

 English
English文献解读|Nat Commun(14.7):非人类灵长类动物季节性转录组图谱揭示生理和疾病的季节性变化
✦ +
+
论文ID
原名:Non-human primate seasonal transcriptome atlas reveals seasonal changes in physiology and diseases
译名:非人类灵长类动物季节性转录组图谱揭示生理和疾病的季节性变化
期刊:Nature Communications
影响因子:14.7
发表时间:2025.04.28
DOI号:10.1038/s41467-025-57994-1
背 景
地球上的生命节律是由天体运动形成的。例如,为了应对环境中的季节性变化,动物在生理和行为的各个方面都会发生显著的变化,如繁殖、换羽、冬眠和迁徙。人类的生理过程也存在季节性差异,包括与激素分泌、新陈代谢、睡眠、免疫功能和繁殖有关的生理过程。此外,人类的心脑血管疾病、流感、肺炎、自身免疫性疾病、抑郁症、躁郁症和精神分裂症等疾病的发病率也有季节性变化,其中大多数疾病在冬季会变得更加严重,死亡率也更高。因此,深入了解驱动这些受季节性调节的生理和疾病易感性的机制至关重要。虽然这些季节性节律是由多种组织中协调的基因表达谱引起的,但其潜在的分子基础仍然未知。
实验设计
结 果
01

激素分泌的季节性变化
研究团队将恒河猴饲养在京都大学人类行为进化起源中心 (EHUB) 的开放式围栏中。在 2019 年 10 月到 2020 年 8 月期间,每 2 个月在 14:00 左右(日本标准时间)他们从雄性和雌性成年恒河猴(年龄从 4 岁到 11 岁不等)采集 80 个组织样本,具体来自 30 个大脑区域和 50 个外周组织。恒河猴的生殖表型表现出明显的季节性变化。在恒河猴繁殖季节(即 12 月和 2 月),雄性会发育出睾丸,而雌性则会表现出肛门生殖器皮肤发红的增加(图 1a)。采用毛细管电泳飞行时间质谱法(CE-MS)对血浆样本进行代谢组分析,检测到222种代谢物,并揭示了20种水溶性代谢物的季节性节律性,包括谷氨酸(Glu)、谷氨酰胺(Gln)、胱氨酸和犬尿氨酸(图 1b)。有趣的是,这些神经递质和代谢物已作为炎症和抑郁症的生物标志物。
在繁殖季节,雄性的睾酮水平和雌性的雌二醇水平都很高(图 1c)。然而,雄性睾酮的增加(即 10 月)早于雌性雌二醇的增加(即 12 月)。这一结果与之前的研究一致,该研究报告了雄性比雌性更早的生殖开始时间。夏季,雄性的睾酮水平和雌性的雌二醇水平也略有增加(图 1c)。值得注意的是,最近的一项人体研究报告了雄性睾酮和雌性雌二醇的双峰分泌曲线,在冬季和夏季都达到峰值。
他们从80个组织中的924个样本中提取了总RNA,并基于Lasy-Seq方案进转录组分析(RNA-seq)。基于转录组的主成分分析(PCA)清晰地区分了神经组织和外周组织(图 1d)。此外,根据外周组织的功能分类,形成了不同的亚聚类。脊椎动物的繁殖受下丘脑-垂体-性腺 (HPG) 轴控制。RNA-seq数据阐明了参与 HPG 轴的基因的季节性表达谱。在雄性恒河猴中,以下每个基因在 8 月至 10 月均出现峰值:下丘脑中的促性腺激素释放激素 (GNRH);垂体中的黄体生成素 (LH) 和促卵泡激素 (FSH) 的 β 亚基(例如LHB、FSHB);以及睾丸中参与睾酮生成的限速酶(类固醇生成急性调节蛋白 [STAR] 和羟基-δ-5-类固醇脱氢酶 3β 和类固醇 δ 异构酶 2 [HSD3B2])(图 1e)。这些基因的季节性表达谱在雌性中不太明显,可能是因为存在发情周期。与雄性相反,雌性中的LHB在12 月到 次年4 月较高,随后雌二醇合成的限速酶基因(STAR和芳香酶 [细胞色素 P450 家族成员 19 亚家族 A 成员 1:CYP19A1 ])延迟增加。在其他轴上也观察到每个过程的时间滞后和季节性表达谱的性别差异,例如参与代谢和体温调节的下丘脑-垂体-甲状腺轴,参与调控生长的下丘脑-垂体-肝脏轴和参与调控应激反应的下丘脑-垂体-肾上腺轴。下丘脑-垂体-外周腺轴的进程延迟发生在几分钟到几小时的量级。然而,他们发现转录水平上多个轴的延迟时间要长得多(例如几个月)。当比较性类固醇调控基因的季节性表达谱时,发现雄性比雌性表现出更早的表达阶段,这与睾酮和雌二醇分泌谱的差异相一致。
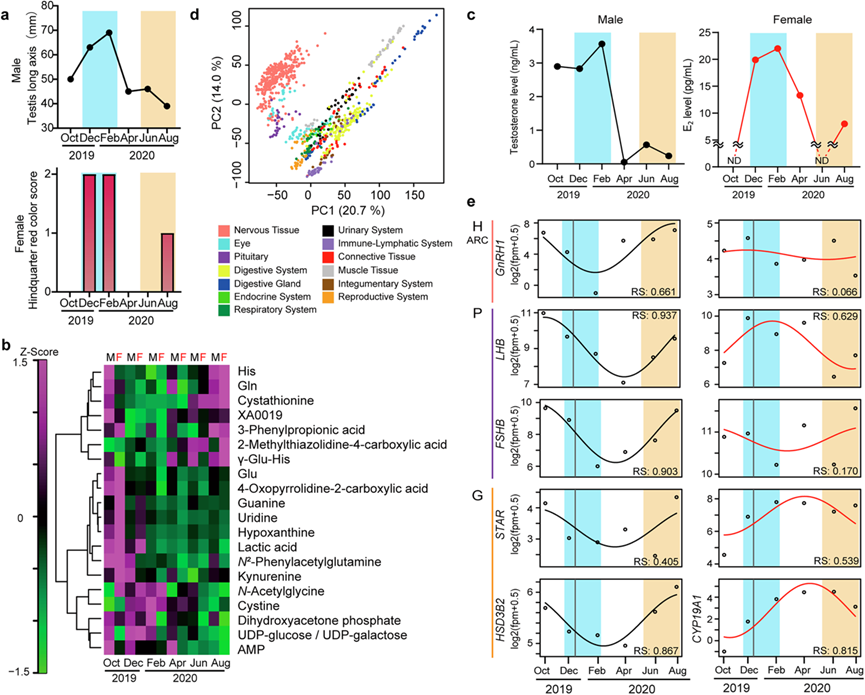
图1. 恒河猴的生理机能表现出明显的季节性变化。
(a) 雄性睾丸长度的季节性变化和雌性后肢皮肤的红色着色评分。(b) 热图显示通过 CE- MS测量的血浆代谢物的季节性变化。(c) 血浆睾酮和雌二醇的季节性分泌概况。(d) 对 80 个神经和外周组织中 924 个样本的转录组进行主成分分析。(e) 通过 RNA-seq 分析确定的下丘脑-垂体-性腺轴相关基因的季节性表达概况。
02
季节性波动基因(SOG)的鉴定
接下来他们确定了每种组织中的 SOG。为了减少因样本量小而导致的假阳性,在分析的多个步骤中应用了严格的参数。根据组织不同,在雄性中共检测到 4501-13544 个基因,在雌性中共检测到 4418-12954 个基因(图 2a)。对雄性和雌性之间表达基因的比较显示了大量重叠基因。为了研究单个基因的季节性表达,使用 R 中的非线性最小二乘 (nls) 函数拟合余弦曲线来分析趋势,并使用判定系数 (R 平方:RS) 评估拟合优度。如果 RS > 0.7 且季节性波动的幅度大于 1(具体来说,大 2 倍),则提取 SOG。该分析在雄性中鉴定出 273-2344 个 SOG,在雌性中鉴定出 342-1943 个 SOG,具体取决于组织(图 2b)。
在所检查的 80 种组织中,胰腺 (PAN) 在雄性和雌性中表达的基因数量和 SOG 数量都最低。腹肌 (MUA)、腓肠肌 (MUG) 和心脏 (HEA) 等肌肉组织也具有较低表达基因和 SOG 数量(图 2a-b)。这些组织中表达基因的数量较低与狒狒 RNA 测序结果一致。相反,两种性别的胃 (STM) 中以及生殖器官,特别是雄性的前列腺 (PRO) 和雌性的子宫 (UTE) 中都有大量 SOG(图 2b),表达基因的数量和 SOG 数量之间仅观察到弱相关性。
雄性和雌性每种组织中 SOG 的总数和峰值阶段有所不同(图 2b-c)。大多数组织类型在春季和/或秋季都有双峰或单峰峰值阶段,并且峰值阶段的分布存在性别差异。此外,对雄性和雌性 SOG 的比较分析,显示重叠的 SOG 数量有限[2.62%(食道:OES)- 24.8%(背部皮肤:SKNB)(平均:9.53%)在雄性中,3.53%(弓状核:ARC)- 31.3%(胃:STM)(平均:9.02%)在雌性中,具体取决于组织](图 3)。这些结果表明不同组织的季节性生理变化存在明显的性别差异。
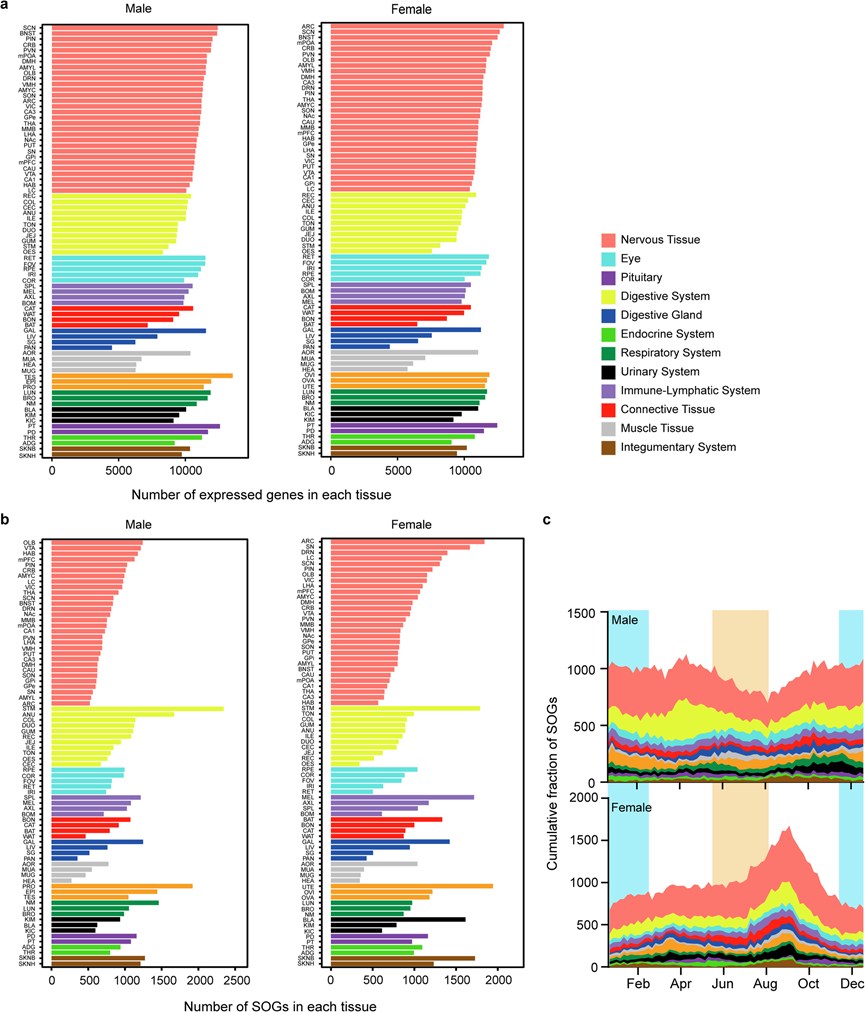
图2. 雄性和雌性季节性波动基因的识别。
(a) 每种组织中表达基因的数量。(b) 每种组织中 SOG 的数量。(c) 一年内不同组织类型中基因表达峰值阶段的累积分布。
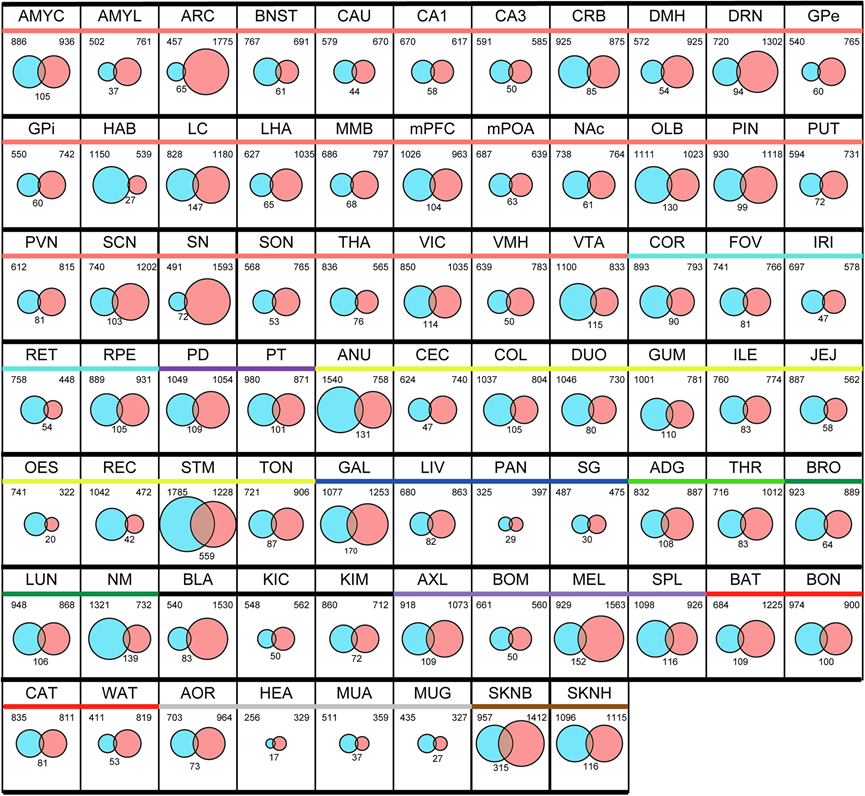
图3. 76 种组织中雄性和雌性恒河猴的季节性波动基因比较。
03
生理变化的分子分析
为了鉴定在两性之间保守的受季节调节的生理功能,他们使用 Metascape技术对两性之间常见重叠的 SOG 进行了功能富集分析(图4a-b)。在棕色脂肪组织 (BAT) 中, NDUFS2,IDH1和COX1观察到季节性波动,与“响应氧化应激”和“前体代谢物和能量的产生”相关的通路发生富集(图4a-b)。在包括人类在内的许多哺乳动物中,毛囊循环与每年的季节相协调。在背部皮肤 (SKNB) 中的FOXN1,COL1A1和KRT5中观察到季节性节律,突出了“表皮发育”相关通路的富集(图4a-b)。据报道,免疫功能具有季节性变化。他们发现肠系膜淋巴结 (MEL) 中LILRB4、IL18和IGF1的表达,以及脾脏 (SPL) 中 LILRB4、CXCL10和CCR2 的表达均存在季节性变化(图4b)。MEL 中“髓系细胞分化”和“活化 T 细胞增殖调节”相关通路富集,SPL 中“T 细胞细胞因子产生”相关通路富集(图 4a)。女性对冬季抑郁症的易感性较高,并且已知患者在冬季会表现出食欲增加和对碳水化合物的渴望。虽然恒河猴全年都以相同的饮食为食,但参与“碳水化合物代谢过程”的基因,如GLO1、LDHB和UGDH ,在雌性十二指肠 (DUO) 中的含量在冬季和春季达到峰值表达(图 4a-b)。季节性节律是由内源性周年生物钟驱动的,而周年生物钟则受到日照长度和温度的季节性变化的影响。据报道,在恒定的光周期、温度和食物供应下饲养的雄性恒河猴会持续保持年度雄激素节律。垂体的结节部 (PT) 是季节性节律的关键调节器,并在周年节律中起主要作用。对 PT 中的 SOG 进行功能富集分析,揭示了参与组织重塑的基因的过度表达,例如NOG、CCN3和CAV1(图 4b),季节性生物钟可能源于组织重塑的产生。
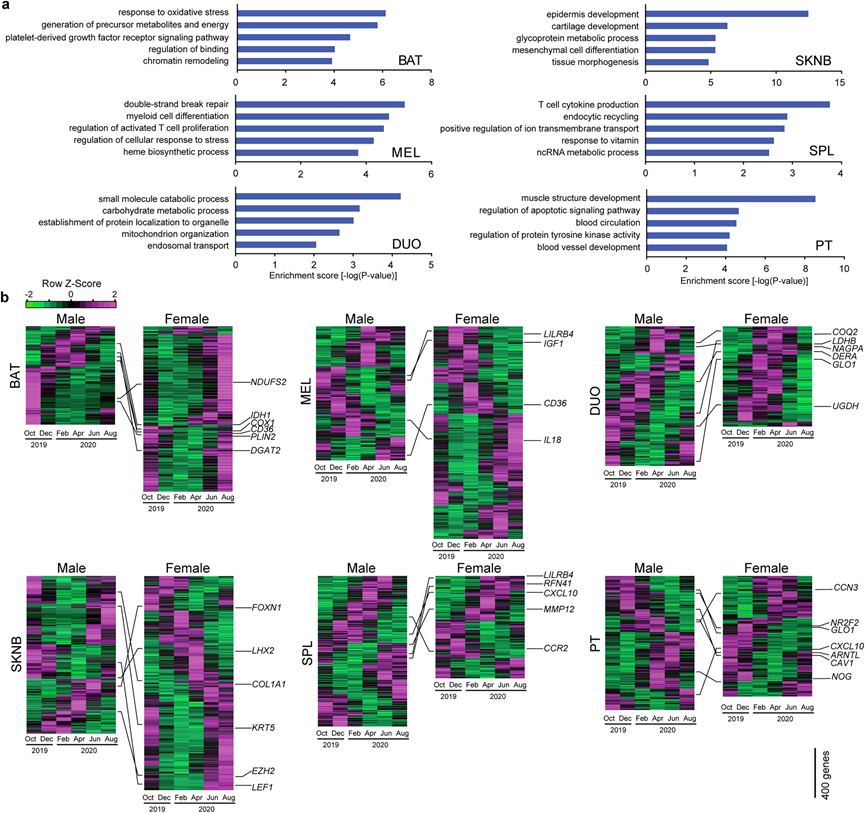
图4. 季节性转录协调季节性生理功能。
(a) 两性常见SOG的GO富集分析。(b) 热图显示了SOG的表达谱。
04
GA结合蛋白(GABP)对SOG的调节
他们进一步分析了在雄性和雌性多个组织中最普遍检测为SOG的基因(图5a)。在80个组织共计19003个SOG中,没有任何基因能在全部80个组织中均归为SOG(图5a)。聚焦前30个最常见的SOG时,他们发现与"需氧电子传递链"相关基因(如NDUFA1、SDHD、LDHB、COX5B和COQ9)以及核糖体蛋白基因(如RPS27L和RPL35)存在显著富集现象(图 5a-b)。
为了进一步了解调节这些季节性节律的分子机制,他们使用公开数据库中的 ENCODE 转录因子靶点数据集对雄性和雌性进行了转录调控网络分析(图 5c)。该分析确定了 E26 转化特异性 (ETS) 家族转录因子 GABP 及其相互作用的转录因子和共激活因子(例如 SP1、YY1、E2F1 和 CREB)(图 5c)。GABP 是由两种不同蛋白质 GABPα 和 GABPβ 组成的异四聚体。GABPα 包含 ETS DNA 结合结构域,而 GABPβ1 和 GABPβ2 包含转录激活结构域。由于在各种恒河猴组织中GABPB2的表达水平比 GABPB1 低约 2 个数量级,因此本研究重点研究了GABPB1。为了检测 GABP 是否调节 SOG 的表达,他们利用恒河猴成纤维细胞研究了小干扰 RNA (siRNA) 敲低GABPA和GABPB1的影响。首先使用定量实时 PCR (qPCR) 来评估敲低GABPA和GABPB1对其表达的影响。GABP 调节自身表达,他们证实了敲低每个基因对其自身表达的影响以及其自身调节效应(即上调其伴侣基因)(图 5d),证明了 siRNA 敲低的有效性。他们还使用 qPCR 证明 GABP 控制最常见的 SOG,例如NDUFA1、RPL35、SDHD1和LDHB(图 5e)。为了进一步探索GABP靶向的SOG,他们比较了转染了GABPA和GABPB1 siRNA的恒河猴成纤维细胞与阴性对照siRNA的RNA-seq分析结果。虽然GABPA和GABPB1的siRNA在统计学上显著敲低,但GABPA和GABPB1的敲低效率分别为50%和69%(图 5d)。因此,他们使用调整后的P值(Padj)阈值<0.05和差异倍数>1.5来检测差异表达基因(DEG)(图 5f)。该分析鉴定了 2058 个 GABP 调控基因(在 GABP 敲低细胞中 1067 个上调基因和 991 个下调基因)(图 5f)。由于成纤维细胞遍布所有组织,他们检查了 GABP 调节基因与所有 19003 个 SOG 之间的重叠。该分析显示,成纤维细胞中 70.4%的 GABP 调节基因(1450 个基因)是至少一种组织中的 SOG(图 5g)。对这 1450 个重叠基因进行GO分析,突出显示了与细胞周期有关的通路(图 5h),这与季节性适应性生理涉及身体成分、器官大小和组织重塑的变化的结果一致。
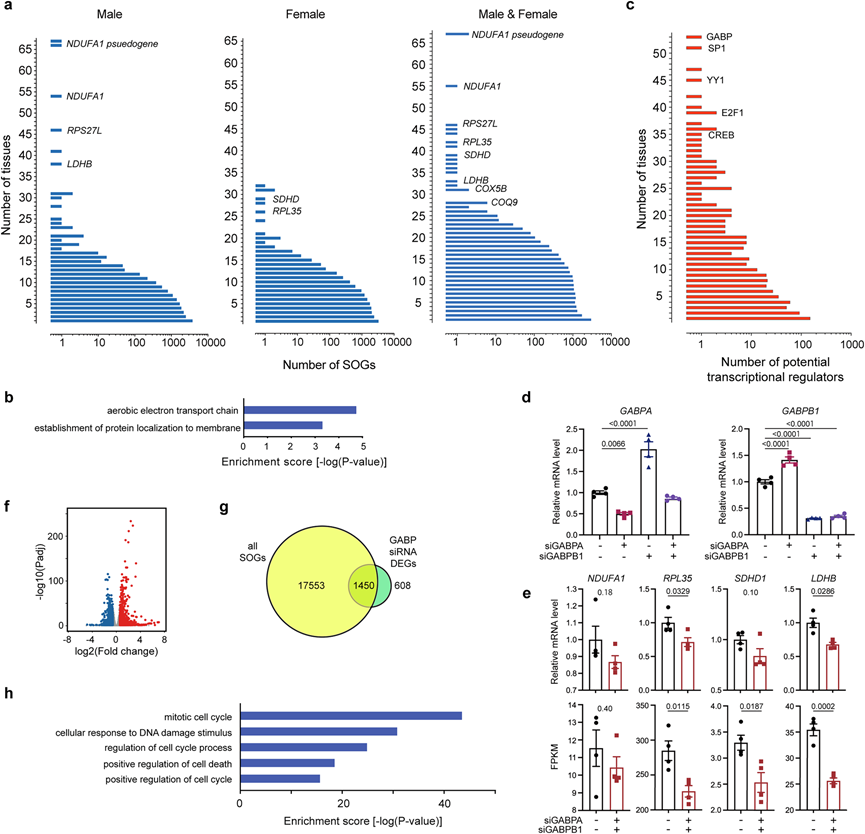
图5. GABP 对 SOG 的调节。
(a) 根据在雄性、雌性和两性中表现出季节性波动的组织数量对 SOG 的分布进行排序。(b) 对前 30 种最常见 SOG 进行GO富集分析。(c) 根据转录调控网络分析确定的组织数量对调节 SOG 的潜在转录因子的分布进行排序。(d) siRNA敲低GABPA和/或GABPB1对猕猴成纤维细胞中GABPA和GABPB1表达的影响。(e) 通过 qPCR(上)和 RNA-seq(下)检测siRNA 敲低GABPA和GABPB1对猕猴成纤维细胞中最常见 SOG 的影响。(f) 比较GABPA和GABPB1 siRNA 处理与阴性对照 siRNA 处理的恒河猴成纤维细胞中的 2058个DEG。(g) 比较所有 19003 个 SOG 和 2058 个 GABP 调控基因。(h)GO分析。
05
小鼠的组织重塑随着光周期和温度的变化而发生
由于灵长类动物的功能分析成本高且耗时,跨物种分析广泛应用于灵长类动物研究。大量研究表明,小鼠可以模拟季节性生理和行为的分子机制。因此,他们接下来重点研究了小鼠进行功能分析。在哺乳动物中,松果体褪黑激素在控制许多季节性功能和行为方面发挥着至关重要的作用。由于大多数常用的实验室小鼠品系在基因上缺乏褪黑激素,将他们它们分类为非季节性动物。然而,在之前的研究中,他们发现褪黑激素过高的 CBA/N 小鼠可以在下丘脑-垂体水平转换光周期信息。因此,人民研究了暴露于类似冬季的短日照和寒冷条件(SC:8 小时光照 16 小时黑暗,10°C)或类似夏季的长日照和温暖条件(LW:16 小时光照 8 小时黑暗,30°C)一个月对褪黑素过高的 CBA/N 和褪黑素缺乏的 C57BL/6 N 品系器官重量的影响。令人惊讶的是,他们发现 HEA、PAN 和肾脏(KID)的器官重量发生了显著变化,CBA/N 和 C57BL/6 N 小鼠在 SC 条件下的重量都较高(图 6a)。这些结果表明小鼠的器官重量可能存在季节性变化,而褪黑素不是这种调节所必需的。
然后他们使用免疫组织化学方法检测了 SC 和 LW 条件对 C57BL/6 N 小鼠组织重塑的影响,并对细胞分裂的标志物 Ki67 进行了检测。对暴露于 SC 或 LW 条件 2 周或 1 个月的小鼠进行初步分析,发现 Ki67 免疫阳性细胞很少。因此,他们使用暴露于 SC 或 LW 条件 1 周的小鼠的样本进行后续分析。SC 下 HEA 和 PAN 中的 Ki67 阳性细胞数量高于 LW 条件(图 6b-c)。虽然两种条件下 KID 中的 Ki67 阳性细胞数量没有差异,但 SC 下的肾小球尺寸明显大于 LW 条件下的肾小球尺寸(图 6b-c)。当他们对 HEA 进行苏木精和伊红 (H&E) 染色时,发现 SC 下的心肌细胞直径大于 LW 条件下的心肌细胞直径(图 6b-d)。定位于腺泡细胞膜的 β-catenin 免疫组织化学分析显示 SC 下的腺泡细胞尺寸大于 LW 条件下的腺泡细胞尺寸(图 6b-e)。这些结果表明,小鼠在SC和LW条件下发生了组织重塑。鉴于SC和LW条件下食物和水摄入量的显著差异(图 6f),这些结果可能反映了模拟夏季和冬季环境之间的代谢差异。虽然SC和LW条件下的光周期和温度变化是季节性变化的组成部分,但仍需要进一步研究来确认这些变化是否在自然环境条件下具有季节性。
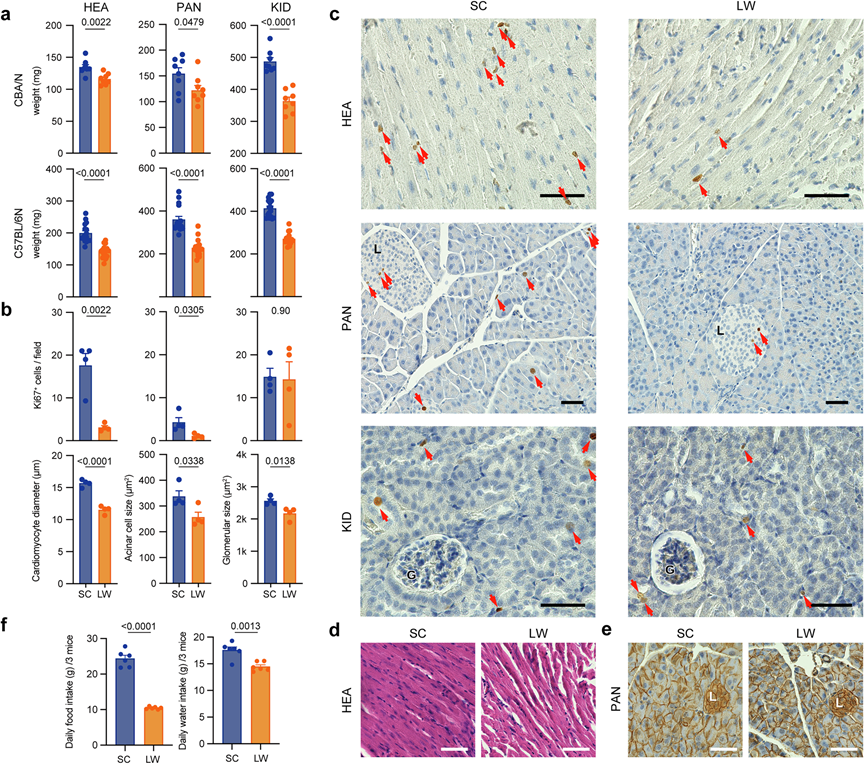
图6. 小鼠心脏、胰腺和肾脏的季节性组织重塑。
(a) 暴露于短日照低温 (SC) 或长日照高温 (LW) 条件1个月对 CBA/N 和 C57BL/6 N 小鼠器官重量的影响。(b) (上)暴露于 SC 或 LW 条件 1 周对 C57BL/N 小鼠心脏 (HEA)、胰腺 (PAN) 和肾脏 (KID) 中 Ki67 阳性细胞数量的影响。(下)暴露于 SC 或 LW 条件 1 周对心肌细胞直径、胰腺腺泡细胞大小和肾小球大小的影响。(c) 在 SC 和 LW 条件下心脏 (HEA)、胰腺 (PAN) 和肾脏 (KID) 的 Ki67 免疫组织化学代表性图像。(d) 在 SC 和 LW 条件下心肌细胞的H &E 染色代表性图像。(e) 位于腺泡细胞膜的 β-catenin 免疫组织化学代表性图像。(f) 在SC 和 LW 条件下饲养的雄性 C57BL/6 N 小鼠的每日食物和水摄入量。
06
GABP在体内组织重塑中的作用
为了测试 GABP 在光周期和温度诱导的组织重塑中的体内作用,他们利用 CRISPR/Cas9 基因组编辑技术生成了Gabpb1外显子 9 缺失突变体(图 7a)。他们将 crRNA、tracrRNA 和 Cas9 蛋白共注射到从 C57BL/6 N 小鼠中获得的 223 个受精卵中,然后将注射的胚胎移植到假孕 ICR 受体的输卵管中,成功获得了 16 个 G0后代。与之前的研究一致,G0嵌合体具有弱的后肢表型。相反,Gabpb1外显子 9 缺失纯合突变体断奶后无法存活。由于纯合Gabpb1外显子 9 缺失突变体在断奶后无法存活,他们分析了杂合突变体 (+/Mut) 和野生型 (+/+) 同窝仔的表型。当雄性野生型和杂合突变小鼠暴露于 SC 或 LW 条件 1 周时,野生型小鼠在 SC 下的Gabpb1表达往往高于 LW 条件(图 7b),而杂合突变小鼠的 Gabpb1 表达低于野生型小鼠(图 7b)。与野生型同窝仔鼠相比,杂合突变小鼠在 SC 和 LW 条件下器官重量变化的幅度较小(图 7b)。此外,杂合突变小鼠的HEA和PAN中Ki67阳性细胞数量的差异小于野生型小鼠(图 7b-f)。此外,杂合突变小鼠的心肌细胞直径、腺泡细胞大小和肾小球大小的变化幅度小于野生型小鼠(图 7c-f)。这些结果表明Gapb1参与调节光周期和温度诱导的组织重塑变化。
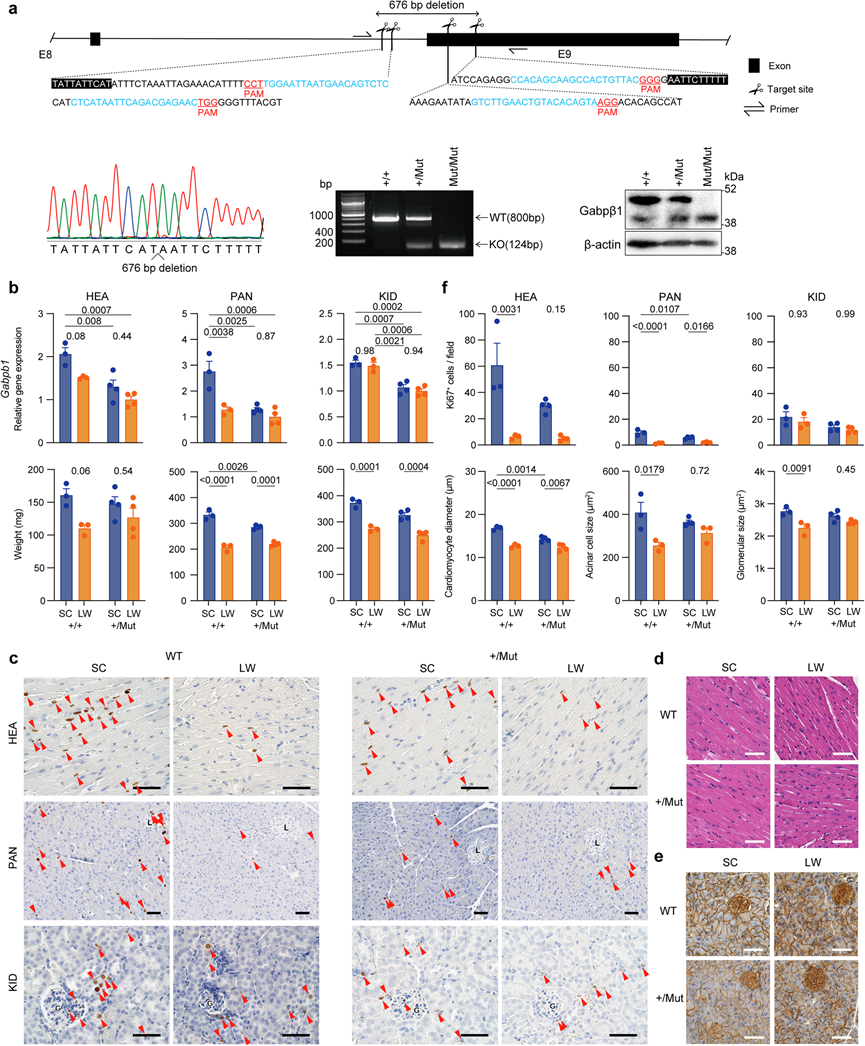
图7. GABP 在小鼠体内的作用。
(a) CRISPR/Cas9 靶向小鼠Gabpb1外显子 9。(b) 为期一周的 SC 或 LW 暴露条件对Gabpb1外显子 9 缺失突变杂合小鼠及其野生型同窝仔的Gabpb1表达和器官重量的影响。(c) 心脏 (HEA)、胰腺 (PAN) 和肾脏 (KID) 中 Ki67 的免疫组织化学。(d) 心肌细胞的H &E 染色。(e) β-catenin的免疫组织化学,其定位于腺泡细胞膜。(f) Ki67阳性细胞数量、心肌细胞直径、胰腺腺泡细胞大小和肾小球大小分析。
07
疾病相关基因的变化
为了了解季节性疾病的分子基础,他们使用了 Metascape 数据中名为 DisGeNET 的人类基因-疾病关联数据源,在雄性和雌性恒河猴常见的 SOG 中寻找疾病风险因子基因,发现 SOG 与多种疾病密切相关(图 8a),并在肺 (LUN)、主动脉 (AOR)、支气管 (BRO) 和结肠 (COL) 中鉴定了它们的风险因子基因(图 8b)。例如,在 LUN 中观察到TLR3和CCR2表达的季节性节律,以及“肺炎和流感”基因的富集(图 8a-b)。在AOR中,CD36和CXCL16表现出季节性转录节律,“血管炎症”和“急性冠状动脉综合征”相关基因富集;而在BRO中,CD40和C3的表达表现出季节性波动,“鼻窦炎”相关基因富集(图 8a-b)。此外,在COL中,ATF4和IRF8表现出季节性节律,“细菌感染”相关基因富集(图 8a-b)。
严重急性呼吸系统综合症冠状病毒 2 (SARS-CoV-2) 及其相关症状的出现,即 2019 冠状病毒病 (COVID-19),已在世界范围内造成了毁灭性的大流行。在流行阶段,SARS-CoV-2 感染浪潮的时间和强度受多种因素的影响,例如疫苗接种、实施和停止非药物干预措施(例如社会封锁、旅行限制、使用口罩)以及病毒进化导致传染性增强。然而,在流行期间,如果没有新的干预措施,SARS-CoV-2 感染预计会遵循与其他季节性冠状病毒(如 OC43、NL63 和 229E 45、46)类似的季节性模式。目前的证据表明,COVID-19 存在性别二态性,男性的严重程度和死亡率高于女性。 SARS-CoV-2 利用血管紧张素转换酶 2 (ACE2) 和丝氨酸蛋白酶 TMPRSS2 进入细胞。值得注意的是,本研究在两种性别的恒河猴的 BRO 中都观察到了 TMPRSS2的动态季节性波动,雄性的节律性比雌性更明显(图8c)。临床研究表明,患有糖尿病、肥胖症、心血管疾病和高血压的患者的 COVID-19 预后更为严重,而这些合并症与外周组织中ACE2和TMPRSS2的表达有关。事实上,他们发现这些受体在各种外周组织中都有表达,尤其是在消化系统和呼吸系统中表达较高。精神分裂症、抑郁症和双相情感障碍等精神疾病也受季节调控。他们发现与精神分裂症相关的基因表达谱在多个脑区存在季节性波动(图 8d)。例如,LRRTM1和DLG2在内侧前额叶皮质中表现出季节性波动,而PPP1R1B在杏仁核中央核和腹侧被盖区表现出季节性节律。
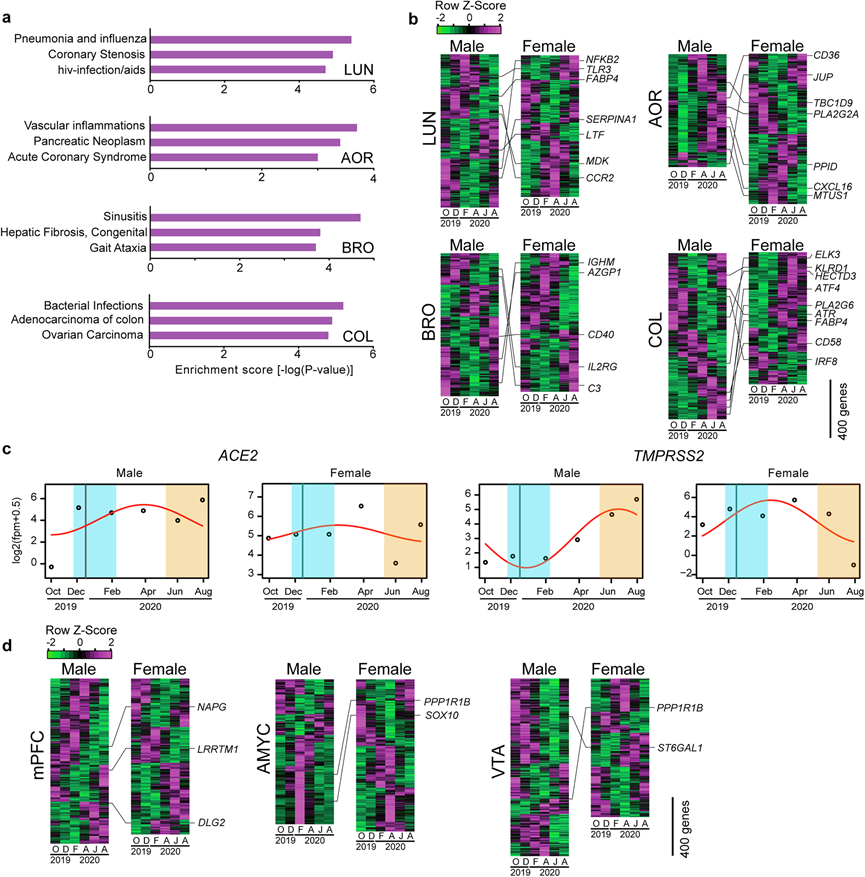
图8. 疾病相关基因表达的季节性变化。
(a) 肺 (LUN)、主动脉 (AOR)、支气管 (BRO) 和结肠 (COL) 中 SOG的 DisGeNET 富集分析结果。(b) 热图显示 SOG的表达谱。(c) 男性和女性 BRO 中ACE2和TMPRSS2的季节性表达谱。(d) 热图显示 SOG的表达谱。
08
药物疗效可能与季节变化有关
越来越多的证据表明各种药物的疗效与一天中的时间有关,而时间疗法旨在通过考虑昼夜节律来优化医疗效果。然而,药物的季节依赖性疗效仍然未知。为了确定哪些药物的疗效可能受季节影响,他们将本研究中的所有 19003 种 SOG 与药物基因相互作用数据库(DGIdb v5.0)进行了比较。根据 USP 医疗保险模型指南,按目标 SOG 数量排名的前 50 种药物中,约 75% 是抗肿瘤药物(图 9a-b)。正如他们所证明的季节性调节组织重塑(图 6和7),抗肿瘤药物的富集似乎是合理的。此外,类固醇激素表现出季节性变化(图 1),因此鉴定激素类药物地塞米松和孕酮似乎是合理的(图 9a-b)。心血管疾病、代谢、免疫功能和精神疾病都受季节调节。值得注意的是,在前 50 种药物中检测到了心血管药物阿托伐他汀、血糖调节剂盐酸二甲双胍、免疫药物雷帕霉素和环孢菌素以及抗精神病药物氟哌啶醇和奥氮平(图 9a-b)。在前 50 种药物中,他们研究了 LW 或 SC 条件对通过口服管饲给予 C57BL/6 N 小鼠的乙醇(EtOH)的影响。与水处理的对照小鼠相比,在 SC 和 LW 条件下,注射 EtOH 1 小时后,自发运动活动(以旷场测试中的总行进距离衡量)均显著减少。SC 小鼠在注射后 3 小时恢复,而 LW 小鼠的活动继续减少(图 9c)。为了验证EtOH功效的季节性变化,他们还使用加速转棒检查了 EtOH 对运动协调性的影响。正如预期的那样,在 SC 和 LW 条件下,治疗后 4 小时,EtOH 治疗小鼠的转棒表现均差于对照小鼠。虽然 SC 小鼠在给药后 5 小时的转棒耐力时间有所提高,但此时 LW 小鼠的转棒耐力时间仍明显短于对照小鼠(图 9d)。血浆 EtOH 检测结果进一步证实了与 SC 条件相比,治疗后 3 和 5 小时 LW 条件下的EtOH代谢降低(图 9e)。这些结果表明,EtOH功效会随着季节相关变量的变化而变化。值得注意的是,每月因EtOH相关原因入院的人数在夏季达到峰值。
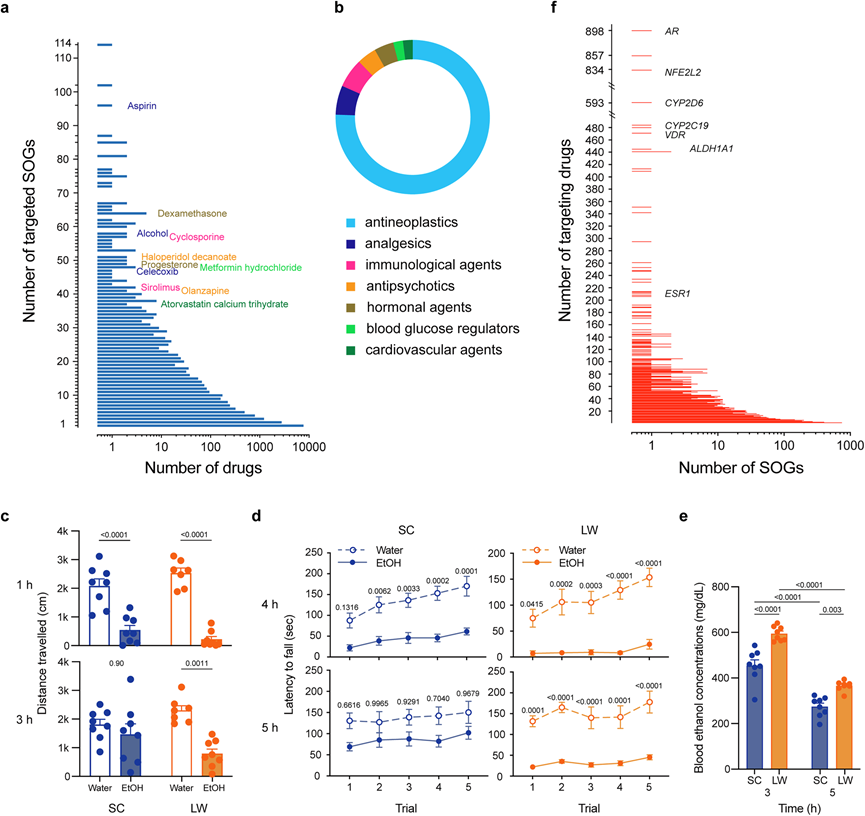
图9. 药物疗效的季节性变化。
(a) 根据目标 SOG 数量对药物的分布进行排序。(b) 按目标 SOG 数量排序的前 50 种药物的治疗类别。(c) 自发运动活动的影响。(d) EtOH对在 SC 或 LW 条件下小鼠的转棒表现的影响。(e) SC和LW条件对EtOH代谢的影响。(f) 根据靶向药物数量排序的SOG分布。
+ + + + + + + + + + +
结 论
本研究分析了在半自然户外环境中饲养的雄性和雌性恒河猴一年多的时间里采集的80个组织的季节性转录组,发现血浆代谢物和激素存在季节性变化。转录组分析确定了所有研究组织中SOG的性别差异。转录调控网络分析、siRNA敲低和突变小鼠分析揭示了GABP对SOG的调控。本研究还证明了疾病风险因素基因和药物相互作用基因表达的季节性波动。 NHPSTA 为季节性调节的生理学提供分子资源,并为季节性调节疾病的治疗干预提供目标。
+ + + + +



