

 English
English文献解读|Nat Microbiol(28.3):无细胞 DNA 中的噬菌体多样性可识别人类脓毒症病例中的细菌病原体
✦ +
+
论文ID
原名:Phage diversity in cell-free DNA identifies bacterial pathogens in human sepsis cases
译名:无细胞 DNA 中的噬菌体多样性可识别人类脓毒症病例中的细菌病原体
期刊:Nature Microbiology
影响因子:28.3
发表时间:2023.06.12
DOI号:10.1038/s41564-023-01406-x
背 景
噬菌体是感染细菌的病毒,在菌株和物种水平上对其细菌宿主具有很强的特异性。然而,噬菌体与相关细菌种群动态之间的关系尚不清楚。
实验设计
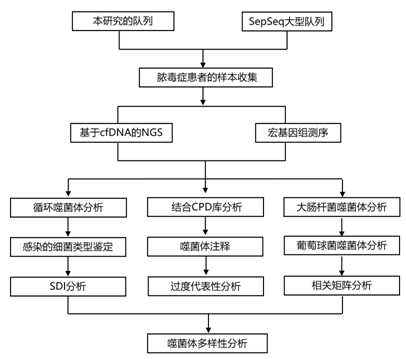
结 果
01

循环噬菌体反映感染病因
无细胞 DNA (cfDNA)适合非侵入性检测,并改变了围产期检测、自身免疫性疾病、癌症分期和移植排斥的诊断。研究者团队试图确定健康受试者中是否存在循环噬菌体,以及血培养阳性感染患者中是否存在循环噬菌体。为此,他们通过新一代测序 (NGS)分析了斯坦福大学急诊科 61 名脓毒症患者以及 10 名无症状对照者的血浆,从样本中收集cfDNA并在Illumina平台上进行测序,平均深度为1867万个读数。脓毒症样本代表了通过阳性血培养证实的 20 种不同的细菌感染,其中 3 个样本的多个细菌属培养结果呈阳性。
无症状对照和脓毒症患者的血浆 cfDNA 主要由人类读数组成(图1b)。不到 1% 的读数对应于哺乳动物病毒,而相当多的读数对应于噬菌体:无症状和脓毒症患者样本中的非人类读数分别为 8.80% 和 9.62%(图 1a)。利用完整的NCBI核苷酸数据库的BLAST搜索显示,尽管非人类读数占所有cfDNA读数的比例相对较小,但却代表了丰富的微生物群落(图1c)。受到攻击的细菌包括许多属。在无症状的个体中,最大的单一属是表皮菌,一种已知的皮肤共生菌。
本项研究的注释反映了多种噬菌体家族(图1d),尽管尾部的噬菌体家族以Myoviridae和Siphoviridae占主导地位。无症状对照组和脓毒症患者循环噬菌体中噬菌体家族代表性无统计学差异。Shannon多样性指数(Shannon Diversity Index, SDI)是衡量循环噬菌体物种丰富度和均匀度的常用生态量化指标,在两组之间没有显著差异(图1e),尽管脓毒症患者的噬菌体SDI范围确实更大,这些发现表明没有广泛的“脓毒症噬菌体”特征。在策划的噬菌体库(CPD)中已知宿主细菌的噬菌体中,大多数对应于假单胞菌、埃希氏菌、克雷伯氏菌、表皮菌和链球菌属(图1f),细菌宿主比例因脓毒症状态而无显著差异(图1f)。这些噬菌体的一部分是肠道宏基因组测序中未表征的肠道噬菌体,这表明肠道噬菌体对循环 cfDNA 库的潜在贡献。许多噬菌体有未知的细菌宿主,在无症状和脓毒症患者组中分别占噬菌体的47.2%和37.4%(图1f)。
由于脓毒症通常是由单一细菌菌株引起的,他们在每次感染的基础上研究了循环噬菌体——假设样本中与致病病原体相关的噬菌体将存在过度代表。他们发现,大肠杆菌、链球菌、葡萄球菌和克雷伯氏菌感染导致的脓毒症患者在病原菌对应的噬菌体比例中存在相应的过代表(图1g-k)。然而,单独的比例分析并不能说明每个样品中噬菌体的总数,也不能说明所有样品中与这些病原体相关的噬菌体群体的组成。因此,他们计算了噬菌体的生态多样性,发现大肠杆菌、链球菌、葡萄球菌和克雷伯氏菌感染引起的脓毒症患者中病原体特异性噬菌体群体的SDI增加(图1l-o)。来自多个细菌属的三个样本具有阳性培养物,证明了两种病原体的不同程度的代表性(图1g)。
总而言之,非人类 cfDNA 的含量,包括细菌、真核和病毒 DNA 以及噬菌体家族、多样性和注释的细菌宿主,在健康和脓毒症中大致相似。然而,血培养呈阳性的脓毒症患者的噬菌体组更能代表其脓毒症的细菌原因。
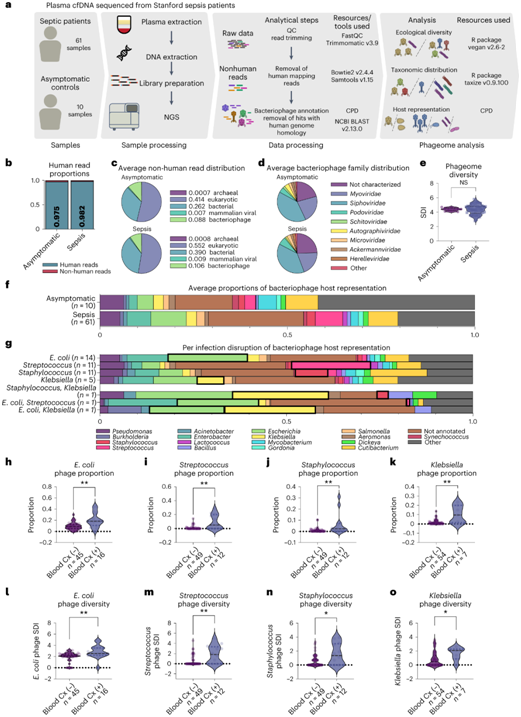
图1. 循环噬菌体反映感染病因。
(a) 样品数量、样品处理和分析方法示意图。 (b) 人类和非人类的比例。 (c) 按古细菌、真核生物、细菌、哺乳动物病毒和噬菌体类别划分的非人类读数的身份分布。 (d) 噬菌体家族的噬菌体分布。(e) 无症状和脓毒症患者噬菌体组中的 SDI 没有显著差异。 (f) 无症状和脓毒症患者噬菌体中细菌宿主属的独特噬菌体的平均分布。 (g) 具有单一识别病原体的样品中细菌宿主的独特噬菌体的平均比例。 (h-k) 小提琴图显示感染特异性噬菌体的比例。 (l-o) 小提琴图显示感染特异性的SDI 。
02
另一项大型研究中 cfDNA 噬菌体反映了病原体
他们试图在更大、临床更多样化的患者群体中验证本项研究结果。 SepSeq 研究是一项针对 391 名个体的队列研究,调查了如何使用细菌 cfDNA 来诊断患者脓毒症的病因,从而触发急诊科脓毒症警报,其数据具有广泛的传染性病因,且数据已公开。SepSeq 研究中有四个患者群体:(1) 血培养阳性脓毒症患者、(2) 血培养阴性脓毒症且其他部位有感染记录的患者、(3) 全身炎症反应综合征 (SIRS) 临床上出现脓毒症但最终未发现传染性病因的患者,以及(4)无症状对照(图 2a)。最初的研究报告了使用细菌 cfDNA 的整体有希望的诊断统计数据,但存在显著的差距:初始血培养阴性的样本敏感性不足,而对慢性感染和共生体过度敏感。首先,他们在本项研究的数据集中观察到的噬菌体组趋势是否与细菌感染患者的 SepSeq 研究中一致;其次,cfDNA 中存在的噬菌体是否可以在病原细菌生态学的研究中提供其他有用的信息。
他们发现,在健康和脓毒症中循环的噬菌体非常相似(图2a)。虽然脓毒症患者和血培养阳性患者的整体噬菌体多样性较高(图2b),但鉴定出的噬菌体所代表的细菌宿主比例基本稳定(图2c)。他们发现最常见的细菌宿主是肠杆菌、埃希氏菌、葡萄球菌和链球菌,即使在健康患者中也是如此(图2c)。与其他组相比,这些噬菌体中特异性细菌病原体的噬菌体再次发生过度代表(图 2d)。在血培养阳性的大肠杆菌、链球菌、葡萄球菌和克雷伯氏菌感染患者中,病原体相关噬菌体的比例(图21-h)和多样性(图21-l)均显著高于其他感染或未分离病原体的个体(SIRS)。有趣的是,对于单一细菌感染病因,患者的噬菌体比例和多样性在血培养阳性和血培养阴性组之间没有变化,这表明循环噬菌体序列能够反映菌血症之外的感染。这些发现可能表明潜在的未诊断感染、感染对体内其他细菌群落的影响,甚至炎症导致的噬菌体易位增加。
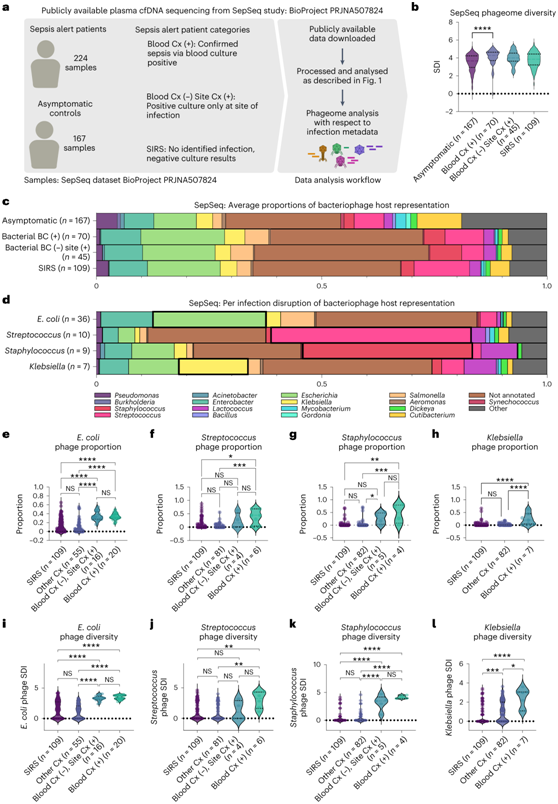
图2. 另一项大型研究中 cfDNA 噬菌体反映了病原体。
(a) SepSeq 示例示意图。(b) 按样本类别划分的噬菌体 SDI。(c) 按样品类别划分的细菌宿主的独特噬菌体的平均比例。(d) 每次感染破坏具有单一鉴定病原体的独特噬菌体的平均比例。(e-h) 按感染类别划分的感染特异性噬菌体比例。(i-l) 按感染类别划分的病原体宿主噬菌体 SDI。
03
大肠杆菌噬菌体反映宿主菌株的特征
大肠杆菌感染是脓毒症的常见原因,包括在本项研究的队列中。大肠杆菌噬菌体在无症状患者中是散发的,在SIRS患者或非大肠杆菌患者中更常见,在大肠杆菌脓毒症患者中普遍存在(图3a)。在大肠杆菌脓毒症患者中,Siphoviridae、Podoviridae和Myoviridae噬菌体家族的大肠杆菌噬菌体显著富集,而自噬病毒科噬菌体在感染状态下仍保持稳定的低水平(图3b-e)。在SIRS患者或非大肠杆菌脓毒症的患者中,大肠杆菌噬菌体中Siphoviridae、Podoviridae和Myoviridae噬菌体家族的噬菌体也有显著的富集(图3b-e)。
在36名大肠杆菌脓毒症患者中发现了500多种独特的噬菌体。在许多大肠杆菌脓毒症患者中都存在适度代表的噬菌体亚群。剩下的噬菌体仅存在于少数样品中——事实上,大多数噬菌体仅存在于一两个患者中(图3f)。然后,通过上述基于流行率的群体分析了单个噬菌体的特征。他们发现大肠杆菌噬菌体家族(因此形态学)的差异与噬菌体在患者中的常见程度相对应。广泛代表的大肠杆菌噬菌体具有明显的Siphoviridae噬菌体富集; Podoviridae噬菌体在中等代表性的噬菌体亚群中占主导地位;罕见的噬菌体亚群主要是Myoviridae噬菌体(图3g)。
利用CPD,他们将细菌宿主标注为大肠杆菌噬菌体在每个流行亚群中的菌株水平,发现大肠杆菌的实验室菌株,大多数最初来自患者样本,是每个噬菌体亚群的共同宿主。与产毒大肠杆菌菌株(STEC和ETEC)相关的噬菌体在广泛代表的噬菌体组中存在过度代表(图3h)。
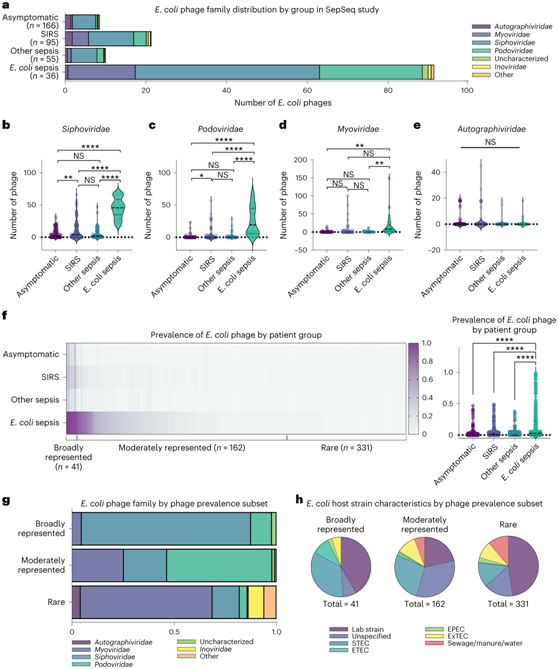
图3. 大肠杆菌噬菌体反映宿主菌株特征。
(a) 大肠杆菌噬菌体反映宿主菌株特征。(b-e) 通过 Brown–Forsythe 和 Welch ANOVA分析以及大肠杆菌噬菌体数量分析。(f) 一些单独的大肠杆菌噬菌体在多个样本中普遍存在,其中大肠杆菌噬菌体脓毒症样本反映噬菌体在整个组中的代表性更高。 (g) 按大肠杆菌脓毒症中的代表性水平按大肠杆菌噬菌体子集组划分的样品中噬菌体代表性的堆积条形图。(h) 按噬菌体代表组划分的大肠杆菌噬菌体形态。
04
噬菌体在葡萄球菌感染中指定细菌种类
在后续的研究中,他们提出了一个问题:噬菌体是否可以帮助恢复一些失去的细菌DNA的“分辨率”?于是,他们评估了金黄色葡萄球菌和凝固酶阴性葡萄球菌之间的噬菌体生态学是否存在差异,金黄色葡萄球菌在血液培养中的存在通常是致病的,而凝固酶阴性葡萄球菌(CoNS)的存在通常是一种污染物。这两个物种共有许多核心基因,如果在cfDNA中检测到这些基因,就很难确定它们属于哪一个。
当定位到金黄色葡萄球菌参考基因组时,无论是本项研究的队列(图4a)还是SepSeq队列(图4b),细菌cfDNA都无法区分金黄色葡萄球菌和金黄色葡萄球菌。与此相一致的是,相应的受试者工作特征(ROC)图的曲线下面积(auc)接近0.5(图4c)。然而,感染金黄色葡萄球菌和CoNS的噬菌体种群是不同的,金黄色葡萄球菌特异性噬菌体的多样性提供了信息,因为金黄色葡萄球菌噬菌体多样性仅在金黄色葡萄球菌感染中显著增加(图4e)。这反映在AUC为0.96的ROC图中(图4f)。虽然基于噬菌体多样性的AUC高于基于细菌DNA的AUC(图4f),但这种增加在本项研究的较小队列中没有达到显著性(图4d),需要在一个单独的、更强大的队列中进一步验证。
这些结果表明,金黄色葡萄球菌噬菌体多样性的增加对金黄色葡萄球菌感染是敏感和特异性的。金黄色葡萄球菌噬菌体多样性与金黄色葡萄球菌感染密切相关,而在CoNS噬菌体多样性与CoNS感染中则没有这种关系(图4g)。总之,噬菌体亚群,特别是金黄色葡萄球菌噬菌体,是比较相关细菌物种感染的潜在目标,其中遗传相似的病原体使NGS分析复杂化。
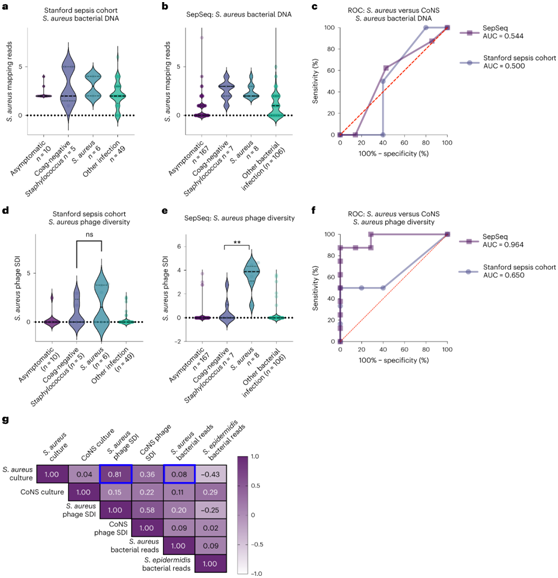
图4. 噬菌体指定葡萄球菌感染中的细菌种类。
(a) 按感染类别读取斯坦福脓毒症队列中金黄色葡萄球菌细菌基因组的图谱。(b) 按感染类别读取SepSeq 中金黄色葡萄球菌细菌基因组的映射。(c) 金黄色葡萄球菌细菌基因组在区分金黄色葡萄球菌培养物与CoNS培养物中的ROC图。(d) 斯坦福脓毒症队列中按感染类别划分的金黄色葡萄球菌噬菌体 SDI。(e) SepSeq 中按感染类别划分的金黄色葡萄球菌SDI 。(f) 金黄色葡萄球菌噬菌体 SDI 区分金黄色葡萄球菌培养物和 CoNS 培养物的 ROC 图。(g) SepSeq 样品中培养状态、噬菌体多样性和金黄色葡萄球菌图谱细菌读数的相关矩阵。
+ + + + + + + + + + +
结 论
本项研究分析了血浆样本中无细胞 DNA 中与噬菌体及其细菌宿主相关的序列。对两个独立队列(包括由 61 名脓毒症患者和 10 名对照者组成的斯坦福队列以及由 224 名脓毒症患者和 167 名对照者组成的 SeqStudy 队列)的分析揭示了所有样本个体血浆中存在循环噬菌体组。此外,感染与病原体特异性噬菌体的过度表达有关,从而可以识别细菌病原体。有关噬菌体多样性的信息可以识别产生这些噬菌体的细菌,包括大肠杆菌的致病变异菌株。噬菌体序列同样可用于区分密切相关的细菌种类,例如金黄色葡萄球菌和凝固酶阴性葡萄球菌。噬菌体无细胞 DNA 可能有助于研究细菌感染。
+ + + + +



