮食干预是促进健康老龄化的重要策略。在大多数模型生物和哺乳动物中,热量限制(CR)可显著延长寿命并延缓与年龄相关的疾病和病理的发生。时间限制喂养 (TRF) 最初源于昼夜节律计时和调整,涉及将每天获取食物的时间限制在 8-10 小时或更短,而无需特别施加热量限制。大多数研究 CR 和 TRF 策略代谢反应的实验都是在雄性小鼠体内,然而,这些喂养方案对雌性动物的影响知之甚少。
实验设计
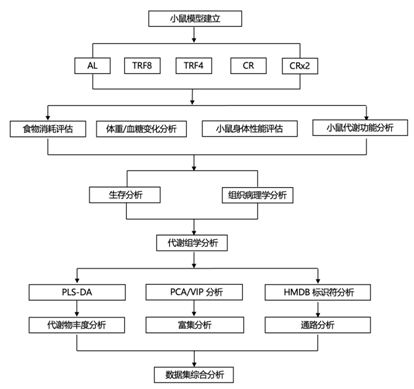
结 果
01

随意进食(AL)、TRF 和 CR 饮食小鼠的食物摄入量和体重 (BW) 变化模式
在对手动操作的7天转盘喂食器系统进行1周的驯化后,喂食AL的61周龄C57BL/6J雌性小鼠消耗恒定量的食物,同时在接下来的24周内保持其体重(BW),之后小鼠(现在为85周龄)开始消耗更多食物,同时体重减少(图1A-B)。食物消耗量在45周左右(106周龄小鼠)趋于平稳,体重继续下降。TRF8组小鼠和TRF4组小鼠分别在8和4小时内无限制进食。与AL对照组相比,两组在饮食转换后的前24周的进食量均减少了22% ~ 24%,但随后逐渐摄入更多的食物,在45 ~ 50周的干预期后接近AL小鼠的水平(图1B-C),同时体重稳步下降(在饮食转换后67周,体重下降了27%)。CRx2(2次进食CR)小鼠在当天晚些时候进行第二次进食,作为与CR动物配对的对照,但禁食时间缩短。由于AL组小鼠摄食量的逐渐增加,CR组的小鼠经历了更高程度的热量限制,从实验开始时的20%过渡到45周后的35%限制(图1C)。AL 组的小鼠体内脂肪增加(以体重百分比计),并在长达 30 周内保持稳定,然后在喂食 43 周时脂肪减少(图 1 D)。尽管与 CR 和 CRx2 相比,TRF4 和 TRF8 组小鼠的食物消耗显著增加,但 9 周后,饮食改变导致这 4个组的小鼠的脂肪均增加,随后逐渐减少脂肪量(图 1 D)。尽管小鼠消耗的食物量不同,但在对各种喂养干预的反应中,没有观察到身体成分的明显差异。
接下来,他们探索了饲喂方式对全身代谢和葡萄糖稳态的影响。在研究开始前9周(52周龄)和饮食转换后26周(87周龄)测量小鼠空腹血糖(FBG)。虽然各组小鼠在基线时的FBG相似,但到了第二个20周时,四种干预措施组中小鼠的血糖水平明显低于AL组(图1E)。
然后进行口服葡萄糖耐量试验(OGTT)以评估血液中葡萄糖的清除率。维持各自喂养方案 17 周的小鼠的血糖水平在口服葡萄糖后迅速上升,在 15 分钟时达到峰值,然后恢复到基础水平,而 AL 小鼠的下降速度要慢得多(图1E)。经过 39 周的干预后,与之前的小鼠组相比,峰值血糖总体下降(图 1F),曲线下面积 (AUC) 值更低(图 1G)。CR组的小鼠表现出比 AL 对照组显著更高的葡萄糖清除率(图 1 G)。
他们采用传统的旋转杆试验和倒置笼顶试验评估了饮食转换和相关成分变化对身体性能的影响。在笼顶试验中,维持CR和CRx2 方案持续39周的小鼠明显优于TRF和AL组小鼠,其中考虑了下落潜伏期、体重和重力来计算动量冲击能 (图1H)。饮食转换 80 周后,CR 和 CRx2 组的小鼠的虚弱指数 (FI) 显著降低,但 TRF 组小鼠没有显著降低(图 1 I),其中肌肉骨骼和耳/眼/鼻成分发生显著变化(图 1 J)。总体而言,这些结果表明 CR 组小鼠的关键表型指标有更大的改善,可能是因为 CR 组和 TRF 组之间的 CR 程度不同。
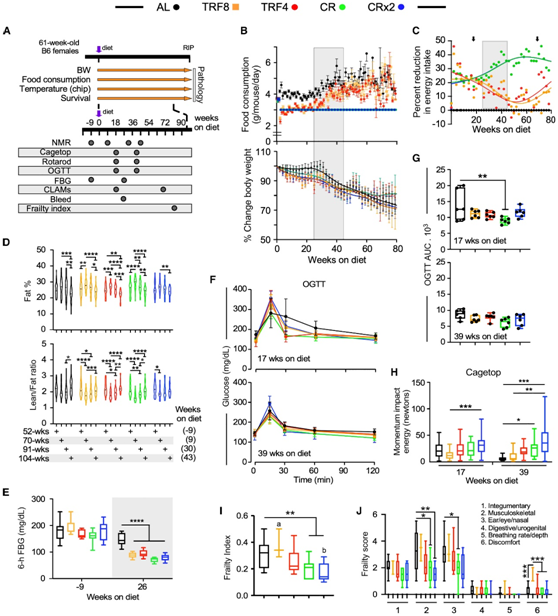
图1. 不同喂养方式对雌性 C57BL/6J 小鼠健康指标的影响。
(A) 测量指定生理参数的时间表。(B) 在指定喂养方案下 80 周内的食物消耗轨迹(上)和体重变化百分比(下)。(C) 饮食转换后 80 周内的热量限制程度。(D) 通过低场核磁共振成像评估的全身脂肪量百分比(上)和瘦脂比计算(下)。(E) 基线和饮食转换后 26 周的空腹血糖 (FBG) 水平。(F) 口服葡萄糖耐量试验 (OGTT) 期间血糖清除率的轨迹。(G) OGTT 实验的曲线下面积 (AUC)。(H) 来自倒置笼顶实验的冲击能动量。(I-J)记录总体衰弱指数评分和特定领域衰弱评分。
接下来,在饮食转换后17周和69周进行3天的实验,收集各种数据包括体重变化、食物摄入量、耗氧量(VO2)和产生的CO 2 (VCO2),计算呼吸交换比(RER)、能量消耗(EE),以及动态活动计数。
经过17周的干预后,AL 组的小鼠平均体重减少0.97 g,而TRF8、TRF4、CR和CRx2组小鼠的平均体重分别下降2.87 g、1.33 g、2.48 g和3.55 g,分别占其初始体重的8.4%、4.5%、7.8%和11.2%(图2A)。相反,经过69周的干预后,AL组小鼠平均体重下降了1.41 g(约为初始体重的5.3%),而TRF8、TRF4、CR和CRx2喂养分别导致小鼠平均体重下降了4.10g、4.12 g、4.57 g和4.50 g,分别占其初始体重的14.5%、15.4%、17.6%和18.9%(图2A)。无论干预时间如何变化,四组实验组小鼠的食物消耗都明显低于AL喂养的对照组(图2B)。摄食效率(定义为在代谢笼中体重相对于所摄入热量的变化)在17周期间不受饮食转换的影响,但只有CRx2组小鼠与AL组小鼠在69周内显著降低(图2C)。
为了确定不同喂养方案对全身代谢功能的可能影响,他们捕获了60小时内每小时的能量参数轨迹(暗期[D];亮期[L])。在 TRF8、TRF4、CR 和 CRx2 组小鼠与 AL 喂养对照组小鼠中观察到RER 的独特昼夜模式(定义为 VCO2 /VO2比率) (图 2 D)。经过 17 周的干预后,在 AL 组的雌性小鼠中观察到暗周期和光周期之间的 RER 波动最小,而将饮食改为 TRF8、TRF4 和 CR 会导致在上午 8:30 喂养时出现较大的 RER 波动(图2D)。由于CRx2小鼠在上午8:30和下午4:30进食了2次,他们注意到与CR小鼠相比,RER振幅更小,碳水化合物燃料利用的持续时间更短(图2E)。在69周的饮食转换后,他们观察到类似的轨迹,尽管AL和TRF小鼠随着年龄的增长摄入的食物比CR和CRx2小鼠更多,但RER波幅更小,持续时间更短(图2D)。
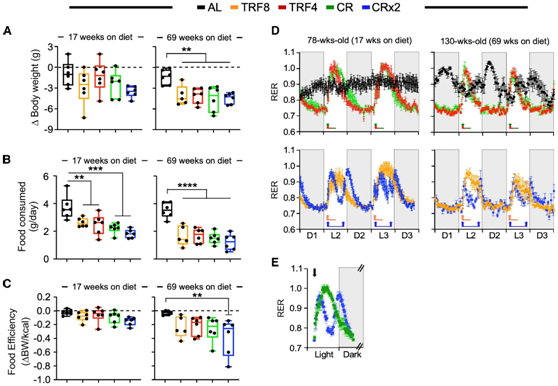
图2. 不同喂养方案对雌性小鼠体内代谢功能的影响。
(A) 小鼠进入和离开代谢笼时体重的变化。(B) 在代谢笼中的每日食物消耗。(C) 食物效率计算为小鼠在代谢笼中时体重变化与净能量摄入 (kcal) 之间的比率。(D) 在 17 周(左)和 69 周(右)接受所示喂养方案的小鼠中,在 3 个黑暗周期和 2 个光照周期内捕获平均每小时 RER 轨迹。(E) CR(绿色符号)和 CRx2(蓝色符号)小鼠在明相和暗相下的平均每小时 RER 轨迹。
02
生存研究和组织病理学分析
与 AL 对照组相比,CR 和 CRx2 方案处理的中年雌性小鼠的最大生存期显著增加(图 3 A)。将所有数据合并后,中位寿命的变化为3.2%。90%小鼠死亡年龄的平均增幅仅为2%(表1)。通过该试验,CR和CRx2与AL小鼠以及CR与TRF8小鼠相比,最大寿命显著增加。因此,CR和CRx2方案使雌性小鼠的寿命得到了小幅但显著的延长,并对其晚年产生了影响。
在所分析的四个器官(肺、肾、心脏和肝脏)中,每种病理的类型、患病率或严重程度均存在非统计学上的显著差异。在长寿TRF8和CRx2小鼠中,综合疾病负担评分(来自4个器官的综合平均评分)(图3B)和单个器官(包括心脏和肝脏)的评分分别显著高于AL对照(图3C)。在肺部,无论选择哪种喂养方式,淋巴网状内皮细胞瘤子(LRN) 和嗜酸性结晶性肺炎 (ECP) 是该组织中最普遍的病理特征。与 AL 组小鼠相比,CR 组小鼠表现出与 LRN 和总体肺损伤相关的最不严重的 ECP 形式(图3D)。这些结果表明,即使从中年开始,CR 干预也可以减少恶性肿瘤的进展和肺部炎症性疾病的严重程度。
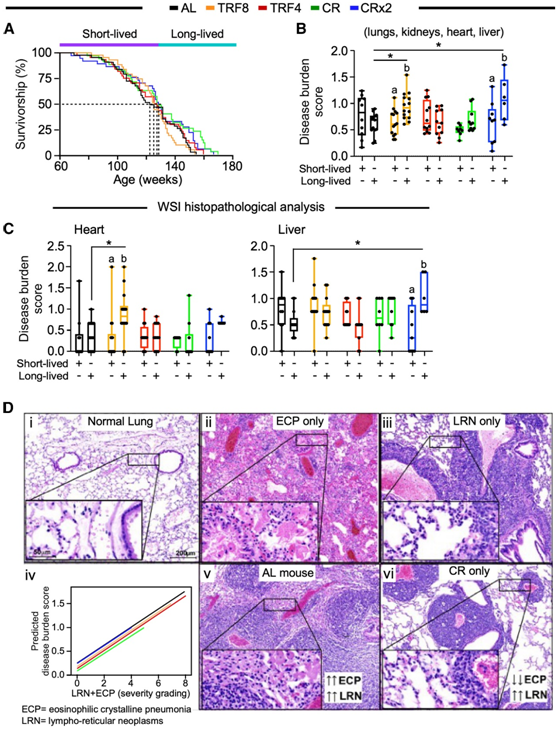
图3. 尸检时最常见肺部疾病的生存分析和组织病理学分级。
(A) Kaplan-Meier生存曲线。(B-C) 全玻片图像组织病理学分析。(D) 正常肺 (i)、患有嗜酸粒细胞性结晶性肺炎的肺(ECP, ii) 和患有淋巴网状肿瘤(LRN, iii) 的肺代表性图像。
表1. 最大寿命比较
03
代谢组学分析揭示了 TRF 和 CR 小鼠血清之间的不同特征
为了研究实验组之间健康寿命指标的差异是否与代谢重塑相关,他们对按各自方案喂养 26 周(87 周龄)的小鼠进行了非靶向血清代谢组学分析。
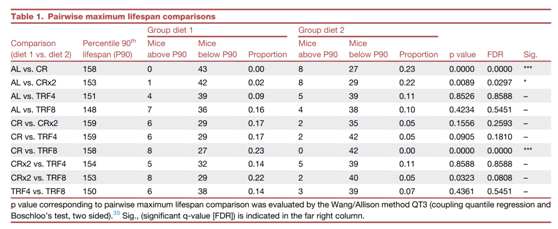
使用平均组强度热图可视化的聚类分析表明了依赖于喂养方案的特征。偏最小二乘判别分析(PLS-DA) 显示出明显的组分离,TRF4 和 TRF8 以及 CR 和 CRx2 之间存在显著重叠(图 4 A)。前25种代谢物的热图显示,与AL组相比,TRF组和CR组中有12种和13种代谢物的水平分别降低和升高(图 4 B)。前一组代谢物主要由外源物质(棕色箭头)组成,主要来自食物成分和氨基酸(深蓝色箭头),而后一组由各种脂类组成,包括酮体和饱和/不饱和脂肪酸(浅蓝色箭头)。
根据 PLS-DA,通过预测变量重要性 (VIP) 分析了分离的主要代谢物,并对前 25 种代谢物的重要性进行了排名(图 4 C)。其中,脂质种类是主要的有机化学物质 (19),其次是外源物质 (3),脂质种类与神经酰胺、碳水化合物(甘露醇/山梨糖醇)和外源生物同时呈负相关(图 4D)。这一模式与以下事实一致:在禁食期间,摄入的食物成分(外源物质)较少,而不同器官利用脂质和氨基酸作为底物。相比之下,TRF组与CR和CRx2组相比并没有显著的变化(图4B-C),这表明反应与摄入的热量有关。
使用四向维恩图,他们通过人类代谢组数据库(HMDB)标识符鉴定了四种喂养方案与 AL 之间的共有或唯一的脂质种类。在TRF4-AL或TRF8-AL配对比较中均未发现脂质类型,或者TRF4和TRF8 与 AL组之间没有共享的脂质类型,而CR-AL、CRx2-AL以及CR和CRx2 组与AL组的配对比较中分别发现了10、1和16个脂质分子的积累(图4E),且具有统计学意义(图4F)。将这27种代谢物(10 + 1 + 16)作为MetaboAnalyst(5.0版)的富集分析模块的输入数据,在CR和CRx2组(例如胆汁酸、脂肪酸酰基肉碱、二羧酸)和AL组(例如单酰基甘油、甘油磷脂)之间检测到显著差异(图4G)。然后使用相同的方法确定了4种喂养方案与AL相比具有共有或唯一的代谢物(脂质除外)(图4H-I)。CR(2/7)、CRx2(2/3)或CR和CRx2(3/13)与AL相比,对独特代谢物丰度的影响不大,而在TRF4和AL的比较中,超过7种代谢物独特富集,而TRF4和TRF8与AL相比有1种代谢物共同富集 (图4I)。化学修饰的氨基酸,包括甲基化、乙酰化和羟基化,占较大比例。富集分析表明,与CR组和CRx2组相比,AL组的Arg和Pro、谷胱甘肽以及Gly、Ser、Thr代谢相关的通路占优势,而TRF4组中与Arg和Pro、Trp代谢以及Val、Leu和Ile的生物合成/降解相关的通路的富集(图4J)。这些结果表明,CR组的血清代谢组学特征与TRF组不同。
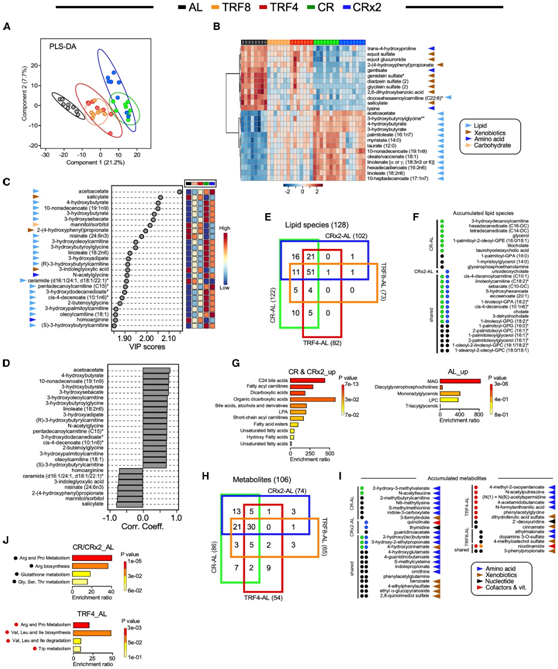
图4. 非靶向代谢组学揭示了 26 周饮食转换后雌性小鼠血清中 CR 的独特特征。
(A)偏最小二乘判别分析 (PLS-DA) 以二维图表示。(B) 热图描绘了通过单向方差分析和事后测试分析对每只动物识别的前 25 个特征的标准化值。(C) 导致喂养方案依赖性代谢差异的前 25 种血清代谢物的预测变量重要性投影 (VIP) 分数。(D) 无论喂养方案如何,前 25 个血清分子的相关系数呈正相关和负相关。(E) 具有人类代谢组数据库 (HMDB) 标识符的 128 种脂质种类的 4 向维恩图。(F)明显的脂质积累。(G)富集分析。(H) 106 种血清代谢物与 HMDB 标识符在所示配对比较中的 4 向维恩图。(I) 血清代谢物显著积累。(J)富集分析。
他们采用类似的分析方法直接评估 CR 和 TRF 组对禁食阶段的不同适应(比较CR 与 TRF4 以及CRx2 与 TRF8;禁食期分别为 20 小时和 16 小时)以及不同长度的影响摄取相同量食物时的禁食效果(比较TRF4 与 TRF8 以及 CR 与 CRx2)。四向维恩图揭示了每个配对比较的独特代谢物。CR-TRF4和CRx2-TRF8都有超过18个脂质,TRF4-TRF8和CR-CRx2分别只有1个和4个脂质(图5A-B)。富集分析证实了CR和TRF4(图5C)以及CRx2和TRF8(图5D)之间脂质特征的显著差异。两种CR方案均与可代谢为能量的脂质丰度增加有关(例如胆汁酸、二羧酸、饱和/不饱和脂肪酸、酰基肉碱),而TRF方案导致甘油磷脂、单酰基甘油、神经酰胺和一些酰基肉碱增加。随后,从这 4 次配对比较中鉴定出了独特的代谢物(脂质除外),CR-TRF4 和 CRx2-TRF8 分别具有11 种和 5 种代谢物,TRF4-TRF8 和 CR-CRx2 分别具有5 种和 3 种代谢物(图5E)。同样,在CR-TRF4的比较中,化学修饰的氨基酸占了富集的代谢物的很大比例,而异生物素、烟酰胺、γ-和β-生育酚是CRx2和TRF8中上调的代谢物(图5F)。
富集分析表明,与CR相比,TRF4导致Arg、Pro和Gly-Ser-Thr代谢相关的反应通路略有增加(图5G)。总之,这些数据表明,长期 CR 和 TRF处理后,由于 CR 程度不同,动物对与血清代谢组相关的禁食刺激的反应和适应不同(图 5 H)。此外,AL 和 TRF 似乎比 CR 更大程度地重塑甘油磷脂和鞘脂代谢。
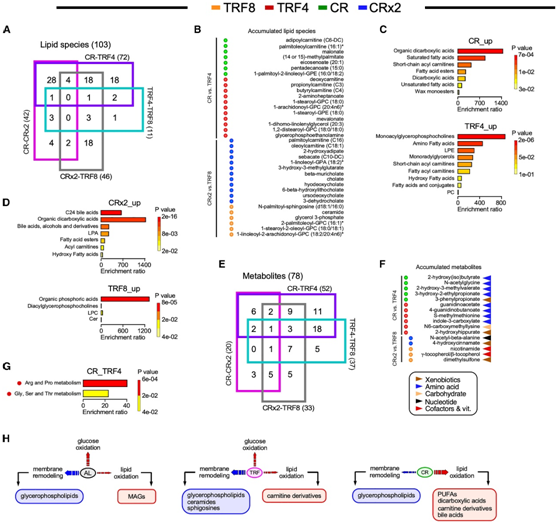
图5. 识别响应喂养方案的独特代谢组学特征。
(A) 具有 HMDB 标识符的 103 种脂质种类的四向维恩图。(B)血清脂质种类的显著积累。(C-D)积累的脂质种类的富集分析。(E) 78 种血清代谢物与 HMDB 标识符的 4 向维恩图。(F)血清代谢物显著积累。(G) CR 与 TRF4 中耗尽的代谢物的富集分析。(H) 图表大致描绘了喂食 AL、TRF 和 CR 的小鼠血清中不同的脂质代谢特征。
04
营养干预的系统影响及其代谢特征
为了评估 CR 和 TRF 营养干预对维持健康和最终生存的能量成本的系统影响,他们首先使用相关矩阵确定代谢物和表型结果之间的相关强度。其次,生成了整体极地热图,描述了来自营养干预、能量学和 FI 的血清代谢组特征的层次聚类。第三,通过主成分分析 (PCA) 量化了营养干预与 AL 方案的相对影响及其程度,具体取决于模式,即 CR、CRx2、TRF4、TRF8。
他们分析了血清代谢组特征与能量摄入百分比(图1C)、动量冲击能量以及FI的肌肉骨骼、皮肤和耳/眼/鼻成分(图1H-1J)之间的相关矩阵(图6A-6D)。
与TRF(即TRF4)相比,CR是产生代谢特征的影响最显著的干预措施,代谢特征由容易获得的脂质能量来源(如肉碱、PUFA、二羧基),而非膜重塑物质(如甘油磷脂、鞘氨醇)组成(图4B-H)。因此,他们将CR-CRx2和由15种上调代谢物组成的TRF4-TRF8的共享代谢特征(图4B)与上述生理指标相关联(图6A-6D)。
CR-CRx2 代谢特征中大多数脂质的负相关性表明 CR 程度越高,能量摄入量减少的百分比越高(图 1 C),导致代谢对脂质氧化的依赖性更高,从而证明它们在血清中水平的降低(图 6 A)。在CR-CRx2营养干预中,脂质代谢物丰度与动量能量再次出现负相关模式(图6B)。
与能量摄入和动量能量影响不同,肌肉骨骼(图6C)和耳/眼/鼻FI成分表现出明显的正相关模式,与TRF4和TRF8相比,CR-CRx2干预主要占主导地位。与之形成鲜明对比的是,FI的表皮成分(图6D)显示出相反的负相关模式,与其他FI成分不同的是,由TRF而不是CR干预主导。
所有5组的生理和代谢变量集合的具有层次聚类表示的极热图(图6E)和PCA(图6F)显示,PC1具有AL 61%的变异,而PC2具有28%的变异,对应于干预类型(即CR、CRx2、TRF4和TRF8)的影响(图6F)。这一结果与代谢组学分析(图 4和图5)以及极地热图中干预措施的聚类(图 6 E)一致,同时它有助于区分潜在的生理和血清代谢组变量(图 6 E-F)。
图6. 对各种数据集的综合分析,包括生理和代谢结果的功能特征。
(A-D)在每组营养干预中,14种代谢物显著变化的分层聚类相关系数与CR程度、动量冲击能量和虚弱指数(FI)成分(如肌肉骨骼和外皮)的热图。(E) 用圆形树状图描绘层次聚类的极坐标热图。(F)基于 5 个实验组的平均值的Z评分归一化生理和血清代谢组学数据的主成分分析 (PCA) 。
+ + + + + + + + + + +
结 论
本项研究将14个月的雌性小鼠置于5种不同的饮食方案中,即CR或TRF的不同喂养方案,并确定这些方案对生理反应、肿瘤和炎症疾病进展、血清代谢物水平和寿命的影响。代谢组学分析显示,与TRF喂养相比,CR引起了强大的全身反应,因为它与能量学和健康寿命指标(隔夜禁食动物的独特血清代谢组学特征)相关,并与寿命增加相关。这些结果表明,在生命晚期开始的延长禁食期的白天(休息期)进食与较低的能量摄入相结合时,产生了较大益处。
+ + + + +



