

 English
English文献解读|Cell Metab(29.0):小鼠急性寒冷应激期间代谢通量的综合量化
✦ +
+
论文ID
原名:Comprehensive quantification of metabolic flux during acute cold stress in mice
译名:小鼠急性寒冷应激期间代谢通量的综合量化
期刊:Cell Metabolism
影响因子:29.0
发表时间:2023.09.27
DOI号:10.1016/j.cmet.2023.09.002
背 景
生热作用使包括哺乳动物在内的温血动物能够适应寒冷环境并在寒冷环境中生存。环境温度下降时,生物体会经历复杂、协调的反应,通过内部产生热量来维持体温。寒冷诱导生热(CIT)由两个不同的过程介导:颤抖和非颤抖生热。颤抖产热通过骨骼肌的快速收缩和松弛释放热量。非颤抖产热主要发生在棕色脂肪组织(BAT) 中,通过 UCP1 介导的线粒体质子梯度解偶联以及其他机制产生热量。
目前对驱动 CIT 的代谢变化缺乏透彻的了解。
实验设计
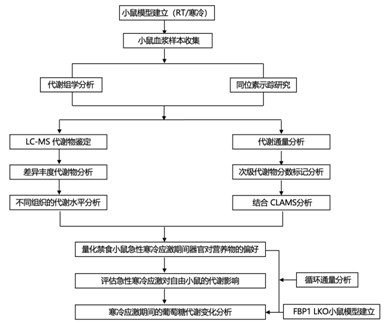
结 果
01

寒冷应激会引起 BAT 以外的广泛和全身代谢变化
为了评估寒冷应激对全身代谢的影响,研究者团队首先对来自未麻醉活体小鼠的动脉血浆样本进行了非靶向代谢组学分析,小鼠在室温 (RT) 或 4°C 下不进食,饲养 6 小时。使用液相色谱-质谱 (LC-MS),总共测量了 605 种代谢物。其中,在冷应激小鼠的血浆中,153 种上调,164 种下调(图 1A-B)。寒冷应激导致游离脂肪酸 (FFA) 水平大幅增加,尤其是不饱和 FFA(图 1 C)。特别是,最丰富的脂肪酸种类棕榈酸酯(C16:0)、油酸酯 (C18:1) 和亚油酸酯(C18:2)各增加了 55%–70%。相比之下,寒冷应激会导致溶血磷脂和甘油三酯水平下降(图 1 D)。在寒冷应激期间诱导最强的代谢物是多不饱和单酰基甘油,其中值得注意的是MG(20:4),也称为2-花生四烯酸甘油(2-AG),它是大麻素受体的内源性配体(图1E)。下丘脑中2- AG介导的大麻素受体信号通路抑制产热和能量消耗,其升高提示其可能是CIT的反馈抑制因子。氨基酸水平的变化各不相同,其中一些增加,特别是丙氨酸和支链氨基酸(BCAAs:亮氨酸、异亮氨酸和缬氨酸),而其他许多氨基酸则减少(图1F)。
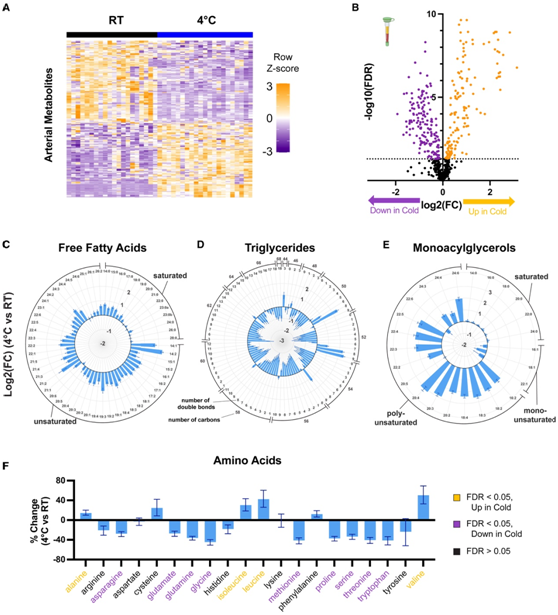
图1. 寒冷应激引起广泛和全身的代谢变化。
(A) 差异丰度代谢物的热图。(B) 火山图比较 4°C 或 RT 下小鼠血浆代谢水平。(C–E) 与RT相比,4°C 下小鼠血浆中个体游离脂肪酸、甘油三酯和单酰甘油的变化。(F) 与 RT 相比,4°C 下小鼠血浆中单个氨基酸的百分比变化。
接下来,他们对类似地饲养在室温下或急性应激于 4°C 的小鼠的 6 个不同器官[BAT、心脏、肝脏、股四头肌 (quad)、膈肌和性腺白色脂肪组织(g-WAT)]进行了非靶向代谢组学分析。寒冷应激会在所有研究的器官中引起显著的代谢变化,而不仅仅是像BAT这样的典型产热器官,而且这些变化在不同组织中差异很大(图 2 A-G)。在所有组织中,只有两种代谢物——苯丙氨酸和 3-羟基丁酸 (3HB)——因寒冷而显著增加,并且在所有组织中均未减少。除 BAT 外,所有器官中的FFA 水平均普遍升高,这与其循环上升和 BAT 中的剧烈消耗一致。柠檬酸盐仅在BAT中增加,这与BAT三羧酸(TCA)循环在冷作用下的强激活一致。肝脏中必需氨基酸普遍升高,提示寒冷诱导显著的肝脏蛋白质水解。综上所述,这些数据表明小鼠对寒冷应激存在协调、广泛和复杂的全系统代谢反应。

图2. 响应冷应激的跨器官代谢重组。
(A-F)火山图比较小鼠在4°C和RT下 BAT、心脏、肝脏、股四头肌(Quad)、膈肌和性腺WAT (g-WAT)中的代谢物水平。(G)差异倍数(FC)和错误发现率(FDR)。
02
量化禁食小鼠寒冷引起的全身碳通量变化
急性寒冷应激会显著增加全身代谢率。为了提供营养以维持这种升高的代谢率,寒冷应激的生物体会激活不同器官的各种代谢程序,包括脂肪组织中的脂解作用、肝脏中的糖异生和糖原分解作用、以及肌肉中的蛋白水解作用。肝脏中的糖异生和糖原分解作用。然而,缺乏对这些通路相关代谢通量和代谢率的定量评估。为了解决这个问题,他们在小鼠体内建立了双线体内稳态重同位素标记方案(图3A)。将微量的均匀标记的13C燃料分别静脉注射到清醒、自由活动的小鼠(分别在RT条件下和在4°C下)中,同时从动脉导管中采集间歇性血液样本。
为了全面了解寒冷引起的代谢通量变化,他们选择了多种丰富的营养素,包括碳水化合物(葡萄糖和乳酸)、氨基酸(谷氨酰胺和支链氨基酸)、脂肪酸(棕榈酸酯)和酮体(3HB),还计算了每种营养物的原子加权循环碳通量 (其碳原子出现的平均速率)(图3B),观察到的冷响应通量变化与血浆丰度的变化部分一致(图 3 C)。葡萄糖、乳酸、支链氨基酸、棕榈酸盐和 3HB(但谷氨酰胺除外)的循环通量因冷应激而增加,尤其是棕榈酸盐和 3HB。支链氨基酸(必需氨基酸)循环通量的增加反映了系统性蛋白水解作用,棕榈酸酯的循环通量则反映了系统性脂肪分解作用。
寒冷应激会促进糖酵解(葡萄糖到乳酸)通量的大幅增加,以及糖异生底物乳酸和谷氨酰胺产生的葡萄糖的增加。他们还观察到多种来源的 3HB 产量有所增加,包括棕榈酸等脂肪酸和 BCAA 等氨基酸(大概是亮氨酸和异亮氨酸,与缬氨酸不同,它们是生酮的)(图3D)。
接下来,它们将计算出的营养碳通量总和与通过综合实验动物代谢系统 (CLAMS) 测量的全身碳代谢(VCO2) 进行了比较。他们利用已发表的比值,从棕榈酸和BCAAs的测量通量推断出未测得的脂肪酸(FA)和蛋白质释放氨基酸的通量,从而生成RT和急性寒冷应激期间的全身终产物(EP)通量模型(图3E)。估计的累积EP通量高于测量的VCO2,表明某些营养素的非氧化命运(例如合成代谢)。在禁食状态下,完全合成代谢是不可能的,因此,这可能反映了通过细胞内大分子库进行的大量营养循环,例如,脂肪酸酯化为甘油三酯,并伴随这些物质的脂解。相反,寒冷诱导的EP通量上升与CO2生成的增加精确匹配(图3F),这意味着寒冷诱导的养分通量完全定向于氧化,而没有增加大分子循环。EP 通量的增加主要是脂肪酸燃烧,这表明脂肪酸是应激在寒冷环境中、禁食的小鼠的主要产热“燃料”。

图3. 量化禁食小鼠寒冷引起的全身碳通量变化。
(A) 演示13 C-代谢物输注系统的模型。(B-C)在室温(RT)或急性应激于4°C而不进食的小鼠中,动脉血浆中关键代谢物的循环碳通量和代谢物丰度。(D)选择循环养分之间的绝对通量,单位为nmol C/min/g。(E)小鼠在RT或4°C时关键代谢物的终产物通量率。(F)寒冷诱导的最终代谢通量率和VCO2的增加。
03
量化禁食小鼠急性寒冷应激期间器官特异性“燃料”偏好
接下来,为了研究寒冷应激如何影响单个器官的营养素使用,他们检测了每次标记营养物输注后不同组织中TCA循环代谢物的13 C 分数富集,以计算源自每种营养物的 TCA 循环碳分数(图4A-F)。
已有研究证明,在禁食动物中,许多组织不直接使用葡萄糖来为 TCA 循环提供燃料,并且在本项研究的数据中,直接葡萄糖氧化仅限于高度氧化的组织,如 BAT、心脏和红色肌肉(如隔膜)(图 4 A)。在禁食寒冷应激期间,心脏葡萄糖的消耗几乎完全受到抑制。在寒冷应激期间,心脏对乳酸的偏好也显著降低(图4B),这表明碳水化合物使用的减少可能至少部分受到丙酮酸利用水平的调节。相比之下,BAT 中葡萄糖的相对使用不受影响。由于 BAT 中的绝对 TCA 循环通量因寒冷应激而大幅增加,BAT 对葡萄糖的相对消耗量没有变化可能表明绝对葡萄糖消耗量大幅增加。因此,心脏对碳水化合物利用的抑制可能反映了寒冷诱导的适应性机制,以节省葡萄糖用于 BAT。
氨基酸代谢也以类似的方式重新连接,有助于将葡萄糖引导至 BAT。为了应对寒冷,肌肉对谷氨酰胺使用的偏好下降(图 4 C),谷氨酰胺转而用于糖异生。相反,寒冷应激显著增加肌肉对 BCAA 氧化的偏好,而 BCAA 不是 BAT 偏好的底物(图 4 D)。然而,血浆 BCAA 水平在寒冷应激期间大幅上升(图 1 F),可能暗示了质量作用效应,并表明骨骼肌中的 BCAA 氧化可能比其他器官对可用性变化更敏感。
他们的循环通量分析表明脂肪酸是禁食冷应激期间主要的产热燃料,与此一致的是,BAT 在应对寒冷时增加了对棕榈酸酯氧化的偏好(图 4 E)。结合寒冷条件下 BAT的 TCA 转化率较高,脂肪的贡献加倍反映了寒冷引起的 BAT 脂肪燃烧的显著上调。另一方面,3HB 在 BAT 中以最低程度地氧化,相反,肌肉组织大量使用它,其代谢通量在股四头肌寒冷应激期间显著增加(图4 F)。
结合这些数据并推断棕榈酸盐数据来预测总脂肪酸利用率,他们估计了在室温和4℃下每个组织中TCA循环的总营养贡献(图4G-H)。在不同的组织以及在不同的条件下,平均TCA循环碳占总量的75%,其余的可能来自细胞内的燃料储存,如糖原和甘油三酯。这些数据表明,在禁食期间,BAT 严重依赖脂肪酸氧化,细胞内脂质在基础条件下充当关键底物,并且急剧激活的 BAT 转变为主要消耗外源脂肪酸。

图4. 量化禁食小鼠急性寒冷应激期间器官特异性营养物偏好。
(A–G) 葡萄糖、乳酸、谷氨酰胺、支链氨基酸、棕榈酸盐、3-羟基丁酸盐和所有营养素的直接贡献,包括估算小鼠在室温下或在不进食的情况下急性应激于 4°C 6 小时不同器官中的总 FA 循环(苹果酸、琥珀酸和谷氨酸)。(H) 营养物质对 TCA 循环的贡献图。
04
评估急性寒冷应激对自由进食小鼠的代谢影响
接下来,他们对自由进食的小鼠进行了类似的通量分析,与维持室温的小鼠相比,应激于寒冷环境的小鼠摄入的食物量增加了一倍多(图 5 A)。与禁食的小鼠一样,它们表现出葡萄糖、乳酸、支链氨基酸、棕榈酸和 3HB 的循环通量增加,但谷氨酰胺则没有增加(图 5B)。尽管存在这种相似性,但禁食和进食条件下寒冷引起的变化的定量程度存在很大差异。与禁食的小鼠相比,喂食的小鼠碳水化合物循环通量的增加多于脂肪酸。进食小鼠的 3HB 循环通量低于禁食小鼠,但在寒冷应激期间其增加幅度相似,而进食小鼠的 BCAAs 通量在基线时较高,并且在寒冷应激期间增加幅度更大。寒冷应激期间 3HB 和 BCAAs 通量增加也与血浆水平增加有关,而其他营养素的丰度没有变化(图 5 C)。
他们再次量化了循环养分之间的相互转化通量并计算了每种养分的 EP 通量,与禁食小鼠不同,寒冷诱导的通量上升超过了 VCO 2,这表明进食小鼠中存在大量的寒冷诱导的合成代谢通量(图 5D)。
通过TCA 循环中间体中13C 的富集分数计算,寒冷应激对组织燃料偏好的影响在自由进食的小鼠中也有显著差异(图5E-5J)。喂食小鼠的所有组织中,葡萄糖对TCA循环的贡献更高,尤其是BAT、心脏和隔膜等高度氧化的器官(图5E)。重要的是,虽然禁食小鼠的BAT主要燃烧脂肪酸,但葡萄糖是喂养小鼠的主要三羧酸底物。此外,正如寒冷应激增加了BAT在空腹状态下对脂肪的偏好(尽管其对葡萄糖的偏好没有变化)一样,寒冷应激也增加了BAT在进食状态下对葡萄糖的偏好(尽管其对棕榈酸的偏好没有变化)。连同寒冷中总体 BAT TCA 循环的增加,这些数据反映了寒冷中脂肪和葡萄糖消耗的持续上调,特别是对于首选底物(禁食期间的脂肪和进食期间的葡萄糖)。
与禁食的小鼠不同,进食的小鼠在寒冷应激期间心脏中保留了碳水化合物的使用。相比之下,在寒冷应激期间,进食的动物表现出对谷氨酰胺的偏好下降,而对支链氨基酸的偏好大幅增加(图5G-H)。在喂养小鼠中,BCAAs对TCA循环的贡献在大多数其他器官(包括BAT)中也有所增加,但除四肢外,其他器官仍低于2%。在喂食的动物中,由于应激在寒冷环境中,隔膜和肝脏对棕榈酸盐氧化的偏好都会降低(图5I)。与禁食小鼠相比,喂食小鼠3HB对TCA循环的贡献始终较低,但在寒冷应激的几种组织中增加(图5J)。
将这些数据结合在一起,他们估计了在RT和4°C下喂养动物组织中TCA循环的总营养贡献(图5K)。与禁食动物一样,BAT 中 TCA 循环中营养成分的无法解释的部分随着寒冷而减少,表明对内部燃料储存的依赖减少。然而,在喂养的小鼠中,这种转变是由碳水化合物而不是脂肪酸的使用增加驱动的,这强调了 BAT 中的营养偏好是灵活的,并且在很大程度上受到喂养状态的影响。
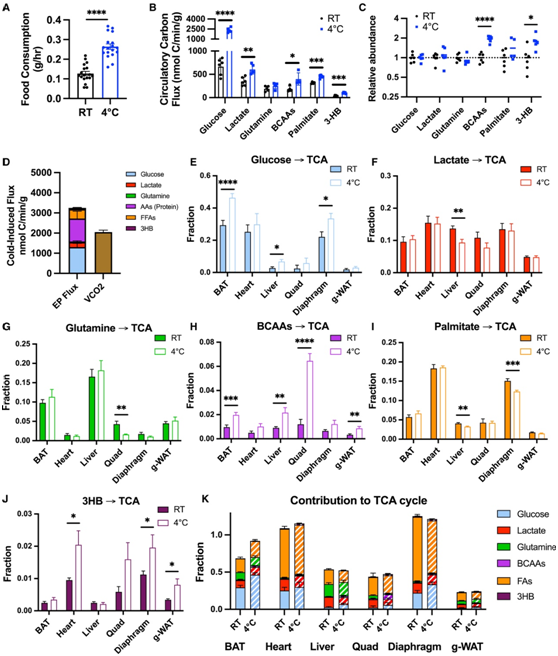
图5. 量化自由进食小鼠急性冷应激期间底物的利用。
(A) 在室温 (RT) 下饲养或急性应激于 4°C 6 小时的自由进食小鼠的食物消耗。(B-C) 循环碳通量和动脉血浆中关键代谢物的代谢物丰度。(D)自由采食期间寒冷诱导的最终代谢通量率和 VCO2的增加。(E–K)葡萄糖、乳酸、谷氨酰胺、支链氨基酸、棕榈酸盐、3-羟基丁酸盐和所有营养素的直接贡献。
05
在急性寒冷应激期间绘制葡萄糖命运图
众所周知,寒冷应激可诱导BAT的葡萄糖摄取,并且葡萄糖摄取可用于测定人类中的BAT质量和活性,寒冷应激也促进骨骼肌对葡萄糖的摄取。他们试图使用稳态输注系统来定量绘制冷应激期间的葡萄糖命运。
他们首先检测了血液循环中葡萄糖到乳酸的通量。在寒冷应激期间,在禁食和喂食的小鼠中,在寒冷中,乳酸的葡萄糖通量明显增加(图6A)。然而,在喂食动物中,只有22%的寒冷诱导葡萄糖通量用于乳酸生成,而在禁食动物中,这一比例为74%(图6A-B)。因此,寒冷诱导葡萄糖通量主要用于在禁食动物中产生乳酸。
葡萄糖衍生的丙酮酸盐不是转化为乳酸,而是运输到线粒体中,为TCA循环提供碳(图4A和5E),可以通过以下两种方式实现:(1)丙酮酸可以由丙酮酸脱氢酶(PDH)氧化脱羧为乙酰辅酶A (CoA),通过三羧酸循环进行完全氧化;(2)丙酮酸可以由丙酮酸羧化酶或其他酶(统称为PC)羧化,生成三羧酸循环中间产物草乙酸盐或苹果酸盐(图6C)。后一种无性腺反应有助于维持TCA循环池。通过同位素分析和13C标记的葡萄糖可以区分通过这两种通路的通量,因为PDH释放一个丙酮酸碳作为CO2来生成乙酰辅酶a的双碳单位,而通过PC生成的草乙酸盐/苹果酸保留了所有三个原始的丙酮酸碳(图6C)。令人惊讶的是,在禁食小鼠中,无论是在室温下饲养还是急性应激于4°C, BAT几乎完全通过PC通量将葡萄糖衍生的碳纳入TCA循环(图6D-E)。在自由进食的小鼠中,BAT 中的 PDH 通量受到影响,但丙酮酸羧化仍然是葡萄糖碳进入 TCA 循环的主要机制,无论是在室温下还是在寒冷应激期间。因此,BAT 中的丙酮酸羧化通量很大,占 TCA 循环总周转量的三分之一。相比之下,隔膜也大量利用葡萄糖进行 TCA 循环代谢,在禁食和进食状态下主要通过 PDH 通量来实现,而 PC 通量在寒冷应激期间完全消除。
在喂食的小鼠中,膳食碳水化合物提供循环葡萄糖,而寒冷引起的葡萄糖通量增加几乎完全可以归因于碳水化合物消耗的增加。然而,在禁食状态下,葡萄糖必须由其他来源提供,主要是糖原分解或糖异生。为了确定肝糖原分解对循环葡萄糖通量的贡献,他们检测了在室温或 4°C 禁食 6 小时的小鼠中肝糖原含量的下降情况。3 小时后,两种条件下的肝糖原含量均下降了 70% 以上。6 小时后,肝糖原进一步降低,应激于 4°C 的小鼠中更是如此。然而,后 3 小时时间间隔内的净糖原消耗仅占测得的全身葡萄糖通量的 1%(图 6F-G),表明在寒冷应激 3-6 小时的时候,肝糖原对循环葡萄糖供应的贡献最小。甘油和其他测量底物计算出的糖异生通量之和几乎占所有葡萄糖通量,无论是基础还是急性寒冷应激期间(图 6 H)。因此,糖异生是禁食小鼠中寒冷诱导的葡萄糖通量的主要来源(图 6 I)。
糖异生主要发生在肝脏中,少量发生在肾脏中。寒冷应激会增加关键糖异生酶 Pepck 和 FBP1 的肝脏蛋白水平,Pepck(而非 FBP1)的 mRNA 水平也增加,表明这种反应仅部分受到转录调节。为了检测 CIT 是否需要葡萄糖供应,他们建立了肝细胞特异性敲除 FBP1 的小鼠,与 PEPCK 不同,FBP1 是所有底物(包括甘油)糖异生所必需的(图6J)。通过注射AAV8-Tbg-Cre的Fbp1flox/flox小鼠建立肝脏特异性敲除(FBP1 LKO)小鼠和AAV8-Tbg-GFP注射的Fbp1flox/flox小鼠[野生型(WT)]作为对照。FBP1 LKO小鼠保持正常体重,肝糖原水平正常。将空腹FBP1 LKO小鼠应激于4℃条件时,其血糖水平相对于对照降低(图6K),证实肝脏糖异生是葡萄糖通量的关键来源。值得注意的是,空腹FBP1 LKO小鼠在急性应激于4℃时无法维持体温(图6L)。相比之下,向FBP1 LKO小鼠提供补充15%葡萄糖的饮用水或自由进食的食物,可在急性寒冷应激期间维持其血糖水平和体温(图6M-N)。总之,这些数据表明,肝脏糖异生作用维持了应激于寒冷的禁食小鼠中的葡萄糖供应,并且需要糖异生作用来支持这些小鼠的生热作用。
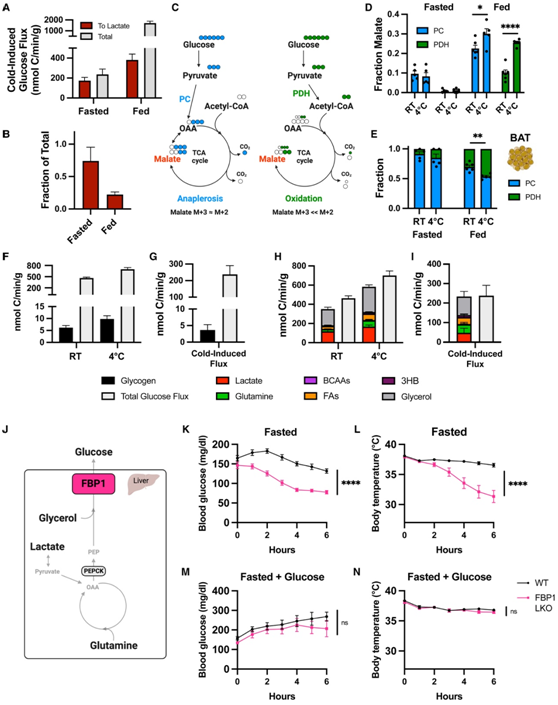
图6. 肝糖异生促进了BAT中的逆转,并且是耐寒性所必需的。
(A)寒冷诱导禁食或自由进食的小鼠中从葡萄糖到乳酸的通量和总葡萄糖通量的增加。(B) 寒冷引起的葡萄糖通量增加的比例。(C) 示意图显示通过丙酮酸羧化 (PC) 或丙酮酸脱氢酶 (PDH) 氧化从葡萄糖到苹果酸的碳示踪。(D)在室温 (RT) 或 4°C 下禁食和自由进食的小鼠的 BAT 中,通过 PC 或 PDH 通量从葡萄糖衍生的苹果酸分数。(E) BAT 中通过 PC 与 PDH 途径转化为苹果酸的总葡萄糖流量的比例。(F) 与禁食小鼠在 RT 或 4°C 下的总葡萄糖通量相比,3 至 6 小时糖原净减少导致的葡萄糖通量变化。(G) 寒冷引起的糖原葡萄糖通量和总葡萄糖通量的增加。(H) 各种营养素到葡萄糖的通量与室温或 4°C 下饲养的禁食小鼠的总葡萄糖通量相比。(I) 寒冷导致各种营养素到葡萄糖的通量和总葡萄糖通量增加。(J) 示意图显示肝脏中 FBP1 介导的糖异生。(K-L) FBP1肝脏敲除(FBP1 LKO)和对照组(WT)小鼠急性应激于4°C无食物环境下的血糖水平和体温。(M-N) 急性应激于 4°C 且不进食但饮用水中含有 15% 葡萄糖的 FBP1 LKO 和 WT 小鼠的血糖水平和体温。
+ + + + + + + + + + +
结 论
在本研究中,他们在未麻醉的小鼠中,利用代谢组学分析和最小扰动同位素示踪研究,对急性寒冷应激的代谢反应进行了全面和定量分析。在寒冷应激期间,棕色脂肪组织(BAT)主要通过禁食小鼠中的脂肪和喂食小鼠中的葡萄糖为三羧酸(TCA)循环提供燃料,这凸显了BAT的代谢灵活性。BAT很少使用支链氨基酸或酮,而在寒冷应激期间,肌肉会大量消耗这些氨基酸或酮。令人惊讶的是,同位素标记分析显示,BAT主要通过丙酮酸羧化来使用葡萄糖进行TCA再生。最后,本项研究发现寒冷诱导的肝脏糖异生对禁食期间的CIT至关重要,证明了其在葡萄糖代谢中的关键功能作用。
+ + + + +



