

 English
English文献解读|Cell(64.5):宿主特征影响野生小型哺乳动物的病毒组组成和病毒传播
✦ +
+
论文ID
原名:Host traits shape virome composition and virus transmission in wild small mammals
译名:宿主特征影响野生小型哺乳动物的病毒组组成和病毒传播
期刊:Cell
影响因子:64.5
发表时间:2023.10.12
DOI号:10.1016/j.cell.2023.08.029
背 景
大多数人类病原体都起源于动物,并通过跨物种传播事件出现。蝙蝠、啮齿动物和鼩鼱是人类传染病最重要的动物来源。SARS-CoV-2的出现和COVID-19大流行的原因,突显了哺乳动物野生动物物种中人畜共患病毒的多样性,以及迫切需要了解它们在将这些病毒传播给人类方面的潜在作用。
实验设计
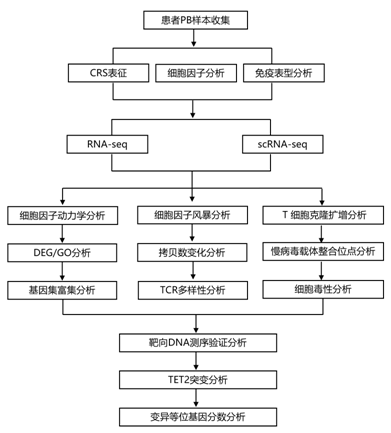
结 果
01

野生小型哺乳动物的丰富度和生态特征
为了更好地了解蝙蝠、啮齿动物和鼩鼱体内和之间病毒的多样性、进化和传播,研究者团队在中国湖北省和浙江省选择了四个地点对这些哺乳动物进行实地研究(图1A)。采样的小型哺乳动物有 13 至 18 个物种,表现出较高的物种多样性和每个地点不同的物种组成(图1B-D)。每个物种的丰度差异很大。以蝙蝠和啮齿动物为例,四个栖息地中每个栖息地都有两到五个物种,样本量超过 30 只。与蝙蝠和啮齿类动物相比,鼩鼱在这四个地点的物种丰富度都相对较低。所有三个目哺乳动物的数量和组成在四个地点之间差异很大(图 1D),反映了当地的生物地理因素。最后,15 种哺乳动物物种出现在两个或多个地点,而其余物种仅在一个地点发现(图 1 F)。为了验证采样的哺乳动物是否反映了每个研究地点的真实丰富度,他们进行了稀疏分析,在所有四个地点都观察到了递减趋势(图 1 E)。
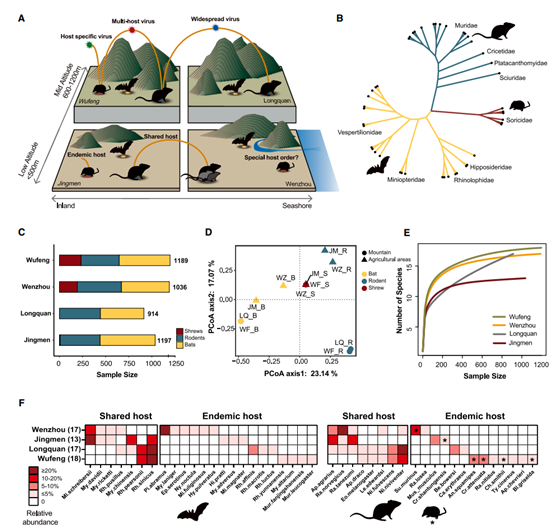
图1. 样本地点、景观和野生小型哺乳动物组成。
(A)本研究的地理位置及景观。(B)三个哺乳动物目的系统发育,标记了本研究中包含的9个哺乳动物科。(C)各地点哺乳动物的目组成。(D)主坐标分析显示山地和农业区之间哺乳动物组成的差异。(E)各地点哺乳动物样本量的稀疏曲线。(F)各采样点的哺乳动物种类组成。
02
野生小型哺乳动物的病毒组
通过宏转录组分析(Metatranscriptome),他们鉴定和表征了来自 > 42 个科的 669 种脊椎动物和无脊椎动物相关病毒,其中包括 > 40 个 RNA 病毒科和两个 DNA 病毒家族(图 2 A),在采样的哺乳动物中存在多种且丰富的病毒并且高度流行(图2A)。
除了仅捕获一只马蹄蝠 (Rhinolophus luctus) 之外,在所有其余哺乳动物物种中均发现了 1-150 种病毒,其中数量最多的是史密斯鼩鼱 (Chodsigoa smithii)。然而,三个哺乳动物目中病毒组的组成在病毒家族水平上差异很大(图 2B-C)。
检测到最多的病毒来自副粘病毒科、小核糖核酸病毒科、星状病毒科、黄病毒科、弹状病毒科、Sedoreoviridae的病毒以及来自小核病毒目的未分类病毒(来自三个目的超过 20 种哺乳动物物种),占本项研究鉴定的病毒总数的 48.0%(图2A-C)。
最后,不同采样地点的病毒组组成也不同,在主坐标分析(PCoA)图中,来自五峰和龙泉(山林)的哺乳动物与来自温州和荆门(有林地和山区的农业地区)的哺乳动物不同(图2B)。特别是五峰地区哺乳动物携带病毒较多(图2C)。

图2. 野生小型哺乳动物中与脊椎动物和无脊椎动物相关的病毒组。
(A)野生小型哺乳动物中脊椎动物和无脊椎动物相关病毒组的物种丰富度和丰度。(B)主坐标分析显示了哺乳动物目之间以及山区和农业区之间病毒组组成的差异。(C)确定每个宿主物种和每个采样位置的病毒组组成。
03
野生小型哺乳动物病毒的器官分布和器官内丰度
接下来他们试图确定病毒在内脏器官内的分布和丰度,来自不同科的病毒在这些野生哺乳动物的器官分布和器官内丰度方面表现出相当大的差异(图3A)。几乎所有来自星状病毒科、杯状病毒科、冠状病毒科和小核糖核酸病毒科的病毒都在粪便中大量存在。副粘病毒科在肾脏中最常见且丰度较高,仅在肾脏中发现了 40.7%。在肺、肝、肾和脾中检测到大量来自沙粒病毒科、动脉炎病毒科、黄病毒科和汉坦病毒科的病毒,但在粪便中未检测到这些病毒。最后,与很少发现的病毒(例如杯状病毒、脊髓病毒)相比,常见的病毒(例如星状病毒、冠状病毒、副粘病毒和小核糖核酸病毒)往往具有多器官分布(图3B)。
值得注意的是,即使来自同一科的病毒在不同哺乳动物中也具有不同的宿主内分布(图3B)。当病毒在主要靶器官中的丰度达到一定水平时,在其他内脏器官中也检测到了病毒(图3C)。例如,当粪便中的星状病毒科、冠状病毒科和小核糖核酸病毒科的病毒丰度分别达到每百万次读取数 (RPM) 101.0、100.72和 100.63时,也在肺、肝、肾和脾脏中检测到病毒(图3C)。对副粘病毒科病毒也进行了类似的观察,病毒从肾脏“溢出”到其他内脏器官的阈值约为 101.05 RPM。除主要靶器官外,脾脏的“溢出”病毒检出率是第二高(41.5%)。
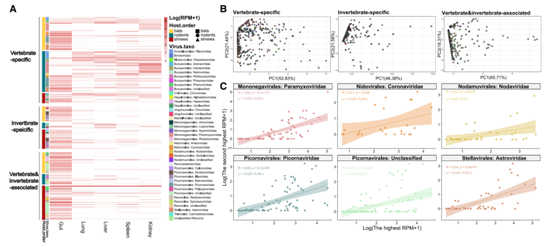
图3. 野生小型哺乳动物中脊椎动物和无脊椎动物相关病毒的器官分布。
(A)热图显示病毒在五种内脏器官的丰度。(B)主成分分析(PCA)图显示了病毒在蝙蝠、啮齿动物和鼩鼱中的多器官分布。(C)主要靶点与次要器官之间的病毒丰度关系。
04
野生小型哺乳动物病毒的多样化和进化
对此处鉴定的病毒的系统发育分析表明,蝙蝠、啮齿动物和鼩鼱拥有高度多样性的哺乳动物病毒,包括具有进化意义的病毒(图4)。例如,在五峰黑腹田鼠中鉴定出的一种新型汉坦病毒(五峰啮齿动物汉坦病毒)与已知的汉坦病毒不同(氨基酸[aa]差异>30%),但与鼩鼱中的病毒(thottimviruses)形成了姐妹分类单元。一种蝙蝠沙粒病毒(Wufeng bat mammarenavirus 1),是第一个在蝙蝠中发现的旧大陆沙粒病毒,它与2000公里外云南省的一种小家鼠体内发现的琉球mammarenavirus 有密切的进化关系。在小核糖核酸病毒科中,两种新的鼩鼱相关小核糖核酸病毒,五峰鼩鼱小核糖核酸病毒5和温州鼩鼱小核糖核酸病毒1,分别与已建立的副病毒和双病毒形成了不同的姐妹病毒系。
他们还发现了大量曾认为是无脊椎动物特有或相关的病毒。根据它们在系统树中的位置以及它们在哺乳动物体内和之间的分布,这些病毒可大致分为四类(图4)。特别值得注意的是,第一类包括首次在哺乳动物中发现的病毒,并且以相对较高的丰度存在于多个物种和器官中——这表明这些哺乳动物是这些病毒的真正宿主,而不是宿主饮食的一个组成部分。例如,温州鼠丘状病毒1型在家鼠和棘鼠肺组织中均检出,阳性率和丰度均较高。
第二类包括在已建立的无脊椎动物相关病毒群中聚集在一起并形成不同分支的病毒,例如在来自不同地点的多个蝙蝠物种中鉴定出的新型荆楚病毒分支,这些病毒形成了与其余荆楚病毒目的氨基酸同一性<40%的不同谱系。
第三类病毒与之前在无脊椎动物中发现的病毒有非常密切的关系。例如,在五峰啮齿动物中鉴定出与蜱相关的永嘉Ledantevirus病毒,与参考序列的氨基酸一致性为98.7%。在啮齿动物和鼩鼱中发现的荆门啮齿动物白垩四病毒1型与先前在蜘蛛中发现的贵阳白垩四样病毒1型相似度为0.96%。
其余的病毒属于最后一类,它们相对较少。总体而言,这些无脊椎动物相关病毒中有 54.5% 取自鼩鼱,并且 >35% 具有多器官分布。
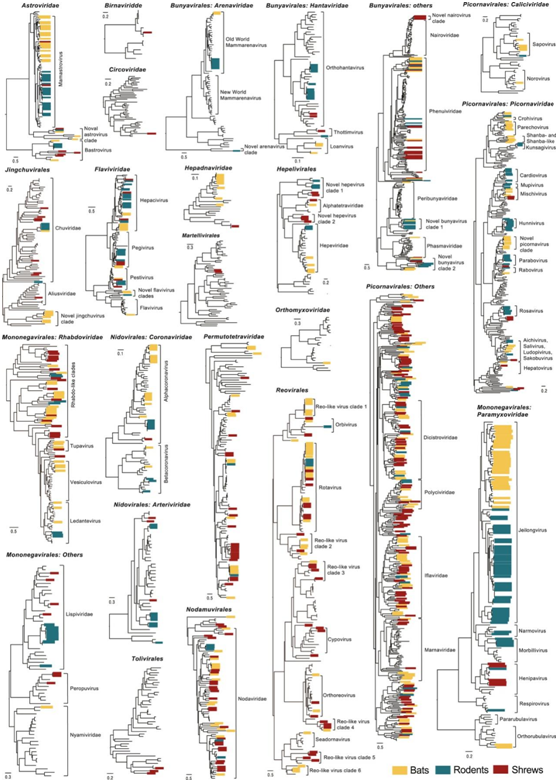
图4. 脊椎动物和无脊椎动物相关病毒的系统发育多样性。
根据RNA病毒的RdRp蛋白和DNA病毒的复制酶蛋白的氨基酸序列估计系统发育树。
05
病毒在野生小型哺乳动物中的传播
尽管在一种宿主物种中发现了 531 种新发现的病毒,但哺乳动物之间在种、属、科甚至目水平上可能发生跨物种传播事件。总共在 ≥ 2 个哺乳动物物种中鉴定出 138 种病毒,在 ≥ 2 个哺乳动物属中鉴定出 106 种病毒,在 ≥ 2 个哺乳动物科中鉴定出 49 种病毒,在两个或全部三个目哺乳动物中鉴定出 18 种病毒(图 5 A-B) 。
这里鉴定的跨物种传播病毒包括 29 个病毒分支(科或目),特别是来自副粘病毒科、小核糖核酸病毒科、黄病毒科、星状病毒科和黄病毒科的病毒(图 5 A)。值得注意的是,能够在哺乳动物目之间传播的18种病毒不仅包括脊椎动物相关病毒(例如肝炎病毒、沙粒病毒、亨尼帕病毒和星状病毒),还包括一些无脊椎动物相关病毒(例如双顺反子病毒和全四病毒)(图5A)。引人注目的是,轮状病毒 A拥有最广泛的哺乳动物宿主(8 个物种),在蝙蝠、啮齿动物和鼩鼱中共同循环。此外,啮齿动物和鼩鼱体内都存在三种新发现的肝炎病毒。然而,大多数跨物种传播事件(例如冠状病毒、汉坦病毒)发生在种或属水平。
为了更好地表征病毒在这些野生哺乳动物中的传播,他们构建了宿主-病毒相关网络(图5C)。对于蝙蝠和鼩鼱来说,病毒的跨物种传播似乎更频繁地发生在生活在同一栖息地的个体之间,而不是生活在不同栖息地的个体之间。然而,就啮齿动物而言,超过一半的跨物种传播病毒在来自不同地点的个体中观察到,这可能是由于它们的迁移和广泛的地理分布(图5C)。此外,啮齿类动物和鼩鼱更有可能拥有相同的病毒,两组中都传播着 13 种病毒(图 5 B )。
网络拓扑分析显示,1种蝙蝠(Myotis chinensis)、3种啮齿动物(Niviventer,N. fulvescens和Rattus norvegicus)和2种鼩鼱(C. smithii和C. attenuata)位于网络中心(图5C)。除了灰貂鼩外,其余的鼩鼱都携带至少两种交叉传播病毒。具体来说,鼩鼱(C. smithii和C. attenuata)都携带了五种跨物种传播病毒,这表明鼩鼱在病毒传播中起着不寻常的作用。
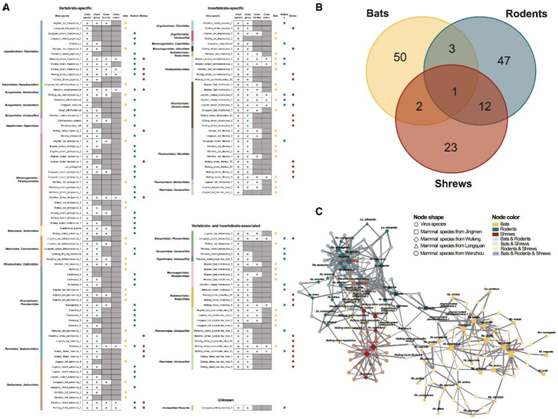
图5. 野生小型哺乳动物之间的病毒传播。
(A)病毒在宿主物种、属、科和目之间传播的概述。(B)显示蝙蝠、啮齿动物和鼩鼱中跨物种传播病毒数量的维恩图。(C)宿主-病毒关联网络。
06
控制病毒组组成和病毒跨物种传播的生态因素
为了进一步识别和排序野生小型哺乳动物中病毒组组成和病毒跨物种传播的生态驱动因素,他们使用广义线性模型(GLM)和广义加性模型(GAM)进行了全子集回归分析。
宿主顺序是影响病毒丰度的最重要因素,其次是影响病毒丰度的最重要因素,在两个模型中分别占了了32.1%和11.3%的总偏差。值得注意的是,鼩鼱对病毒丰富度和丰度有显著的积极影响(图6A-C),其病毒丰富度和丰度高于蝙蝠和啮齿动物(图6B-D)。这凸显了鼩鼱作为病毒宿主的重要性。宿主样本量也与病毒总丰度和丰度显著相关,样本量越大的哺乳动物往往携带更多的病毒,丰度也越高(图6A)。然而,在蝙蝠和啮齿动物中观察到显著的相关性,但在鼩鼱中没有(图6B-D)。此外,宿主栖息地影响病毒丰度,尽管影响各不相同(图6C)。温州、荆门农区鼠类病毒丰度明显高于龙泉、五峰山区鼠类病毒丰度(图6D)。然而,鼩鼱表现出相反的趋势,生活在山区的鼩鼱倾向于携带更多的病毒(图6D)。
预测跨物种传播病毒数量的最佳模型占了72.1% 的方差,包括总病毒丰富度、宿主顺序、宿主样本大小、宿主栖息地(海拔)和总病毒丰度(图 6 E)。总病毒丰富度占了最大的部分(占总偏差的24.5%),证明了较高的总病毒多样性与更频繁的病毒“溢出”相关。值得注意的是,啮齿动物的病毒“溢出”频率最高,其次是蝙蝠,与宿主效应兼容,独立于病毒的总多样性(图6F)。与此一致,宿主顺序和宿主样本量也是跨物种传播病毒数量的重要预测因子(图6E)。值得注意的是,啮齿动物与跨物种传播病毒的数量有显著的正相关(图6E)。
最后,他们使用混合GAM来确定与病毒是否能够跨物种传播相关的因素。最佳拟合模型占了20.8%的总偏差,包括宿主顺序、病毒分类、病毒总丰度和病毒是否多器官分布的影响。啮齿类动物中的病毒比蝙蝠和鼩鼱中的病毒具有更高的跨宿主物种潜力(图 6 G)。
在宿主的科水平的最佳拟合模型中(模型5),来自菊头科蝙蝠的phenuiviruses和来自Vespertillionidae科蝙蝠的弹状病毒在宿主物种之间跳跃的潜力明显更高(图6G)。此外,最佳模型还表明,丰度更高的病毒和分布在多个器官中的病毒更有可能溢出并感染更多的宿主(图 6G-H)。
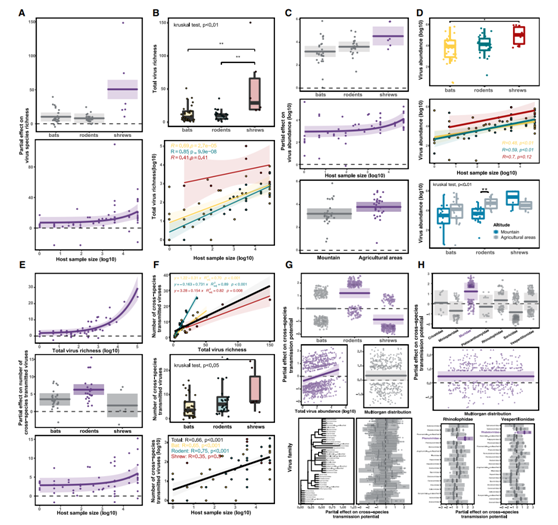
图6. 控制病毒组成和跨物种病毒传播的生态因素。
(A)病毒总丰度最佳拟合模型的部分效应图。(B)宿主样本量对每个宿主物种病毒总丰度的影响。(C)病毒总丰度最佳拟合模型的部分效应图。(D)宿主样本量和宿主栖息地对病毒总丰度的影响。(E)跨种传播病毒数量的最佳拟合模型的部分效应图。(F)病毒总丰度和宿主样本量对跨物种传播病毒数量的影响。(G)病毒跨种传播势最佳拟合模型的局部效应图。(H)病毒跨物种传播潜力的最佳拟合模型的部分效应图,包括宿主家族内嵌套的病毒分类的随机效应。
+ + + + + + + + + + +
结 论
本项研究通过对来自中国四个栖息地的2443份野生蝙蝠、啮齿动物和鼩鼱的内脏和粪便样本进行宏转录组测序,鉴定出669种病毒,其中包括534种新病毒,从而极大地扩展了哺乳动物病毒组。本项研究揭示了高水平的系统发育多样性,确定了跨物种病毒传播事件,阐明了病毒起源,并确定了哺乳动物宿主中的无脊椎动物病毒病例。宿主顺序和样本大小是影响病毒组组成和病毒溢出模式的最重要因素。鼩鼱携带着丰富度极高的病毒,包括许多与多器官分布的无脊椎动物相关的病毒,而啮齿类动物携带的病毒具有更强的宿主跳跃能力。这些数据突显了当地栖息地中哺乳动物病毒的显著多样性及其在新宿主中出现的能力。
+ + + + +



