

 English
English文献解读|Nature(64.8):衰老人类骨骼肌的多模态细胞图谱
✦ +
+
论文ID
原名:Multimodal cell atlas of the ageing human skeletal muscle
译名:衰老人类骨骼肌的多模态细胞图谱
期刊:Nature
影响因子:64.8
发表时间:2024.04.22
DOI号:10.1038/s41586-024-07348-6
背 景
骨骼肌由大的多核肌纤维组成,具有不同的收缩和代谢活动(慢肌纤维/氧化性,也称为 I 型肌纤维;快肌纤维/糖酵解性,也称为 II 型肌纤维),由与肌纤维接触的运动神经元的活动控制。神经肌肉接头 (NMJ)。肌肉还含有多种数量较少的单核细胞,包括肌肉干细胞(MuSC、卫星细胞)、纤维脂肪祖细胞 (FAP)、脂肪细胞、成纤维细胞样细胞、免疫细胞、血管细胞和施万细胞。
肌肉萎缩和功能衰退(肌肉减少症)是虚弱的常见表现,也是老年人发病和死亡的关键因素。破译肌肉减少症的分子机制对于理解人类衰老具有重大意义。
实验设计
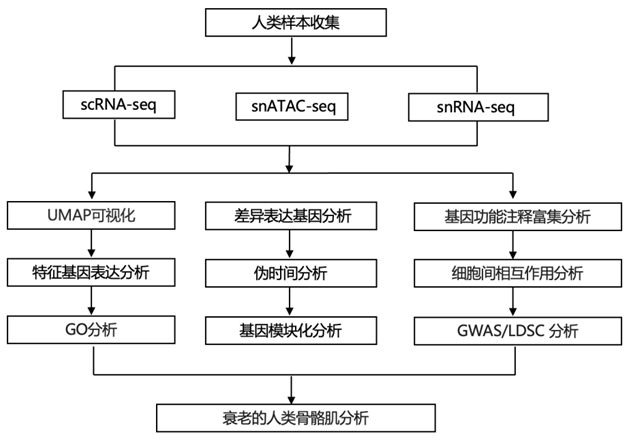
结 果
01

人体骨骼肌多模态图谱
为了研究人类骨骼肌随着衰老发生的分子变化,研究者团队对来自西班牙和中国的31名参与者(17名男性和14名女性)进行了后肢肌肉活检,分为两个年龄组:成年组(15至46岁)和老年组(74至99岁),男女年龄中位数分别为36岁和84岁(图1a)。将每个活检组织分为不同的样本,(1)用多聚甲醛固定进行组织学检测;(2)液氮快速冷冻单细胞核RNA测序(snRNA-seq)和单细胞核转座酶可及染色质测序(snATAC-seq);(3)单细胞RNA测序(scRNA-seq)。UMAP可视化显示了多核肌纤维室中代表I型和II型和特化肌核的聚类。在单核细胞中,主要的肌肉细胞类型是MuSC,基质细胞(FAP,成纤维样细胞和脂肪细胞),血管细胞(周细胞、平滑肌细胞和内皮细胞),免疫细胞(髓样,淋巴样和肥大细胞)和神经胶质细胞(雪旺细胞)(图1b)。对snATAC-seq数据的分析显示了对主要细胞类型的可靠鉴定(图1c)。广义线性混合模型显示,与年龄相关的 肌核减少,尤其II型肌纤维、MuSC和周细胞,以及与年龄相关的脂肪细胞、成纤维细胞样细胞和免疫细胞增加(图1d)。他们注意到大多数细胞类型在单个细胞/细胞核之间表现出转录异质性增加,这是衰老的一个新特征(图1e)。
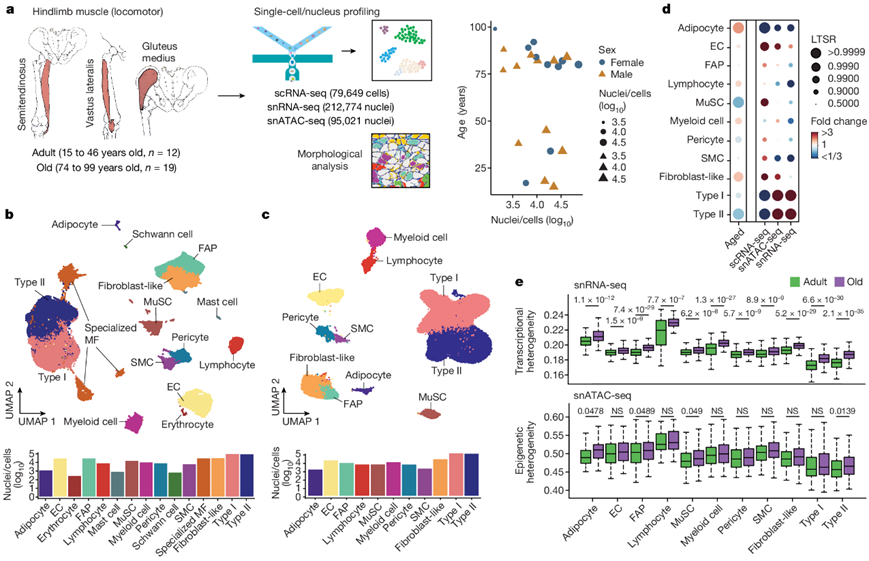
图1. 多模式人类运动骨骼肌老化图谱。
(a)本研究分析的后肢骨骼肌样本示意图。(b) sc/snRNA-seq 图谱的 UMAP 分析。(c) 基于已建立标记基因的基因活性评分。(d)每种细胞类型随衰老(第 1 列)和每种单细胞模式(第 2-4 列)的相对比例变化。(e) 按年龄组和细胞类型对转录(上)和表观遗传(下)异质性进行定量。
02
肌核组成的变化
他们根据已知的肌纤维类型特异性标记对snRNA-seq数据进行评分,分析不同年龄的肌核的异质性。除了MYH7+肌核(I型,TNNT1+)外,他们鉴定了两个已知的表达MYH2(IIA型)或MYH1(IIX型)的II型肌核(TNNT3+)亚型,以及同时表达两种MYH基因的杂合肌核(图2a)。随着年龄的增长,男性和女性的II型肌核减少,I型肌核相对增加,进而导致肌纤维的结构改变(图2b)。II型肌核的减少在IIX亚型中更为明显,其次是IIA/IIX杂交的肌核,其变化程度与个体的年龄高度相关(图2b)。
进一步的snRNA-seq亚聚类分析发现,在两种主要的肌纤维类型中,在肌腱连接处和NMJ存在特化的肌核(图2c)。肌腱连接肌核富集与细胞-基质相互作用相关的基因(COL22A1、ADGBR4),而NMJ肌核富集与突触传递反应相关的基因(PHLDB2、CHRNE)。重要的是,亚聚类鉴定了其他在成人或老年人肌肉中富集的细胞群体。例如II型肌纤维特异性ENOX1+肌核在成人组中富集。相反,TNNT2+、ID1+、DCLK1+和SAA2+的肌核在老年组中富集:(1)TNNT2+和DCLK1+的肌核主要存在于I型肌纤维中;(2) ID1+在两种类型的肌纤维中均有表达;(3) SAA2+群体,II型为主(图2c-d)。所有亚群均通过Hotspot分析得到证实,该分析将基因表达谱聚成模块(图2e-g)。
与NADPH氧化酶对骨骼肌的保护作用一致,ENOX1+肌核可能代表了健康的II型肌纤维群体,这是由快速收缩所需的碳水化合物代谢相关基因的高表达水平支持的(图2f)。心肌肌钙蛋白T (TNNT2)表达与去神经支配和衰老有关。TNNT2+肌核富集于与心肌收缩相关的基因(MYH6、TNNT2),提示骨骼肌肌小节特异性缺失。免疫荧光分析证实了TNNT2在老年肌纤维中的表达(图2h)。DCLK1编码双皮质素样激酶1,参与微管组装和动力学,并在营养不良再生(RegMyon)肌核中高度表达。ID1是一种转录因子(TF),参与与小鼠肌肉萎缩相关的BMP信号传导。血清淀粉样蛋白A2(由SAA2编码)是一种主要的急性期蛋白,在炎症和慢性组织损伤反应中高度表达。ID1+、DCLK1+和SAA2+老年肌核高水平表达NMJ相关基因(CHRNA1、CHRNG、MUSK、COLQ)和细胞黏附基因,如PCDHG基因家族成员,这可能表明对神经支配丧失的代偿反应(图2f)。
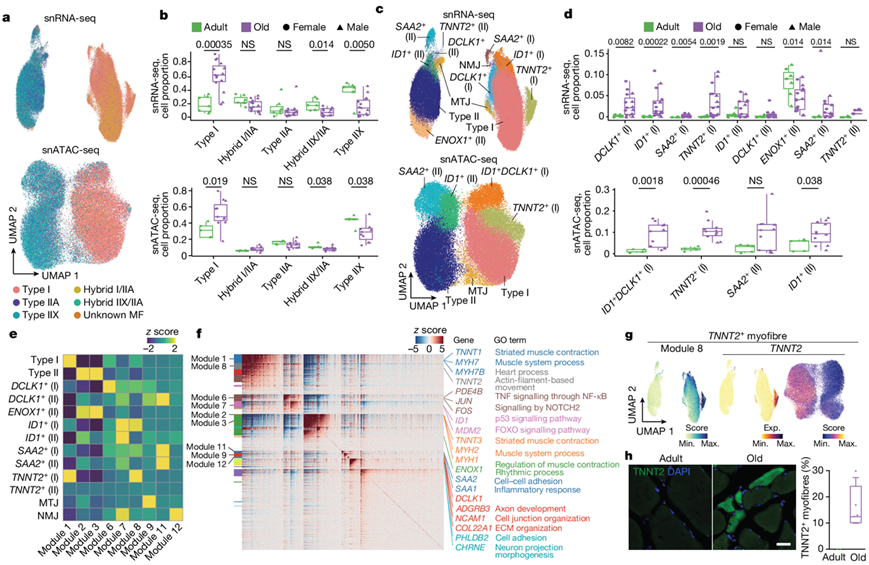
图2. 随着人类肌肉老化而出现的肌核群。
(a) 根据肌纤维类型特异性分类对 snRNA-seq(上)和 snATAC-seq(下)中的肌核进行 UMAP 分析。(b) 根据 snRNA-seq(上)和 snATAC-seq(下)中分类的肌纤维类型对成人(绿色)和老年人(紫色)个体的肌核比例进行定量。(c) snRNA-seq(上)和snATAC-seq(下)数据的肌核亚群的UMAP分析。(d)对成人和老年人中检测到的肌核亚群比例进行定量。(e) 每个模块中共表达基因的每个肌核群体中的缩放聚合表达水平。(f) 共表达模块的基因表达水平。(g) 模块8(左)和TNNT2基因表达(中)及其基因评分(右)的聚合表达水平的UMAP分析。 (h) 免疫荧光分析。
03
肌纤维类型特异性恶化
为了研究肌纤维转录变异的方向性,他们进行了伪时间细胞轨迹分析,观察了 I 型和 II 型肌核随衰老的确定路径(图3a)。这些肌核的轨迹终点与主要出现在老化肌肉中的新群体的转录谱相对应。当绘制受衰老逐渐影响的特定骨骼肌功能(分组为分数)时,例如肌节或萎缩相关基因(图3b),随着年龄的增长,I型肌核的变化轨迹是渐进的,而II型肌核的变化轨迹是突然的(图3a)。这种差异与 II 型肌纤维对衰老的敏感性更高相一致,这导致它们优先损失。相比之下,I 型肌纤维持续存在于衰老的肌肉中,并积累渐进性损伤,随着时间的推移进一步加剧肌肉功能障碍。伪时间的进一步分析显示了10个主要的转录变异聚类,其中大多数分别反映了I型或II型肌纤维的进行性或突发性变性过程(图3c)。例如,炎症小体(NFKB1、TXNIP),自噬(NBR1、ATG7)和氧化应激反应(SOD2、NFE2L2)基因的轨迹在I型肌核中稳步增加,但在II型肌核中急剧增加(聚类1)。在I型和II型肌纤维(聚类1、2和10)中,促萎缩Notch信号(HES1、NOTCH2)随着年龄的增长呈现相似的趋势。在I型肌核中,IL-6信号(IL6ST、SOCS3)的表达随着年龄的增长而明显上调(聚类3)此外,两种肌纤维类型均表现出增强的去神经特征(胆碱能突触;ITPR1、GNG12)(聚类10)。
作为最大的人体组织,骨骼肌是全身能量消耗的主要贡献者。线粒体对于维持骨骼肌纤维稳态以及通过氧化磷酸化和脂肪酸降解匹配能量产生至关重要。肌肉产生维持收缩的能量的能力随着年龄的增长而大幅降低,线粒体缺陷导致了这种现象,氧化磷酸化(IDH2、MDH1)和脂肪酸降解(ACADM、ACAT1)在I型肌核衰老轨迹(聚类8)中下调(图3d)。
为了检测肌核变性的顺式调节景观,他们使用基因调节的功能推理(FigR)定义了潜在的基因调节网络(GRN)。首先使用典型相关分析(CCA)整合了 snRNA-seq 和 snATAC-seq 数据集,在共同 CCA 空间内使用约束最优细胞映射方法识别最可能配对的细胞核,这在 snATAC-seq 中分别产生了 I 型或 II 型肌核向衰老状态一致的渐进或突然的细胞轨迹(图3e)。他们沿着衰老轨迹区分了 I 型或 II 型肌核中的数十种假定的转录激活因子和抑制因子(图3f)。其中,观察到与压力相关的转录因子(FOSL2、JUN、FOS、JUNB、STAT3)上调,并沿着退化轨迹驱动协调的基因表达程序。

图3. 肌核老化轨迹。
(a) snRNA-seq 数据集中 I 型和 II 型肌核群体的衰老轨迹(伪时间)的 UMAP 分析。(b) 肌核肌节评分的 UMAP 分析(上)以及显示 I 型(红色)和 II 型(蓝色)肌核沿着衰老轨迹的平均肌节评分的折线图。(c) I 型(红色)和 II 型(蓝色)肌核(左)沿衰老轨迹的基因聚类的模块评分。(d) 每个基因聚类的功能富集分析。(e) 从 snRNA-seq 数据转移的 snATAC-seq 数据集中 I 型和 II 型肌核老化轨迹的 UMAP 分析。(f) 沿着老化轨迹对I型和II型核进行每个TF的所有DORC的平均调节评分。
04
过早激活衰老导致的肌肉衰竭
他们鉴定了三个表达TF PAX7的MuSC亚群和一个表达MYOG的MuSC亚群。根据之前定义的标志物,PAX7+ MuSC在不同状态下进行亚分类:降钙素受体(CALCR)用于标记深静息状态MuSC(qMuSC);MYF5 用于标记早期激活的MuSC (epMuSC);以及中等水平的MKI67用于标记晚期激活/增殖的MuSC (lpMuSC)(图4a)。epMuSC富含FOS、JUN和EGR,这些已描述为MuSC在肌肉损伤后从静止状态快速退出。由于MYOG MuSCs富含肌源性分化基因(ACTC1),他们将其命名为分化型MuSC (dMuSC)。更广泛的DEG和功能富集分析表明,qMuSC富集了细胞外基质(ECM)重塑基因(FBN1、VIT、COL5A2和CALCR)和激素核受体(ESRRG、GHR)。
虽然MuSC异质性随着衰老而持续存在,但老年肌肉中epMuSC的比例有所增加(图4b)。与MuSC干性相关的通路(例如qMuSC的FOXO信号传导)和增殖能力(例如epMuSC的翻译和lpMuSC的细胞周期)随着年龄的增长而减少(图4c)。除epMuSC外,所有老年MuSC亚型均表现出线粒体氧化磷酸化增强。对来自成人和老年人群的qMuSC的详细分析表明,ECM相关过程(ITGBL1)的下调与年龄逐渐相关,而肌生成(MEF2D)的上调在74-82岁的成人qMuSC中达到峰值,而炎症和应激通路(TNF/NF-κB和NFAT-JUN-FOS)在≥84岁的成人qMuSC中达到峰值(图4c)。snATAC-seq分析表明,较老的qMuSC富集了调节晚期肌源性阶段的TF的结合基序,如分化相关(NFYA、NFYB、NFYC)和应激反应(ETS2、 EGR1)TF(图4d)。相反,生长激素相关转录因子(PGR、NR3C1、AR)的基序富集在qMuSC中随着年龄的增长而丧失。
接下来,他们对sc/snRNA-seq数据集中的单核细胞进行亚聚类。这些常驻细胞类型不仅对整体骨骼肌稳态至关重要,而且还支持损伤后MuSC的再生活动。在血管腔室中,他们发现了四种内皮亚型:(1)表达SEMA3G的动脉内皮;(2)表达CA4的毛细血管内皮细胞;(3)表达ACKR1的静脉内皮细胞;(4)表达IL6的venEC亚群(IL6+ venEC)(图4e)。在老年肌肉中,毛细血管内皮细胞和周细胞的比例下降,动脉内皮细胞和静脉内皮细胞的比例增加(图4f)。随着年龄的增长,血管细胞类型下调与细胞连接组装和跨膜转运体活性相关的基因,上调炎症(IL-6和AP-1通路)、纤维化(TGFβ通路)和自噬通路(图4g)。这些结果证明衰老通过增加促炎和压力相关信号改变骨骼肌血管完整性。
在免疫细胞中,他们从sc/snRNA-seq和snATAC-seq数据中鉴定了不同的髓系细胞和淋巴细胞亚群,包括CD14+和FCGR3A+ (CD16)单核细胞,它们对不同的病原体和刺激具有不同的反应;巨噬细胞[脂质相关巨噬细胞(LAM)和LYVE1+巨噬细胞]在骨骼肌中具有尚未明确的特殊功能;肥大细胞;树突状细胞;B细胞(幼稚和记忆);自然杀伤(NK)细胞;NK T细胞;CD4+ T细胞(效应性CCR7−,幼稚性CCR7+和调节性IL2RA+);CD8+(效应CCR7−和初始CCR7+);以及一组CCL20+ T细胞(图4h)。与组织学分析显示的炎症细胞浸润增加一致,肥大细胞、LAM和单核细胞在老年肌肉中增加,而一些T细胞亚型和树突状细胞减少(图4i)。除肥大细胞外的所有免疫细胞亚群,随着年龄的增长,下调了稳态免疫功能,包括抗原处理和呈递(MHC通路,B细胞受体信号和TCR信号)(图4j)。同样,在一些免疫细胞类型中,抗炎反应下调(淋巴细胞中ERBB4信号传导),而在其他免疫细胞类型中,促炎反应上调(髓细胞中IL-6通路,髓细胞和淋巴细胞中补体激活和NTRK1信号传导)。此外,老年免疫细胞在吞噬相关过程(内质网蛋白加工、网格蛋白介导的内吞作用和ECM降解)中富集,表明其处于主要激活状态。
在基质细胞中,他们鉴定了各种亚型的纤维成脂细胞(FAP),包括CD55+、CD99+、GPC3+、 MME+和RUNX2+,以及表达THY1的成纤维样细胞(图4k)。成纤维细胞样细胞随着年龄的增长而显著增加,而MME FAP则减少(图4l)。RUNX2 FAP富集SOX5,参与迁移和胶原生成。老年FAP亚型在很大程度上具有衰老特征,其特征是生长因子途径(VEGF和Wnt)的下调,促纤维化(TGFβ信号)和促炎症(IL-6信号)通路和天冬酰胺N-连接糖基化的上调(图4m)。这些结果表明基质群体(特别是CD55和成纤维细胞样细胞)向激活状态的转变,其特征是活跃的ECM重塑。
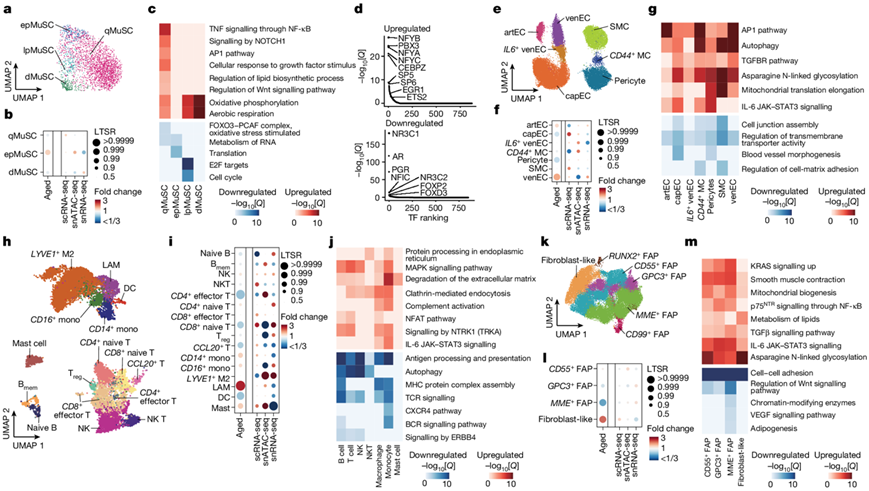
图4. 随着衰老,人类骨骼肌中常驻的单核细胞群。
(a) 对 snRNA-seq 数据中检测到的 MuSC 亚群进行 UMAP 分析。(b) 每个 MuSC 亚群随衰老(第 1 列)和每种单细胞模式(第 2-4 列)的相对比例变化。(c) 每个 MuSC 亚群的成人和老年人组之间获得的 DEG 的功能富集分析。(d) 随着衰老(老年人与成人),qMuSC 中上调(上)和下调(下)峰值的 TF 基序富集。 (e) 血管细胞亚群的UMAP分析。(f) 血管细胞亚群的相对比例变化。(g) 血管细胞亚群的 DEG的功能富集分析。(h) 免疫细胞亚群的UMAP分析。 (i) 免疫细胞亚群的相对比例变化。(j) DEG的功能富集分析。(k) 基质细胞亚群的UMAP分析。(l) 基质细胞亚群的相对比例变化。(m) DEG的功能富集分析。
05
细胞间通讯发生改变
与肌纤维相比,配体-受体相互作用主要涉及单核细胞,并且相互作用的总数随着年龄的增长几乎翻倍(图5a)。与其他细胞类型相比,涉及骨髓细胞和淋巴细胞的相互作用,以及较小程度的 FAP、成纤维细胞样细胞和 I 型肌纤维的相互作用,随着年龄的增长而显著增加(图5b)。基于这些结果,他们重点关注三个相互作用类别,作为衰老引起的肌肉萎缩过程的潜在效应器:炎症、ECM 和生长因子(图5c)。
主要的促纤维化因子TGFβ主要由免疫细胞(TGFB1)、MuSC (TGFB2)和I型肌纤维(TGFB2、TGFB3)产生,并通过其受体(TGFBR1、TGFBR2、TGFBR3)作用于多种细胞类型,特别是FAP,成纤维细胞样细胞,脂肪细胞和内皮细胞(图5d-e)。促萎缩激活素信号随衰老而上调,肌核中的激活素受体(ACVR1、ACVR1B、ACVR2A、ACVR2B)上调,EC、FAP、MuSC和成纤维细胞样细胞中的激活素配体INHBB上调(图5f-g)。免疫荧光分析验证了ACVR2A表达的增加(图5h)。值得注意的是,卵泡抑素(FST)在FAP中增加,可能是为了抵消激活素信号的促萎缩作用。
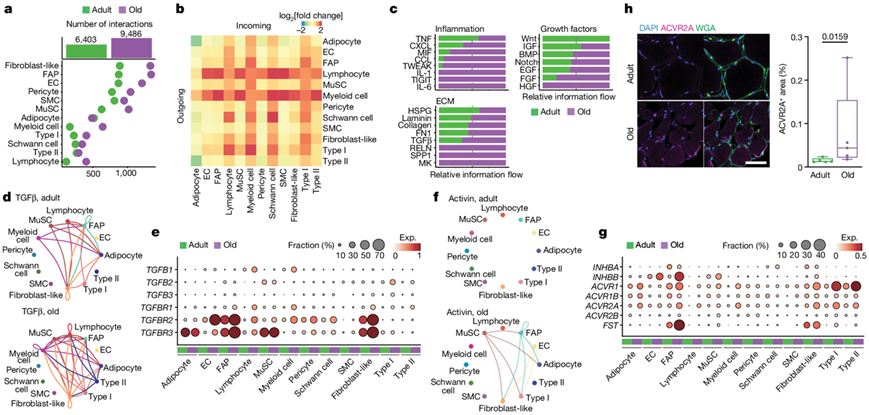
图5. 骨骼肌细胞成分的相互作用组分析。
(a) 成人(绿色)和老年人(紫色)年龄组中每种细胞类型的预测相互作用(L-R 对)的数量。(b) 每种细胞类型的发送信号(传出,水平侧)和接收信号(传入,垂直侧)数量随老化的差异倍数。 (c) 成人(绿色)和老年人(紫色)年龄组中每个描绘的交互组的所有对之间的交互概率差异(相对信息流)的总和。(d) 成人(上)和老年人(下)的 TGFβ 信号网络。(e) 成人(绿色)和老年人(紫色)肌肉中与 TGFβ 信号通路相关的基因的表达水平。(f) 信号网络。(g) 激活素信号通路和肌肉萎缩相关基因的表达水平。(h)免疫荧光分析。
06
将遗传风险变异与细胞类型联系起来
通过整合来自不同细胞类型和年龄组的所有细胞核的片段,他们生成了一个包含636363个峰的联合峰集,其中,从所有测试个体中鉴定出93565个富含单个细胞类型的峰(图S13a)。成年组和老年组在这些峰的开放度方面表现出相似性和差异,强调表观遗传改变可能是肌肉衰老和肌肉减少症的重要驱动因素。他们进行了连锁不平衡 (LD) 评分回归 (LDSC) 分析(图6a),虽然瘦体重(lean body mass)特征如预期在 II 型肌核中富集,但与肌肉力量相关的特征却出乎意料地在衰老的成纤维细胞样细胞和 FAP 中富集,但在肌纤维中却没有富集,遗传变异可以通过改变细胞间通信网络来促进肌肉减少症。老年人的腿部阻抗与 MuSC 高度相关,单纯跌倒导致的骨折与成人 MuSC 和老年 II 型肌核相关。此外,他们注意到睡眠时间、肌酐和空腹血糖与肌纤维相关,表明这些细胞群在昼夜节律调节和代谢调节中的潜在作用。
他们使用多层方法优先考虑从低手握力性状和瘦体重性状中提取的变异特征(图S13b)。他们将先导变异体和与细胞类型特异性峰高度相关的变异体重叠,确定了3158个候选变异体(图S13c)。其中,他们发现rs1862574位于肌纤维的GDNF位点,可能影响肌肉的神经支配。他们还观察到TRIM63的rs3008232 (MuRF1)、ANGPL2的rs1281155和FOXO1的rs571800667,这些是肌肉萎缩的关键驱动因素。接下来,他们使用deltaSVM框架来预测调控变异对TF结合的影响。在一个潜在的致病变异体(rs6488724)中,重叠峰位于参与肌肉生成的MGP启动子(图6b)。这种单核苷酸多态性(SNP)产生G至A突变,从而增加HSF2的结合亲和力,而HSF2参与肌小节伴侣蛋白的转录调节,以维持收缩器官。rs73746499位于FKBP5基因座的内含子区域,位于肌纤维中(图6c)。该 SNP 产生 A 至 G 突变,破坏雄激素受体的结合亲和力,雄激素受体是维持肌肉质量的关键 TF 之一。
本项研究人类骨骼肌衰老参考图集为老年人肌肉减少症增加和虚弱发展提供了一系列令人信服的综合细胞和分子解释(图S13d)。
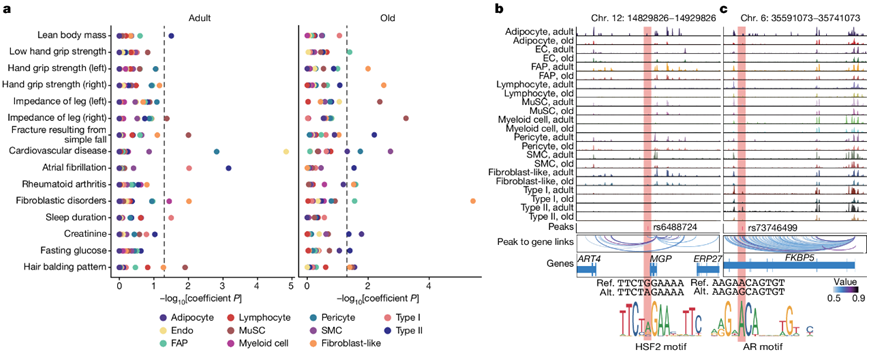
图6. 与肌肉减少症相关的遗传变异。
(a) 通过对成人(左)和老年人(右)年龄组之间每种细胞类型内映射的 snATAC-seq 峰进行 LDSC 分析获得的复杂性状的差异富集。(b-c) 基因组浏览器轨迹(上)显示了成人和老年肌肉中每个细胞群(行)与MGP位点的遗传变异rs6488724和FKBP5位点的rs73746499相关的标准化聚集信号。
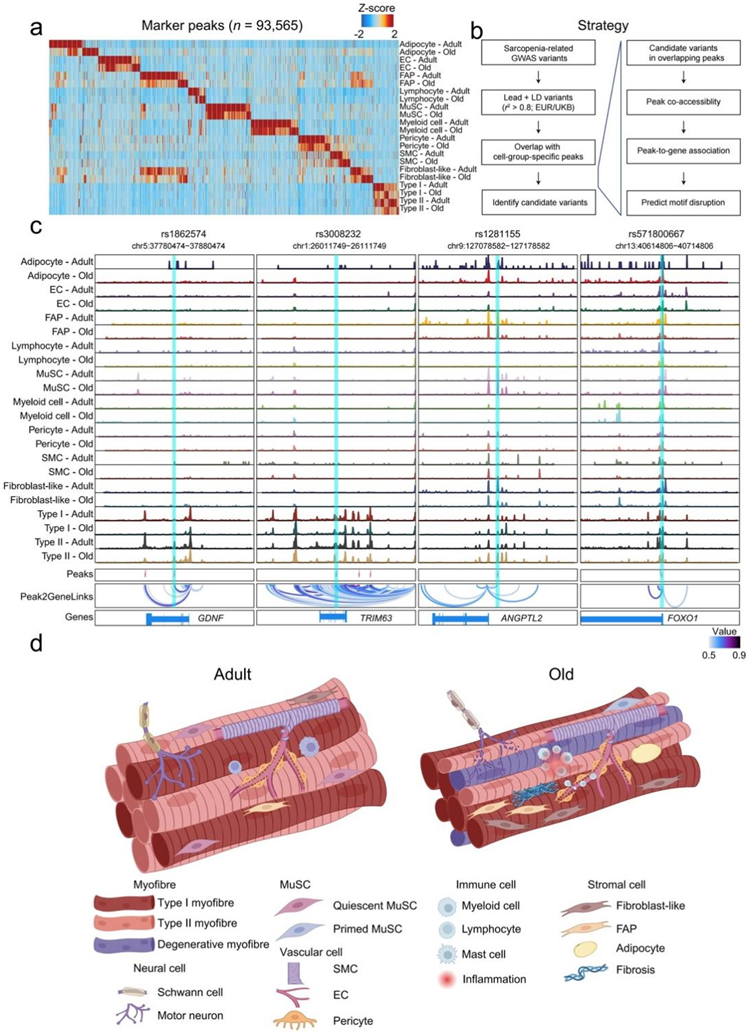
图S13. 衰老肌肉的全基因组关联研究 (GWAS)以及人体骨骼肌随衰老变化的空间分布。
(a) 通过 snATAC-seq 识别的每种细胞类型中衰老相关标记峰的热图。(b) 用于鉴定候选肌肉减少症相关遗传变异的方案。(c) 基因组浏览器跟踪成人和老年肌肉中的每个细胞群,显示分别与GDNF、TRIM63、ANGPTL2和FOXO1基因座内已识别的 SNP rs1862574、rs3008232、rs1281155 和 rs571800667 相关的峰值。(d) (成人)人类骨骼肌主要由合胞体多核II型和I型肌纤维组成,每种肌纤维均由单个运动神经元终板支配,其轴突由雪旺细胞细胞包围。
+ + + + + + + + + + +
结 论
本项研究生成了人类肢体骨骼肌的单细胞/单核转录组和染色质可及性图,其中包含来自 15 至 99 岁具有不同健康和虚弱水平的个体的超过 387000 个细胞/核,描述了细胞群在衰老过程中如何变化,包括老年组中新细胞群的出现,以及与这些变化相关的细胞特异性和多细胞网络特征(在转录组和表观遗传水平)。在与遗传数据交叉比较的基础上,本项研究还确定了标记肌肉减少症易感性的染色质结构的关键因子,为确定骨骼肌中适合晚年医学、药理学和生活方式干预的靶标提供了新的理论依据。
+ + + + +



