

 English
English文献解读|PNAS(11.205):具有自激活效应的NB-LRR导致小麦半矮化
✦ +
+
论文ID
原名:An autoactive NB-LRR gene causes Rht13 dwarfism in wheat
译名:具有自激活效应的NB-LRR导致小麦半矮化
期刊:PNAS
影响因子:11.205
发表时间:2022.11.23
DOI号:10.1073/pnas.2209875119
背 景
第二次绿色革命的代表性工程就是矮化表型小麦的育种。矮化小麦由于株高矮小,根吸收的碳源优先分配到种子用于繁殖,使得其产量相较于野生型大幅增加,同时还可以节约水资源。其矮化的原理主要是Rht18和Rht24基因座上发生的突变,可以使得编码赤霉素氧化酶的基因表达量上调,进而清除赤霉素的前体物质和赤霉素,使得植株出现矮化的表型。但这条路径存在弊端,例如会影响小麦的抗病性、降低小麦的出苗率等。所以找到一个不影响赤霉素途径的矮化基因是十分迫切的。
实验设计
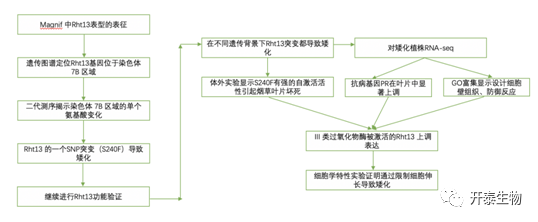
结 果
01

Rht13到染色体 7B 区域的精细遗传定位
首先,该团队从1980年的一篇诱导小麦矮化突变的文章中找到线索,1980年的团队使用甲基亚硝基脲作为突变诱导剂处理以Magnif(简称为MG)为遗传的小麦,获得了具有矮化表型的突变体Magnif M(MM),并得知其突变发生于Rht13 基因座上。作者将矮化突变体Magnif M与野生型同时种植比较。结果表明,突变体的株高出现了显著下降。同时,作者发现株高下降的主要原因是由于花序梗的变短。
已有研究表明,Rht13基因座位于小麦的7B号染色体的长臂上,其两侧有简单序列重复标记,gwm577和wmc276,同时作者又提供了多种简单序列重复标记,将Magnif M与正常株高表型的品种进行杂交后获得T2代,随后进行Genetic mapping(遗传作图),初步确定了Rht13的位置。

图1 NB-LRR基因Rht13的基因定位
02
二代测序揭示了染色体 7B 区域中表达基因之间的单个氨基酸变化
为了进一步确认Rht13的单核苷酸多态性SNP位点,作者在Magnnif 野生型x Magnnif M矮化突变体的杂交后代中选出了Rht13纯合的株系,通过流式分选分离出7B号染色体,然后对染色体进行测序。他们以CDC Stanley(具有显著单倍型差异的玉米品种)的染色体序列作为原始序列,对杂交后代的序列进行比对,发现在Rht13上存在着12个SNP和1个INDEL(插入缺失突变)。对了确定Rht13基因座上的矮化基因,作者对杂交后代的花序梗组织进行了转录组测序,发现相比于Magnnif野生型而言,Magnnif M矮化突变体存在着一个表达量更高的转录本,但是这个转录本并不会翻译成超过76个氨基酸的蛋白质,说明这是一个伪基因。随后作者将染色体测序和转录组数据结合,找到了一个可以翻译成蛋白质的转录本,其上有一个SNP。这个转录本编码了一个NB-LRR蛋白,SNP的发生使得240位的丝氨酸突变为苯丙氨酸(S240F),可能导致矮化。
03
不同遗传背景下Rht13矮小表型的表征
为了进一步确认这一个SNP的突变是否是导致小麦矮化的关键突变,作者在小麦突变体库中找到了基因序列于此突变完全一致的Cadenza0453株系。在Cadenza0453株系中,发生SNP的Rht-B13b相较于野生型Rht-B13a出现了明显的矮化,这种矮化的主要原因还是花序梗的缩短。同时,作者还将具有SNP突变形式的Rht-B13b转入了无Rht13b基因座的Fielder小麦中,发现转基因系均出现了明显的矮化。说明该SNP导致的氨基酸突变S240F就是致使小麦发生矮化的关键位点。
作者还使用了不同遗传背景的小麦作为底盘,分别是EGA Gregory、Espada和Magenta三种澳洲背景小麦。同时拿传统的矮化基因Rht-B1b 和 Rht-D1b与Rht-B13b进行矮化效果的比较。结果表明,对于茎的长度来说,在EGA Gregory(简称EG)和Espada(简称EP)背景中,Rht-B13b对于花序梗的缩短作用是强于Rht-B1b和Rht-D1b。在Magenta背景中,Rht-B13b对于茎和花序梗缩短都存在着更强的作用。结果说明,Rht-B13b在各种常用遗传背景的小麦中都可以发挥作用,且其矮化作用相比于传统的Rht-B1b 和 Rht-D1b更强。

图2 Rht-B13b中的S240F突变会导致株高降低
04
Rht-B13b具有自激活活性
在番茄中,NB-LRR也有同样的突变,即240位的丝氨酸突变为苯丙氨酸(S240F),而在番茄中该位点发生突变的结果就是导致NB-LRR发生了自激活,即获得了活性。所以作者提出了假设,猜测小麦的NB-LRR突变后也可有自激活活性。作者通过烟草瞬时转化实验,发现相较于未突变的Rht-B13a,Rht-B13b注射后烟草叶片发生显著细胞凋亡,类似于植物抗病响应的表型。但Rht-B13b类型的小麦并没有表型出和烟草一样的表型,所以作者猜测Rht13的自激活可能会增强防御反应,导致生长减少,而不会导致细胞凋亡。通过Rt-PCR检测发现,抗病基因PR3 和 PR4 在Rht-B13b在基部花序梗中都出现了显著的上调,说明NB-LRR的自激活促进了小麦的抗病反应。
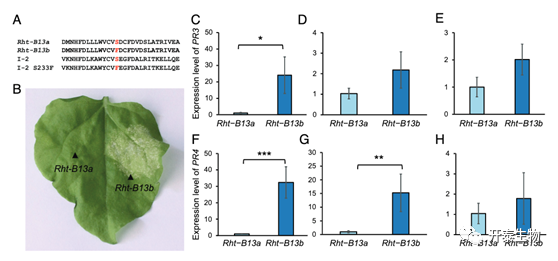
图3 Rht-B13b诱导小麦和烟草的防御基因响应
05
RNA-seq表明激活的Rht13 上调III 类过氧化物酶的表达
作者将Rht-B13a,Rht-B13b两种小麦的花序梗组织进行转录组测序分析,在GO富集中发现抗病基因出现了显著上调。随后作者提出假设:突变体系中防御反应的激活将导致活性氧的产生,活性氧可以促进细胞壁交联和细胞壁硬化,从而导致生长减少、植株矮化。所以作者关注了转录组数据中III类过氧化物酶的表达,发现有许多III类过氧化物酶基因出现上调,对花序梗中的过氧化氢进行检测,发现突变体中的过氧化氢相较于野生型显著减少,相应的,突变体的花序梗的强度和硬度都显著增加,细胞长度也显著缩短。作者进一步探究了花序梗强度和硬度的增强是否是由于木质素的积累引起,通过甲苯胺蓝染色实验,结果显示在花序梗的顶端和中部,木质素积累并没有显著的差异,而在基部花序梗中,突变体的木质素积累较野生型而言较少,说明花序梗硬度的变化主要还是由于细胞壁交联的发生。
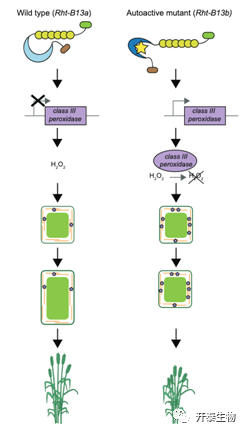
图4 Rht-B13b导致小麦半矮化的途径模型
+ + + + + + + + + + +
结 论
英国约翰英纳斯中心(John Innes Centre)的Philippa Borrill团队与合作者揭示了小麦新型矮杆基因Rht-B13的突变位点以及分子机理。等位基因的一个SNP突变S240F引起矮化,自激活的Rht-B13b等位基因导致抗性基因以及与细胞壁重塑相关的III类过氧化物酶基因的转录上调,从而导致小麦绕过赤霉素信号途径而实现矮化。矮化小麦增强了抗倒伏能力,极大的增加了小麦的产量,矮秆基因的鉴定为粮食增产,培养抗倒伏新品种提供了物质基础。
+ + + + +



