

 English
English文献解读|PNAS(11.1):磷酸胆碱诱导的能量来源转移可减轻由地理特异性 PM 2.5成分引起的肺细胞线粒体功能障碍
✦ +
+
论文ID
原名:Phosphocholine-induced energy source shift alleviates mitochondrial dysfunction in lung cells caused by geospecific PM2.5 components
译名:磷酸胆碱诱导的能量来源转移可减轻由地理特异性 PM 2.5成分引起的肺细胞线粒体功能障碍
期刊:Proceedings of the National Academy of Sciences.USA
影响因子:11.1
发表时间:2024.03.26
DOI号:10.1073/pnas.2317574121
背 景
空气污染,特别是空气动力学直径小于2.5 μm (PM2.5)的颗粒物(PM),是一种严重的环境风险,据世界卫生组织称,每年约有700万人死亡与此有关。PM 2.5由于其异质成分不断变化,可能导致许多疾病。因此,了解造成 PM 2.5总毒性的基本成分以及它们在不同城市如何相互作用非常重要。
实验设计
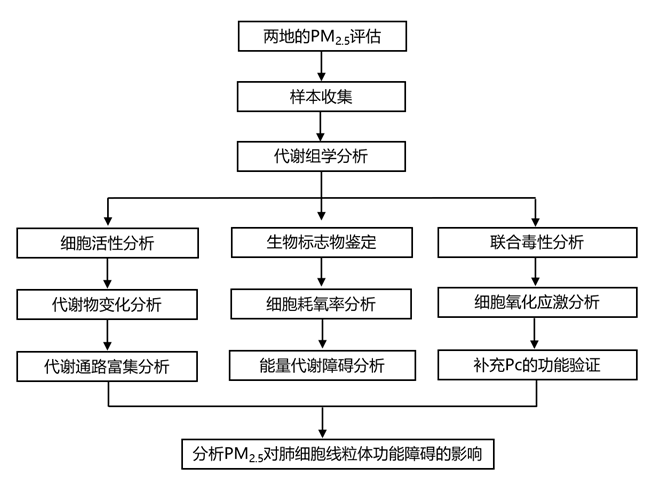
结 果
01

太原和广州的PM 2.5评估
采样期间,太原和广州的PM 2.5平均水平分别为57.7 μg/m 3和39.9 μg/m3,超过了中国推荐限值(35 μg/m 3)和世界卫生组织(10 μg/m3)。研究者团队鉴定并定量了太原和广州 PM 2.5样品中的 82 种化学物质,涵盖离子、金属、多环芳香烃(PAH)、硝基-PAH(N-PAH)、羟基-PAH (OH-PAH)和 多环芳香族硫杂环(PASH)。总体来看,太原无论是总浓度还是单项浓度均较高,且季节性波动较为明显(图1 A-B)。各污染物的质量比例主要由其类型决定,彼此之间差异很大,两个城市的浓度差异接近10个数量级(图1B)。诊断率显示,大部分(72%)太原市PM 2.5样本的FLT/(FLT+ PYR)和IcdP/(IcdP + BghiP)比值均大于0.5,说明燃煤是太原市PM 2.5的主要来源之一(图1C)。两个城市的平均增量终生癌症风险 (ILCR)水平都随着年龄的增长而升高(图1D)。
在两个城市收集的PM 2.5样本中,暴露于BEAS-2B细胞的细胞毒性试验得出了不同的结果。夏季、秋季和春季采集的太原PM 2.5样品对细胞进行挑战,细胞活力在86 ~ 109%之间,与暴露于广州PM 2.5样品后的细胞活力范围(78 ~ 112%)相似(图1E-F)。细胞活力与太原冬季PM 2.5样本量呈负相关(图1E)。太原特定日期PM 2.5的毒性要比广州相同浓度的样本强得多(图1E)。然而,通过超氧化物歧化酶(SOD)测定,PM 2.5暴露在BEAS-2B细胞中引起的氧化应激与污染物样品的氧化电位无关(图1 I-J),这表明PM 2.5的毒性主要取决于成分的生物利用度及其与生物系统的相互作用。
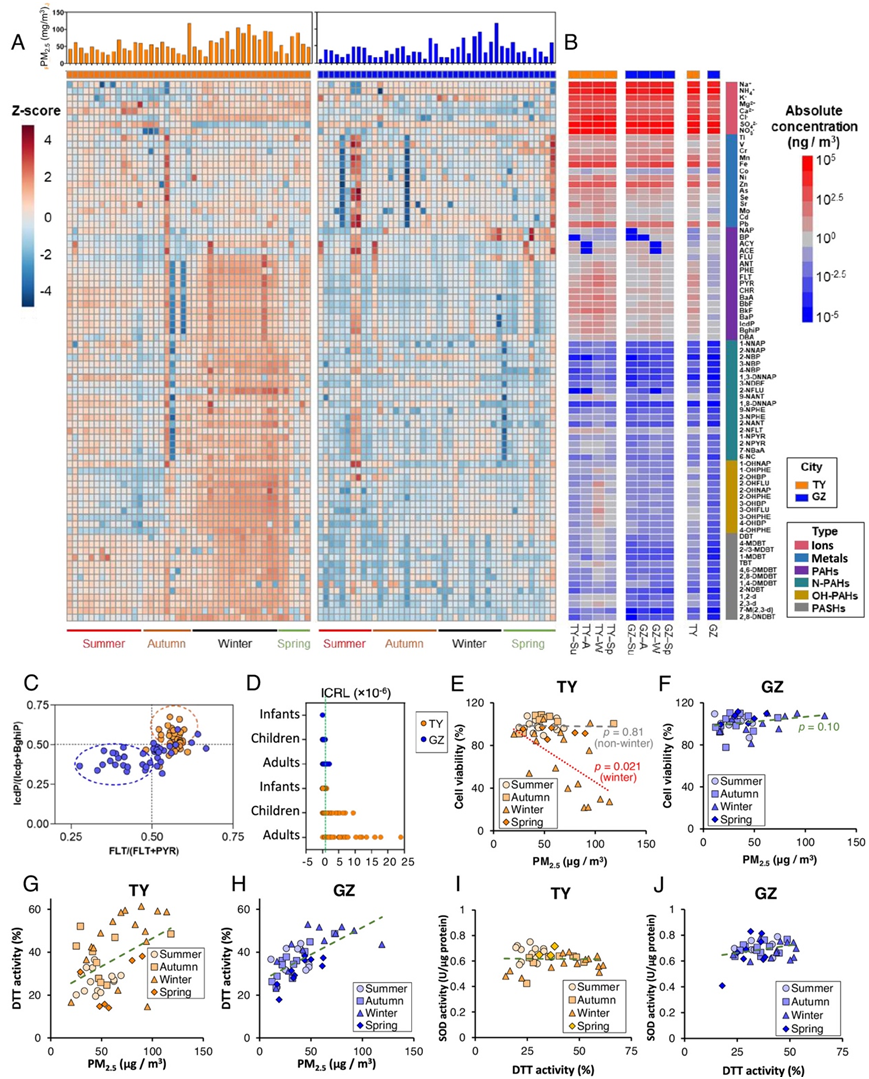
图1. 2017年至2018年收集的太原和广州全年PM 2.5概况。
(A) 每个样品中 82 种主要化学物质的 Z 分数。(B) 化学品绝对浓度的季节性和年度平均值。 (C)诊断率分析。 (D) ICRL 分析。(E-F) 太原和广州的细胞活力随 PM 2.5浓度的变化。(G-H) DTT 活性随PM 2.5浓度变化。(I-J) SOD活性随DTT活性的变化。
02
细胞毒性代谢组学分析
他们对暴露于太原和广州PM2.5样本的BEAS-2B细胞进行了整体代谢组学分析。BEAS-2B细胞是一种永生化的人支气管上皮细胞,已知其反应与原代肺细胞相似,已广泛用于PM2.5毒性研究。PLS-DA图(图S5 A-B)显示了太原和广州样品引起的不同的代谢紊乱,而来自每个城市的PM2.5引起了细胞的特定变化。虽然不完全相同,但两个城市的PM2.5引起的代谢组变化相似,太原的PM2.5引起的代谢紊乱更为明显(图S5 C-G)。总的来说,太原和广州样本分别显著改变了49种和33种代谢物(图S5H)。这些代谢产物在暴露于两个城市的PM2.5后发生了相同的变化,太原污染物总体上导致了更大的变化。所有代谢物的变化随采样日期而变化(图S5H),表明细胞代谢紊乱可能与PM2.5的剂量和成分有关。富集分析表明,与氧化应激(嘌呤代谢和谷胱甘肽代谢)和氨基酸代谢(如苯丙氨酸代谢、精氨酸和脯氨酸代谢、苯丙氨酸、酪氨酸和色氨酸代谢)相关的通路是显著的(图S5I-K)。相关分析鉴定出11个生物标志物与PM2.5成分(≥10个)显著相关(图2、图S6A-B),包括肌酸、谷胱甘肽(GSH)、磷脂(Pc)、6-磷酸果糖(F-6-P)、谷氨酸(Glu)、天冬氨酸(Asp)、尿苷二磷酸N-乙酰氨基葡萄糖(UDP-GlcNAc)、苯丙氨酸(Phe)、腺嘌呤、油酰胺、牛磺酸和N-乙酰-天冬氨酸(NAA)。值得注意的是,这11种细胞代谢物均未显示出与PM2.5总量的相关性(图2),这突出了PM2.5成分在毒性中的重要性。虽然PM2.5暴露导致38种代谢物增加,只有11种代谢物减少,但大多数代谢物在PM2.5处理后减少。
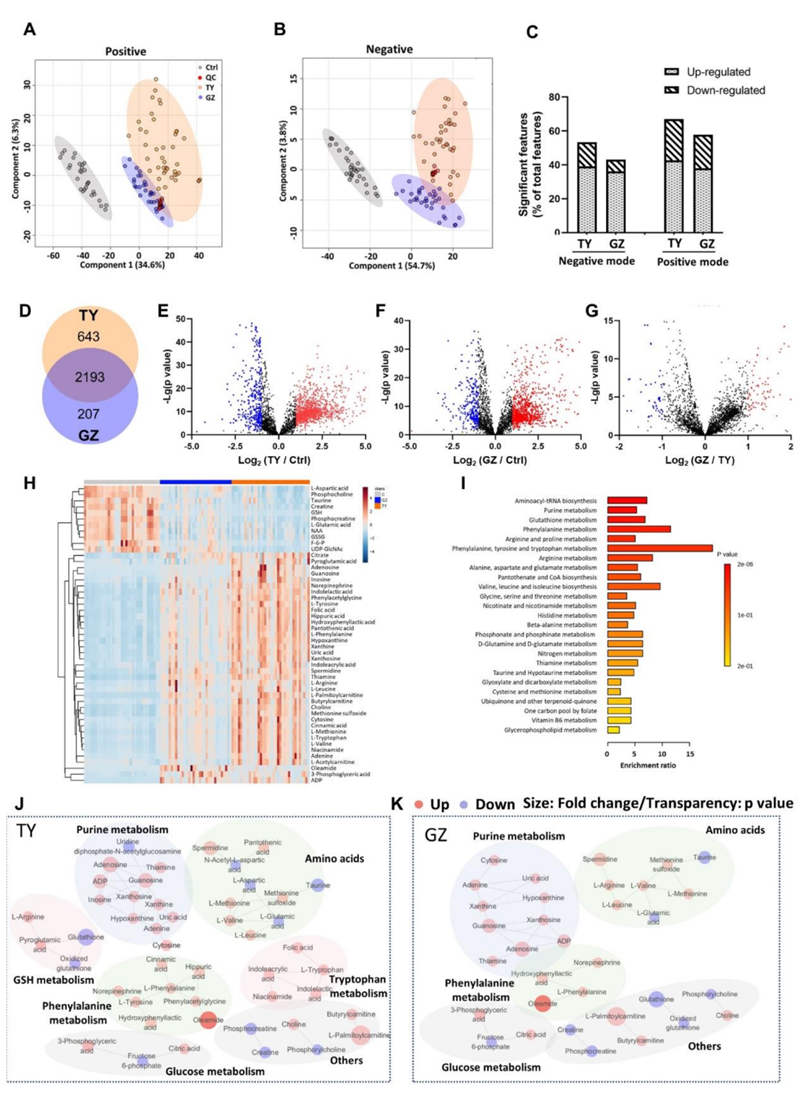
图S5. PM2.5暴露后肺细胞代谢组学分析。
(A)正模式的PLS-DA图。(B)负模式下PLS-DA图。(C)整体代谢组学分析检测出太原和广州PM 2.5暴露后BEAS-2B细胞中上调和下调的显著特征。(D)正负模式下不同特征数量的维恩图。(E-G)火山图显示代谢物丰度的变化。(H)太原和广州PM 2.5暴露鉴定的代谢物热图。(I) PM 2.5改变的显著通路富集分析。(J-K) PM 2.5暴露后BEAS-2B细胞的代谢网络。
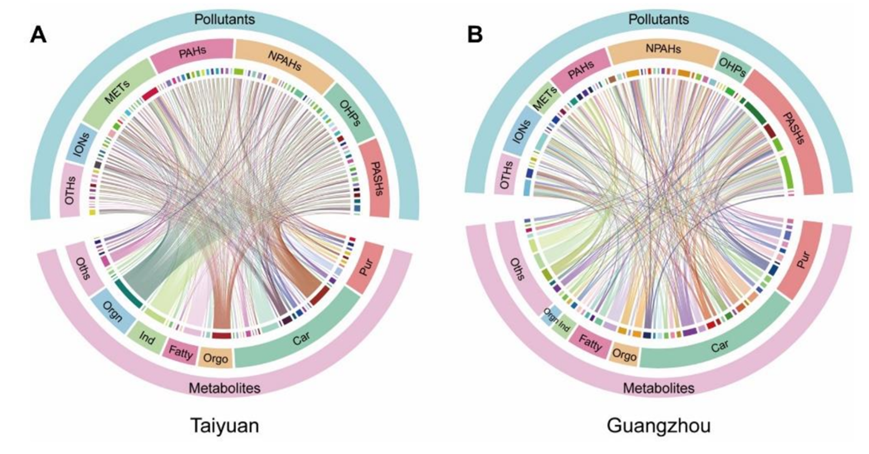
图S6. 鉴定的代谢物与测定的组分之间的相关性分析。
(A-B)样品中鉴定的代谢物与PM 2.5组分的相关性分析。
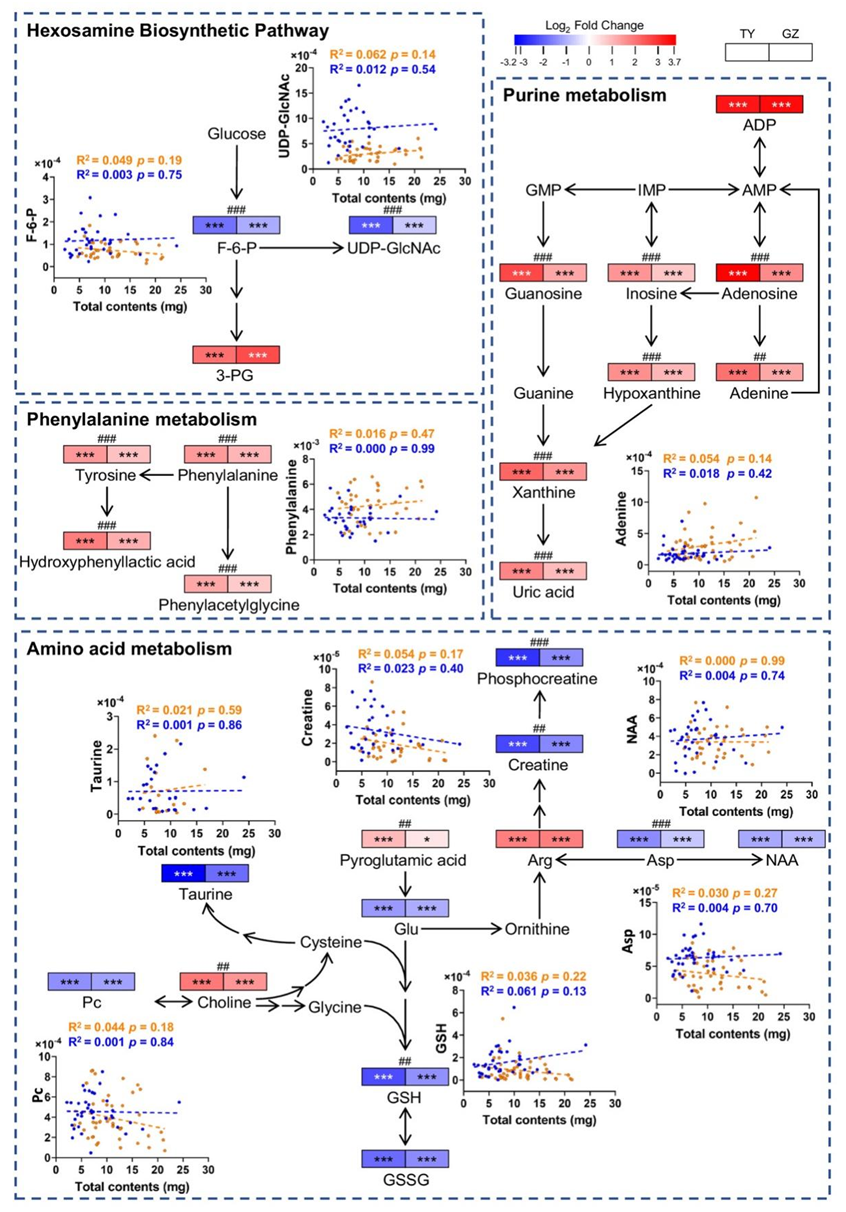
图2. 从相关分析中选择的生物标志物。
03
选定的代谢生物标志物在暴露于 PM 2.5后的细胞反应中的作用
然后,他们研究了PM 2.5暴露后对细胞代谢物的影响。补充Pc、牛磺酸和肌酸可以改善因 PM 2.5暴露而降低的细胞活力,表明这些化合物对 PM 2.5损伤具有保护作用(图 3 A-B)。人肺细胞线粒体是PM2.5细胞毒性的特异性靶点,已有研究证明了PM2.5侵袭后能量代谢对细胞存活的重要性。因此,他们评估了这些代谢物对细胞耗氧率(OCR)的影响。在正常生长条件下,这些代谢物都不会影响线粒体呼吸的效率(图3C-F)。然而,暴露于来自太原和广州的PM2.5后,Pc、UDP-GlcNAc和NAA可以显著增加细胞的基础呼吸、最大呼吸和三磷酸腺苷(ATP)的产生,表明这些代谢物在PM2.5阻碍的能量代谢中具有缓解作用(图3 G-N)。
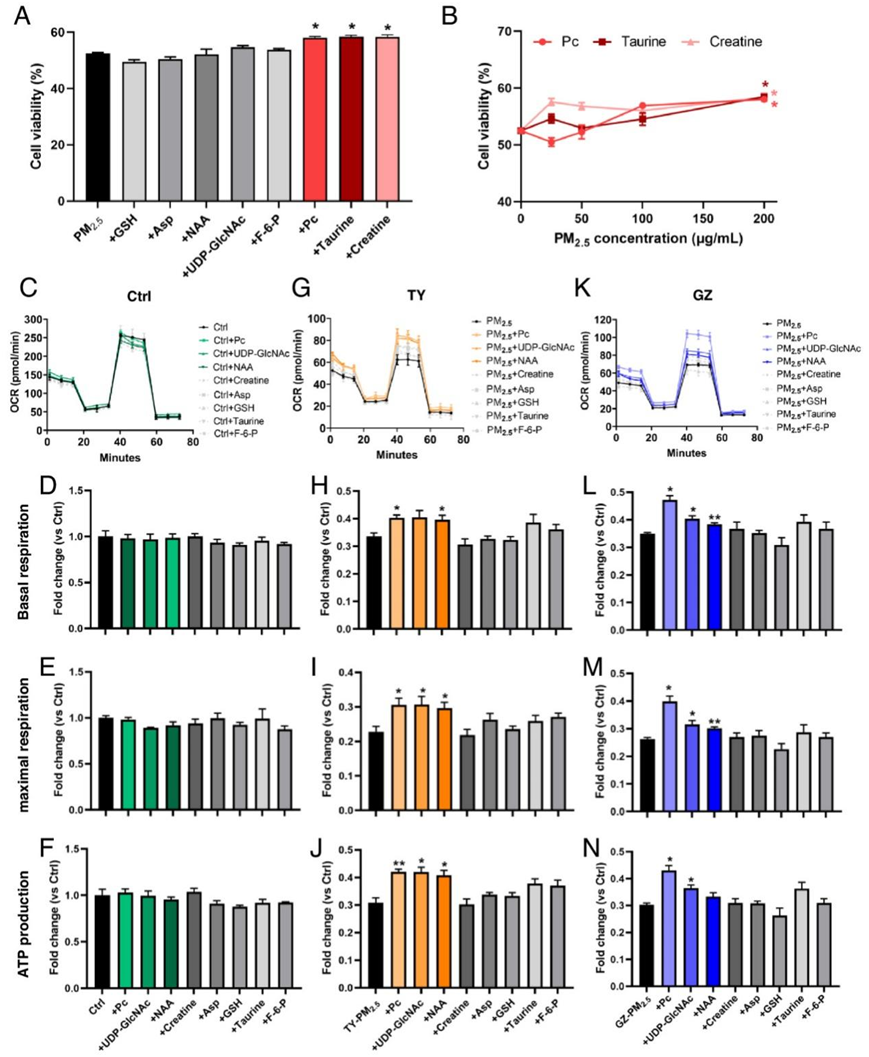
图3. 选定的代谢生物标志物在暴露于 PM2.5后的细胞反应中的作用。
(A)暴露于 200 μg/mL PM2.5样品的 BEAS-2B 细胞的细胞活力的影响。(B) BEAS-2B 细胞暴露于含有不同浓度 Pc、牛磺酸和肌酸的200 μg/mL PM2.5样品中细胞活力的变化。(C-F)OCR、基础呼吸差异倍数、最大呼吸差异倍数和 ATP 生成的差异倍数。(G-J) OCR、基础呼吸差异倍数、最大呼吸差异倍数和 ATP 生成差异倍数。(K-N) OCR、基础呼吸差异倍数、最大呼吸差异倍数和ATP产量差异倍数。
04
减轻功能生物标志物的影响
虽然PM 2.5具有抑制作用,但葡萄糖氧化抑制剂UK5099显著降低了细胞的最大呼吸,而脂肪酸氧化抑制剂etomoxir处理却没有发生明显改变(图 4 A-B)。这些结果表明,无论是在正常情况下还是在应对 PM 2.5毒性时,肺细胞主要使用葡萄糖而不是脂肪酸来提供能量。有趣的是,Pc的施用仅在暴露于PM 2.5的肺细胞中显著增加了最大呼吸,在此过程中,etomoxir处理也起作用(图 4 A-B)。这些数据表明,应用Pc可以将细胞代谢重新编程为脂肪酸氧化,以满足PM2.5暴露期间能量供应的不足。
相关基因的表达进一步证实了这些反应。L-谷氨酰胺:果糖-6-磷酸氨基转移酶(GFAT)是己糖胺生物合成途径中的限速酶,并初始化代谢为UDP-GlcNAc。暴露于PM2.5后,GFAT显著下调,而在细胞培养基中加入UDP-GlcNAc与GFAT的下降有相互作用。O-linked β- n -乙酰氨基葡萄糖转移酶(OGT)催化O-连 β-N-乙酰氨基葡萄糖(O-GlcNAc)对丝氨酸/苏氨酸蛋白残基的翻译后修饰。OGT的活性对其底物UDP-GlcNAc敏感。暴露于PM2.5后,OGT的表达也降低,并通过添加UDP-GlcNAc部分恢复。磷酸乙醇胺/磷酸胆碱磷酸酶1 (Phospho1)是控制Pc含量的主要酶。在正常细胞中,添加Pc对Phosoho1的表达没有影响。而暴露于PM2.5后,Phosoho1由污染物诱导大量上调,而Pc则显著抑制Phosoho1(图4C)。与此同时,调节氧化磷酸化的关键酶—过氧化物酶体增殖体激活受体γ辅助激活因子1- α (PGC-1α)的表达受到PM2.5的抑制,但其表达在Pc处理后恢复(图4D)。总而言之,这些结果表明,施用 Pc 确实将暴露于 PM 2.5的肺细胞的能量代谢转变为脂肪酸依赖性方式。能源的利用减轻了 PM 2.5引起的细胞功能障碍,这可能有利于细胞的生长。
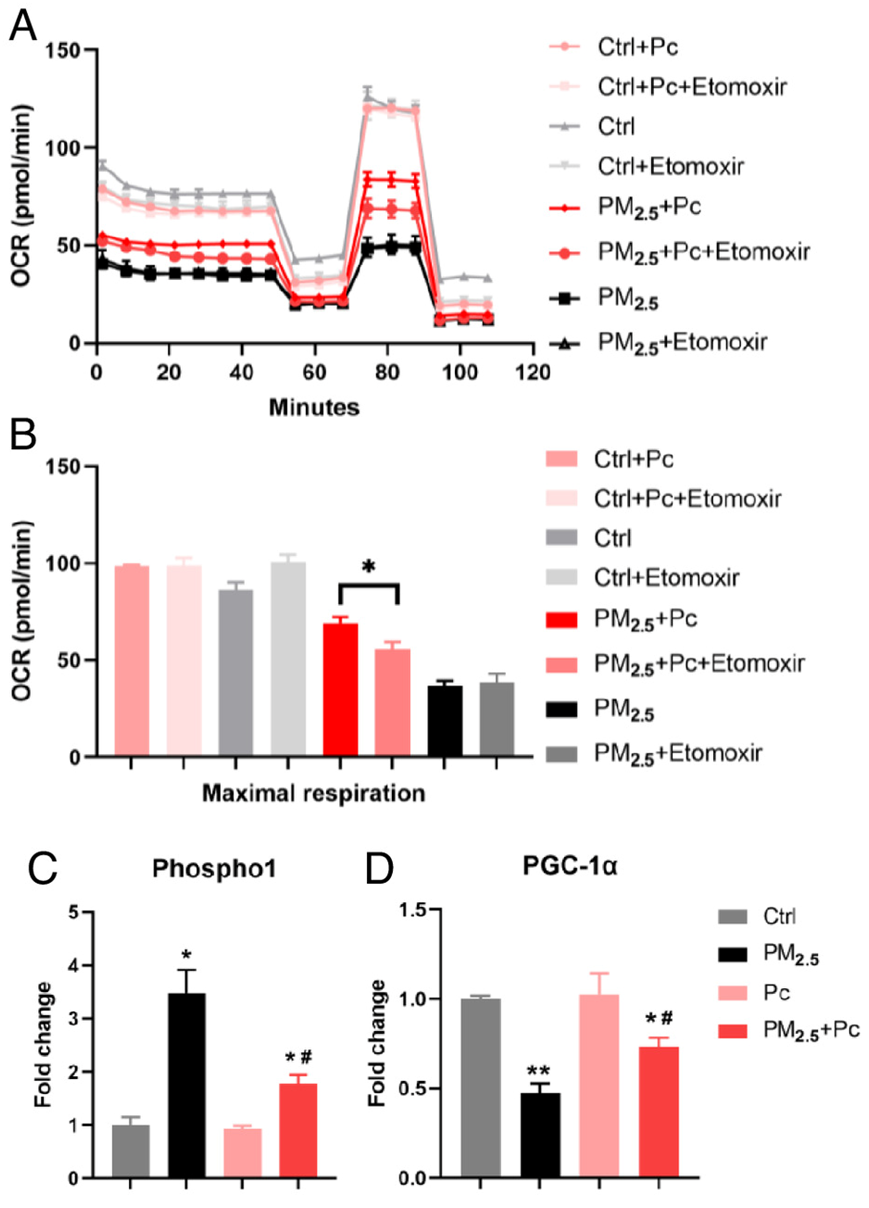
图4. 生物标志物Pc对PM 2.5能量代谢障碍的缓解作用。
(A)OCR脂肪酸氧化测试。 ( B ) 补充 Pc后,最大呼吸相对于正常细胞的差异倍数。(C-D)在有或没有PM 2.5的细胞中添加Pc后Phospho1和PGC-1α表达的差异倍数。
05
联合毒性分析
如上所述,PM2.5样品的氧化电位和毒性并不完全取决于PM2.5浓度,而PM2.5的成分,特别是在太原,起着至关重要的作用(图1、图2)。这些成分的毒性不仅取决于其成分的浓度,而且取决于它们的毒性当量。为了评估环境标志物对PM2.5毒性的贡献,他们探索了与代谢变化相关的PM2.5成分。根据相关代谢物的数量和PM2.5各组分的浓度,他们从太原PM2.5中选择了6个环境标志物,命名为TY1。根据它们在冬季的平均浓度,他们列出Na+ (30 mM)、Ti (50 μM)、FLT (25 μM)、2-NFLT (1 μM)、2-OHFLU (5 μM)和1,2-d (0.5 μM)进行毒性分析。他们测试了所有64种组合,每种组合由TY1中的一到六种污染物组成。所有六种污染物的混合物使细胞活力降低至72%(图5A),略高于太原冬季样品造成的平均细胞活力。综合毒性分析的总体结果表明,有机成分中的化合物在引起PM2.5的毒性中起重要作用。太原的单一环境标志物不会产生ROS,但当浓度相当于200 μg/mL PM2.5时,所有六种环境标志物的混合物会以剂量依赖性方式诱导细胞中ROS释放(图5 B)。
此外,补充Pc还显示出针对TY1对线粒体功能的不利影响的保护作用(图5C-D) ,与太原PM 2.5样品的结果相似。此外,用TY1的混合物替换太原PM 2.5样品导致Phospho1和PGC-1α的表达模式相同(图5E-F)。综上所述,这些结果表明,TY1 组对肺细胞具有与太原 PM 2.5类似的细胞毒性,可以通过额外补充Pc 来减轻这种毒性。
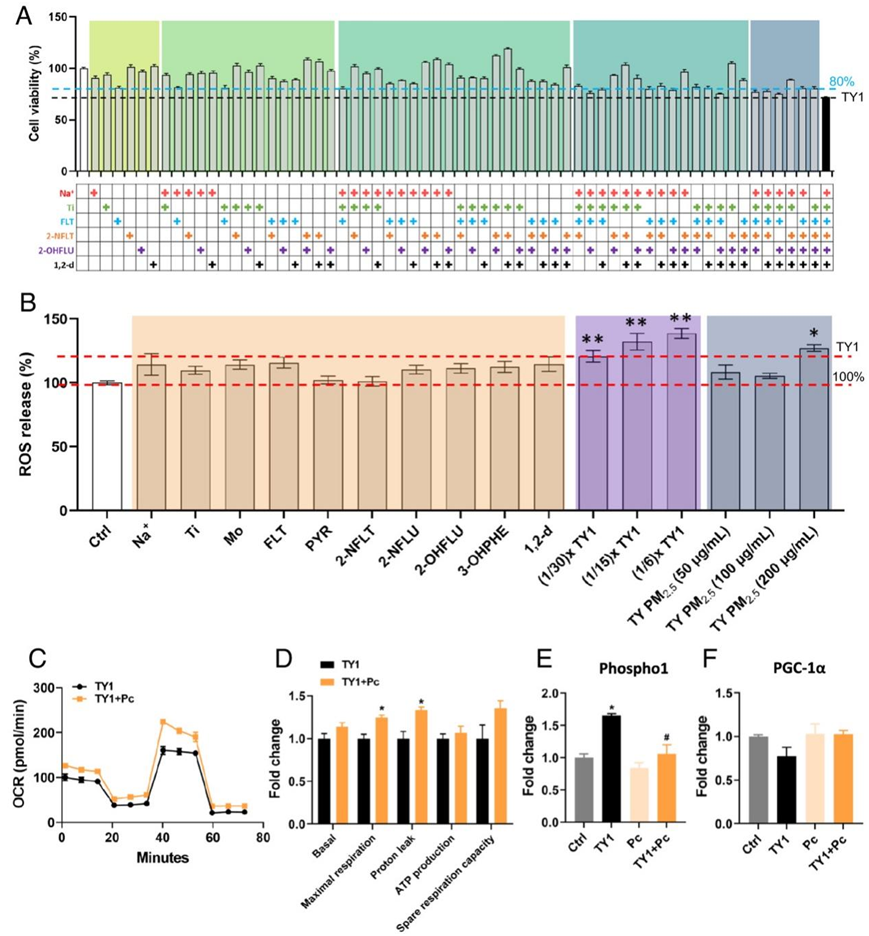
图5. Pc对暴露于太原PM2.5样本中环境标志物混合物的细胞的有益作用。
(A) BEAS-2B 细胞暴露于 TY1 组的各种组合后的细胞活力。 (B)暴露于不同组合后ROS的产生。(C) 供应 Pc 后暴露于 TY1 的细胞的 OCR。 (D) 提供和不提供 PC 的 OCR 实验的不同读数的差异倍数。(E-F)添加Pc后TY1暴露细胞的Phospho1和PGC-1α表达差异的倍数。
+ + + + + + + + + + +
结 论
本研究全面表征并比较了太原和广州两个地点的主要 PM2.5成分及其改变的代谢物。对长达一年的 PM2.5样本的分析揭示了 84 种主要成分,包括有机碳、元素碳、离子、金属和有机化学品。太原的PM2.5比广州的 PM2.5表现出更高的污染、相关的健康风险、二硫苏糖醇活性和细胞毒性。应用代谢组学方法,对暴露于两个城市PM2.5的 BEAS-2B 肺细胞进行了分析,揭示了两个地区PM2.5改变的代谢物以及PM2.5 的关键有毒成分。在PM2.5下调的代谢物中,磷酸胆碱有望成为PM2.5细胞毒性的干预靶点。它的补充通过激活脂肪酸氧化和抑制Phospho1表达,有效减轻PM2.5诱导的能量代谢紊乱和细胞死亡。本项研究提供了一种有前途的功能代谢物来缓解PM2.5引起的细胞紊乱,并提供了对有毒PM2.5成分的地理变异性的见解。
+ + + + +



