

 English
English文献解读|Nature(48.5):肠道菌群介导的米色脂肪细胞对饮食信号的诱导
✦ +
+
论文ID
原名:Microbiota-mediated induction of beige adipocytes in response to dietary cues
译名:肠道菌群介导的米色脂肪细胞对饮食信号的诱导
期刊:Nature
影响因子:48.5
发表时间:2026.03.04
作者/通讯作者:Takeshi Tanoue/ Kenya Honda
单位:日本东京庆应义塾大学医学院微生物与免疫学系
DOI号:10.1038/s41586-026-10205-3
背 景
饮食是宿主生理的基本调节因子,影响代谢、免疫以及对多种疾病的易感性。肠道菌群已成为饮食效应的关键介质,导致个体对营养和分子输入的反应在性质、方向和强度上存在差异。在众多受饮食-菌群相互作用影响的生理过程中,脂肪组织重塑的调控已成为一个引人入胜但尚未得到充分研究的领域。与富含线粒体并表达产热蛋白[如解偶联蛋白1 (Ucp1)]的棕色脂肪组织 (BAT) 不同,白色脂肪组织 (WAT) 储存过剩能量。然而,值得注意的是,WAT 具有相当大的可塑性,在特定条件下(例如暴露于寒冷环境和儿茶酚胺刺激)可以获得类似棕色脂肪的特性。这一过程为白色脂肪棕色化(米色化),其特征是线粒体生物合成增强,并表达产热和脂质氧化相关基因。饮食与肠道菌群的相互作用对代谢健康至关重要,影响着能量平衡和疾病易感性。然而,饮食和微生物因素如何共同调节宿主生理的潜在机制仍不清楚。
实验设计
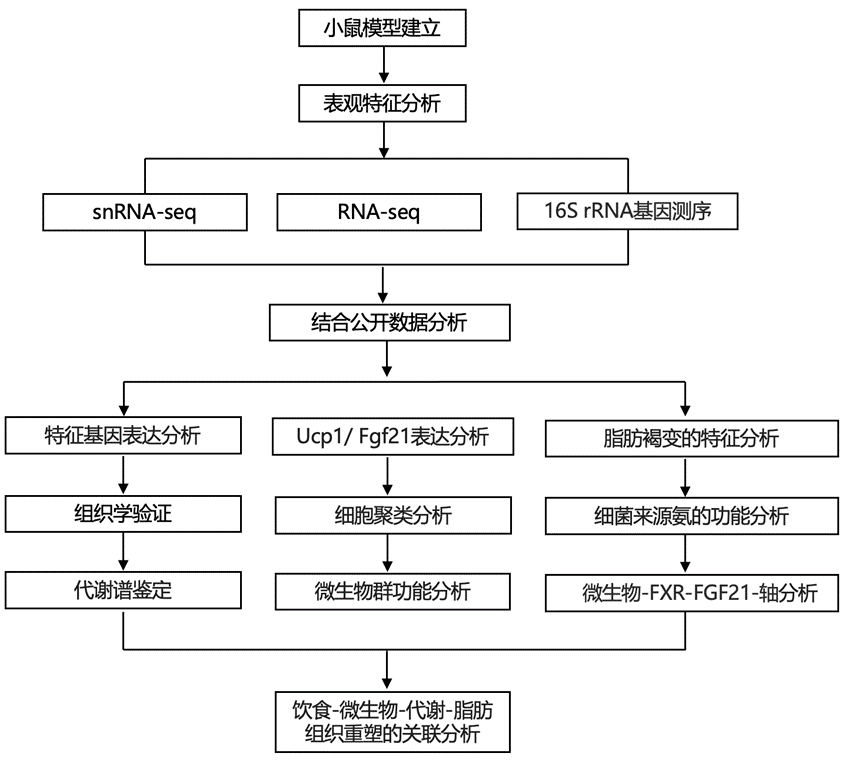
结 果
01

LPD促进白色脂肪组织褐变
研究团队首先通过配制不同蛋白质、碳水化合物和脂肪比例的饮食,研究了饮食调整对WAT褐变的影响。虽然脂肪含量的变化会影响总热量,但所有其他饮食均设计为等热量。将这些饮食喂给饲养在室温(23℃)下的无特定病原体清除(SPF)C57BL/6(B6)小鼠。无论脂质和碳水化合物含量如何,喂食低蛋白饮食(LPD)均显著增加了小鼠腹股沟白色脂肪组织(iWAT)中Ucp1和Cox7a1 mRNA的表达。mRNA表达采用三种不同的方法进行评估,结果一致。LPD喂养小鼠导致iWAT质量下降,并增加棕色化组织学特征,包括多房性脂滴细胞,尤其是在腹股沟淋巴结附近的中央区域。喂食不同蛋白质含量饮食的B6小鼠,当饮食蛋白质含量降低至7%或更低(比对照组低约60%)时,iWAT中Ucp1、Elovl3和Cox7a1的表达显著上调(图1b)。时间进程分析表明,这些基因在7%蛋白质饮食喂养2周内即发生诱导,并在6-8周达到平台期(图1c)。转录组分析(RNA-seq)分析显示,iWAT中多种米色脂肪特征基因表达上调,包括参与线粒体功能(如Cox7a1和Cox8b)、产热(如Elovl3、Dio2和Ucp1)以及脂质代谢(如Cidea、Dio2、Cpt1b和Acott11)的基因;这种上调在LPD喂养后1周内开始,并在接下来的6周内逐渐增强。LPD喂养诱导的iWAT转录谱与β3-肾上腺素能受体激动剂诱导的转录谱非常相似。膳食蛋白质含量的降低也导致肩胛间棕色脂肪组织中Ucp1、Elovl3和Cox7a1的表达增加,尽管其幅度低于在 iWAT 中观察到的。
LPD即使在热中性条件(30 °C)下也能显著诱导米色脂肪的特征基因表达。在包括BALB/c和ICR在内的多个小鼠品系中均观察到LPD诱导的iWAT中米色脂肪细胞的形成,因此并非B6小鼠特有。然而,与雄性或幼年小鼠相比,雌性或老年B6小鼠的棕色化程度有所减弱。此外,性腺周围WAT未表现出米色标记基因的诱导。恢复正常饮食后,米色标记基因的表达和米色脂肪细胞样形态均有所下降,表明LPD诱导的iWAT棕色化是可诱导且可逆的。当小鼠从高脂饮食(HFD)转为LPD或高脂低蛋白饮食(HFD转HF/LPD)时,也观察到Ucp1表达增加和代谢效应,但随着脂肪暴露时间延长或脂肪暴露量增加,这种诱导作用的强度减弱。总的来说,这些结果表明,LPD诱导的脂肪褐变作用显著,但具有脂肪组织特异性,并受年龄、性别和饮食环境的调节。
由于蛋白质类型和消化率可能影响结果,他们接下来研究了含有特定氨基酸而非天然蛋白质的日粮。降低日粮氨基酸含量导致iWAT中米色脂肪标记基因表达显著增加,在氨基酸含量为5%或更低时,诱导作用达到最大值,与7% LPD的效果相当(图1d)。随后,他们配制了必需氨基酸(EAA)或非必需氨基酸(NEAA)含量降低的日粮。降低EAA含量重现了总氨基酸限制条件下观察到的米色脂肪标记基因的诱导作用,而限制NEAA含量则没有明显影响(图1e)。接下来,他们研究了降低日粮中每种EAA含量的影响。异亮氨酸、亮氨酸、苯丙氨酸、色氨酸、赖氨酸、蛋氨酸或苏氨酸的减少导致褐变程度轻微且不一。这些结果表明,虽然限制单个必需氨基酸的影响有限,但同时限制多种必需氨基酸会激活互补通路,从而驱动更强的褐变反应。
接下来,他们研究了LPD介导的腹股沟iWAT褐变的机制。给小鼠喂食LPD可显著诱导Ucp1、Elovl3和Cox7a1的表达,其水平与冷暴露(6℃,7天)或β3-肾上腺素能受体激动剂处理所诱导的水平相当(图1f)。值得注意的是,LPD与冷暴露相结合会导致米色脂肪标记基因表达的叠加增加,这表明褐变涉及不同的(但可能存在重叠的)机制。尽管免疫细胞与冷诱导的褐变有关,但LPD诱导的褐变在多种免疫缺陷小鼠模型中均得以保留,包括缺乏αβ和γδ T细胞、B细胞、固有淋巴细胞、I型、II型或I7型免疫分子、髓系细胞或淋巴组织的小鼠。因此,他们重点研究了非免疫依赖性机制。
接下来,他们使用无菌(GF)小鼠评估了肠道菌群在LPD诱导的脂肪褐变中的作用。与SPF小鼠相比,GF小鼠在喂食LPD后,Ucp1、Elovl3和Cox7a1的表达显著降低,且iWAT的组织学褐变指标也较少(图1g-h)。此外,虽然LPD抑制了SPF小鼠的体重增加和WAT质量,但在GF小鼠中未观察到这种效应(图1i-j)。在喂食LPD的GF小鼠中偶尔观察到Ucp1的低水平诱导,提示存在一条不依赖于肠道菌群的通路,尽管这种反应并不显著。对LPD喂养的GF小鼠的iWAT进行RNA-seq发现,与LPD喂养的SPF小鼠相比,多种米色脂肪组织特征基因的表达降低,包括Ucp1、Cidea、Elovl3、Cox7a1、Cox8b、Clstn3和Acot11(图1k)。GO富集分析表明,与SPF小鼠相比,LPD喂养的GF小鼠中参与线粒体生物合成和脂质代谢的基因表达降低。在低必需氨基酸(EAA)饮食喂养的小鼠中也观察到了类似的趋势。用抗生素混合物处理成年SPF小鼠也导致iWAT中Ucp1、Elovl3和Cox7a1的表达显著降低。这些研究结果表明,微生物群是 LPD 介导的米色脂肪细胞诱导的关键驱动因素。
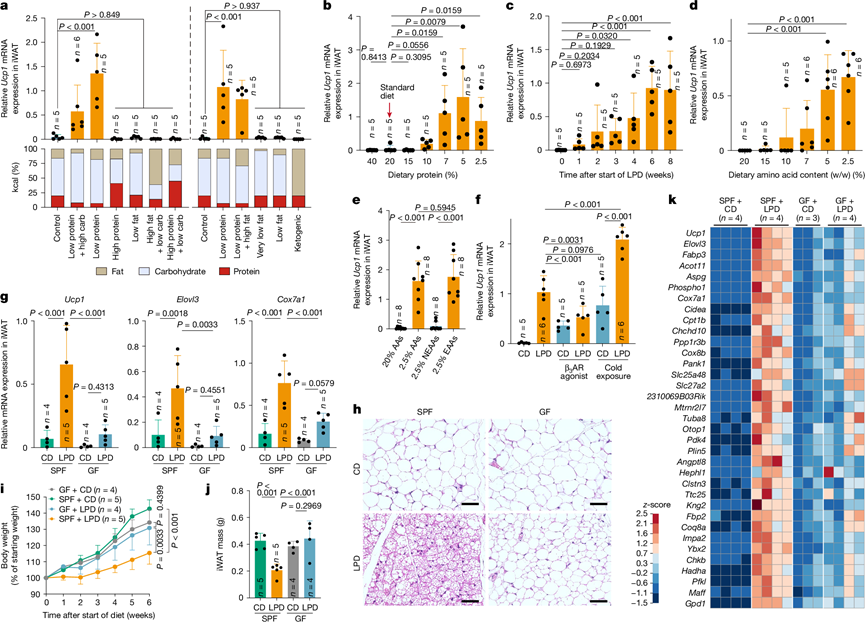
图1. LPD以微生物群依赖的方式诱导褐变。
(a) SPF B6雄性小鼠喂食不同比例的蛋白质、碳水化合物和脂肪的饲料,持续6周。图中显示了iWAT中Ucp1 mRNA的表达水平(以Ppib为内参进行标准化),以及每种饲料的宏量营养素组成。(b-g) 小鼠iWAT中Ucp1 mRNA的相对表达水平。(h) 代表性的苏木精-伊红(H&E)染色切片。(i-j) 体重变化和iWAT质量。(k) 热图显示与SPF + CD、GF + CD和GF + LPD小鼠相比,SPF + LPD小鼠iWAT中富集的基因。
02
LPD-微生物群-FXR通路在褐变中的作用
接下来,他们旨在鉴定可能促进iWAT褐变的微生物群来源分子。对喂食对照饮食或LPD的SPF和GF小鼠的回肠和血浆样本进行非靶代谢谱分析,结果显示,LPD增加了SPF小鼠血浆中非结合胆汁酸的水平,包括胆酸(CA)和鼠胆酸(MCA)。后续的靶向液相色谱-质谱(LC-MS)分析证实,与GF小鼠和对照饮食喂养的SPF小鼠相比,LPD喂养的SPF小鼠血浆中CA、α-甲基胆酸(αMCA)、β-甲基胆酸(βMCA)和鹅脱氧胆酸(CDCA)的水平升高,同时7α-和7β-脱羟基化产物[包括7-氧代脱氧胆酸(7oxoDCA)、熊胆酸(UCA)和熊去氧胆酸(UDCA)]的水平也升高(图2a),尽管在肠道内容物中未观察到类似的趋势。这些胆汁酸在体外FXR报告基因检测中表现出激动活性,表明该胆汁酸受体发生激活。因此,他们接下来研究了FXR信号通路是否参与了胆汁褐变过程。与野生型对照组相比,FXR(由Nr1h4编码)缺陷小鼠在LPD喂养期间,Ucp1和Elovl3的诱导显著降低,iWAT中形态学上的米色脂肪细胞数量减少,体重增加(图2b)。相反,另一种重要的胆汁酸受体TGR5的缺失(Gpbar1-/-)并不影响LPD诱导的脂肪褐变(图2c)。这些结果表明,肠道菌群来源的胆汁酸在LPD喂养期间促进米色脂肪细胞的诱导,至少部分是通过FXR依赖性、TGR5非依赖性途径实现的。
鉴于FXR在肠道、肝脏和WAT中广泛表达,他们利用组织特异性Nr1h4敲除小鼠来确定FXR信号通路对iWAT棕色化的器官特异性贡献。在肠道上皮细胞(Nr1h4fl/fl ; Vil1-cre)或肝细胞(Nr1h4fl/fl; Alb-cre)中敲除FXR并不影响LPD诱导的Ucp1表达。相反,脂肪细胞特异性敲除FXR(Nr1h4 fl / fl; Adipoq-cre)显著降低了iWAT中Ucp1和Elovl3的表达以及形态学上的米色细胞(图2d),表明FXR在脂肪组织中发挥关键作用。为了鉴定iWAT中表达FXR的细胞类型,他们对喂食对照饮食或LPD的GF和SPF小鼠的iWAT进行了单细胞核转录组(snRNA-seq)。在LPD喂养的SPF小鼠中几乎只检测到表达Ucp1和Elovl3的假定米色细胞聚类(图2e)。在胆汁酸受体中,FXR的表达模式较为局限,并且在表达脂肪干细胞和祖细胞(ASPC)标志物(包括Wnt2和 Dpp4)的细胞中富集(图2e-f)。在Dpp4+细胞中条件性敲除FXR(Nr1h4 fl/fl;Dpp4-cre)显著减弱了LPD喂养后Ucp1和Elovl3的诱导(图2g)。伪时序分析进一步表明,细胞从头分化为米色细胞,其特征是线粒体脂质代谢和产热基因的顺序诱导。
值得注意的是,Fgf21(另一种ATF4诱导基因)在SPF小鼠肝脏中显著上调,但在GF小鼠中未见此现象,且在LPD喂养开始后1周即可观察到,同时循环FGF21水平也相应升高(图2h)。Fgf21-/-小鼠的表型与GF小鼠相似,在iWAT中米色脂肪标记基因的诱导程度极低,且在LPD喂养期间体重增加未受到抑制(图2i-j)。然而,给予重组FGF21不足以诱导正常饮食喂养的GF小鼠iWAT褐变,表明还需要其他依赖于肠道菌群的信号,包括胆汁酸-FXR轴。事实上,Fgf21−/−和Nr1h4 −/−小鼠的 iWAT 基因表达谱呈现出不同的模式,并且Nr1h4和Klb(编码 FGF21 受体的关键组分)在 iWAT 内不同的细胞群中表达。此外,LPD喂养后,Fgf21−/−小鼠和野生型小鼠的血浆胆汁酸谱相似(图2k),相反,Nr1h4−/−小鼠和野生型小鼠的血浆 FGF21 水平没有显著差异(图2l)。这些结果表明,FXR 和 FGF21 通过平行且非冗余的信号通路发挥作用,共同促进 iWAT 中米色细胞的诱导。
由于儿茶酚胺信号传导在脂肪褐变中起着核心作用,他们接下来评估了LPD介导的β1(Adrb1)、β2(Adrb2)或β3(Adrb3)肾上腺素能受体缺陷小鼠的米色脂肪细胞诱导情况。β3肾上腺素能信号传导的缺失,无论是单独缺失还是与β1或β2受体缺失联合缺失,均导致米色脂肪细胞诱导严重受损,而β1和β2受体的缺失则没有影响(图2m),这表明LPD诱导的脂肪褐变特异性地依赖于β3肾上腺素能受体信号传导。对iWAT进行全组织免疫染色显示,在LPD处理后,SPF小鼠(而非GF野生型小鼠)的酪氨酸羟化酶阳性交感神经元和血管发生了显著重塑(图2n)。具体而言,交感神经元形成了更精细、更密集的网络,尤其是在Ucp1表达细胞高度聚集的区域。相比之下,LPD喂养的Fgf21-/-和Nr1h4-/- SPF小鼠的交感神经支配显著减少,与GF小鼠的模式相似(图2n)。值得注意的是,给予β3-肾上腺素能受体激动剂后,喂食对照饮食的GF小鼠以及Nr1h4-/-和Fgf21-/-小鼠中Ucp1和Cox7a1的表达水平显著升高,达到与喂食LPD的SPF小鼠相当的水平(图2o-q)。这些结果表明,FXR和FGF21通过不同的信号通路发挥作用,但这些通路最终共同促进LPD诱导的褐变过程中交感神经支配和β3-肾上腺素能信号的激活。
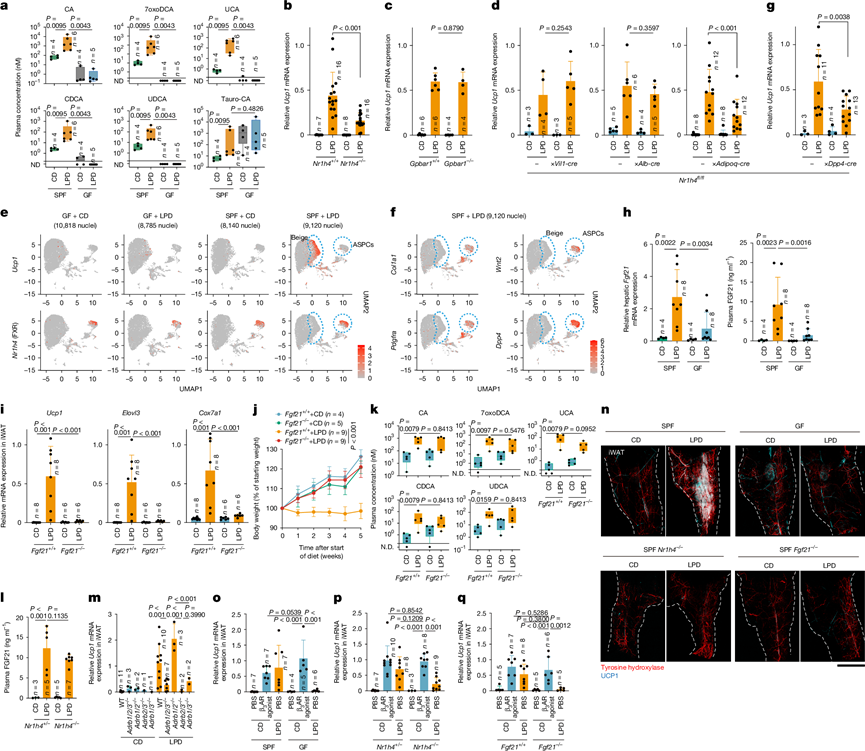
图2. iWAT FXR 和肝脏 FGF21 促进 LPD 介导的棕色化。
(a) 采用液相色谱-串联质谱法(LC-MS/MS)定量分析喂食标准饲料(CD)或LPD的GF或SPF小鼠的血浆胆汁酸水平。(b-d, g) iWAT中Ucp1的表达水平。(e-f) iWAT中Ucp1和Nr1h4以及Col1a1、Wnt2、Pdgfra和Dpp4的snRNA-seq数据的均匀流形近似投影(UMAP)图。(h) 检测肝脏Fgf21 mRNA表达和血浆FGF21水平。(i-k) iWAT基因表达、体重变化和血浆胆汁酸水平。(l) 小鼠的血浆FGF21水平。(m) iWAT中Ucp1 mRNA的表达情况。(n) 代表性的iWAT全组织图像。(o-q) iWAT中Ucp1的相对表达量。
03
20种细菌菌株诱导小鼠褐变
为了评估LPD诱导的iWAT褐变是否可通过肠道菌群传播,他们将LPD喂养的SPF供体小鼠(供体A和B)的小肠腔内容物移植到GF小鼠体内(图3a)。无论移植来源如何,受体小鼠均观察到褐变,但仅当受体小鼠喂食LPD时才会出现褐变(图3b),表明持续的饮食-肠道菌群相互作用是必需的。使用粪便移植也获得了类似的结果。为了鉴定能够介导褐变的细菌群落,他们选择了小鼠B28,该小鼠在接受LPD喂养的小鼠B的小肠内容物并继续喂食LPD后,Ucp1的诱导作用最强(图3b)。将小鼠 B28 的小肠内容物转移到新的无菌受体小鼠体内,重现了LPD喂养后 iWAT 的显著褐变(图3c)。他们选择表现出最强米色脂肪细胞诱导的小鼠 B28-1 进行后续分析(图3c)。从小鼠 B28-1 的小肠内容物中,分离出 18 株细菌(mu18-mix)(图3d)。然而,用 mu18-mix 定植无菌小鼠不足以诱导褐变(图3e)。添加另外两种分离株,即Blautia sp. (St.27G3) 和Turicibacter sp.,则未能诱导褐变。(St.80E1)菌株构建了一个包含20个菌株的菌群(mu20-mix),该菌群在LPD喂养后能显著诱导褐变,其水平与SPF小鼠相当(图3f-g),表明这两个菌株是该菌群发挥完全米色细胞诱导活性所必需的。接种mu20-mix并喂食LPD的GF小鼠血浆中CA、CDCA、7oxoDCA、UCA和UDCA水平升高(图3h),FGF21水平也升高(图3i)。相反,接种mu20-mix的Nr1h4 -/-和Fgf21-/- GF小鼠的米色细胞诱导显著降低(图3j),进一步支持了特定微生物群成员介导的LPD诱导褐变需要FXR和FGF21信号通路。
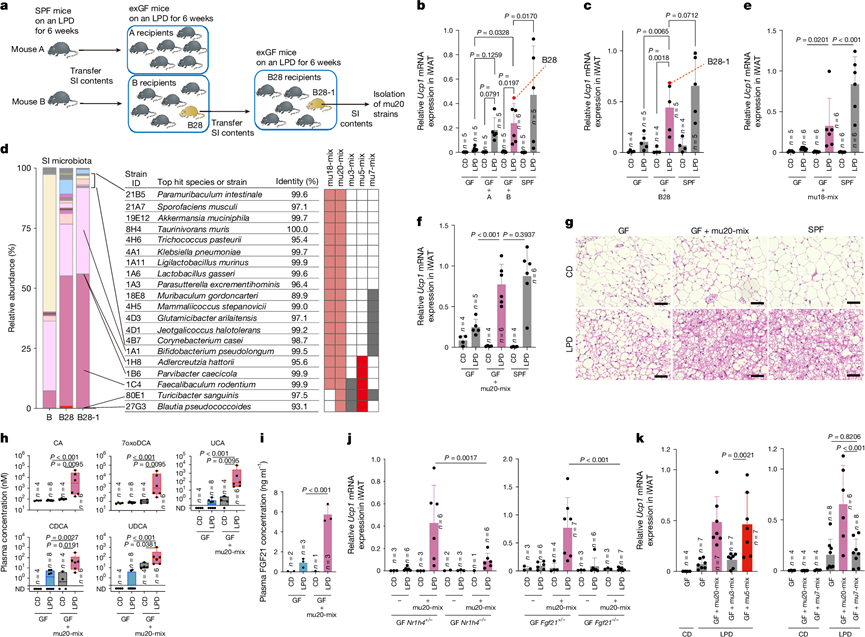
图3. 小鼠来源的微生物分离物促进 LPD 诱导的褐变。
(a) 示意图展示了从SPF小鼠肠道菌群中分离棕色化诱导共生菌株的策略。(b-c, e-f) iWAT中Ucp1的相对表达量。(d) 通过16S rRNA测序确定小鼠B、小鼠B28和小鼠B28-1的回肠菌群组成。(g) 来自指定组的代表性H&E染色iWAT切片。(h-i) 分别采用液相色谱-串联质谱法(LC-MS/MS)和酶联免疫吸附试验(ELISA)定量分析。(j) 检测iWAT中Ucp1的表达。(k) 检测iWAT中Ucp1的表达和肝脏中Fgf21的表达。
04
四种人源分离物促进褐变
在确认少量微生物群落足以促进LPD诱导的脂肪褐变后,他们接下来试图鉴定能够发挥此作用的人体相关微生物。为此,他们招募了25名健康志愿者,并进行了18F-氟代脱氧葡萄糖正电子发射断层扫描(FDG-PET),该扫描可检测米色或棕色脂肪组织。约40%的志愿者表现出锁骨上FDG聚集(图4a)。他们将FDG阳性率最高的四名志愿者(T17、T10、T19和T18)的粪便样本移植到GF小鼠体内,并将小鼠分别喂以对照饮食或LPD。在用来自供体T17、T10或T19的肠道菌群定植的exGF小鼠中,LPD显著诱导了iWAT中Ucp1的表达和形态学上的米色脂肪细胞(图4b)。来自供体T18的粪便肠道菌群移植效果则差得多,这可能是由于关键效应菌株的移植失败所致。他们还用来自供体FF2和T07的粪便样本移植了GF小鼠,这两位供体分别表现出中等程度或无FDG积累。与人类表型相似,FF2受体小鼠表现出中等程度的脂肪褐变,而T07受体小鼠则未见脂肪褐变迹象(图4c)。
接下来,他们从接受T10微生物群的小鼠(T10-4和T10-5)的粪便样本中分离出33株菌株,并从接受T19微生物群的小鼠(T19-5和T19-6)的粪便样本中分离出另外33株菌株(图4b-h)。他们将这33株菌株的混合物分别接种到GF小鼠中。接种T19来源的33株菌株混合物的小鼠在LPD喂养后表现出显著的米色细胞诱导,而接种T10来源的混合物的小鼠的诱导则明显较弱(图4d)。接种T19来源的33株菌株混合物的小鼠血浆中CA、7oxoDCA、UCA、CDCA和UDCA以及FGF21的水平升高(图4e-f)。相比之下,接种了 T19 衍生的混合物的Nr1h4 −/−或Fgf21−/− GF 小鼠未能显著上调 iWAT 中的Ucp1(图4g),表明这些品系通过激活 FXR 和 FGF21 通路促进米色细胞的诱导,类似于 mu20 混合物。
为了鉴定最小效应菌群,他们将33株T19衍生菌株分为两个系统发育群:一个包含19株菌株的菌群(19 BEEO),包括拟杆菌属、肠球菌属、丹毒丝菌科和其他门;以及一个包含14株菌株的菌群(14 RL),包括瘤胃球菌科和毛螺菌科(图4h)。用19 BEEO菌群定植的无菌小鼠重现了亲代33株菌群在iWAT中Ucp1的强诱导表达,而用14 RL菌群定植的小鼠则未观察到此现象(图4i)。他们进一步将33株T19衍生菌株细分为5个系统发育群(图4h),并评估了排除每个系统发育群对iWAT褐变的影响。排除归类为其他门类的5株菌株(5 Others)后,Ucp1诱导的降低最为显著(图4j)。因此,他们比较了5 Others混合菌株与19株BEEO组中剩余14株菌株(14 BEE)定植的效果。5 Others混合菌株以LPD依赖的方式有效诱导米色标记基因的表达,而14 BEE菌株仅产生轻微的效果(图4k)。5 Others混合菌株(而非14 BEE混合菌株)定植还能使肝脏Fgf21 mRNA表达和血浆胆汁酸浓度升高至与定植亲代19 BEEO混合菌株的小鼠相当的水平(图4l-m)。
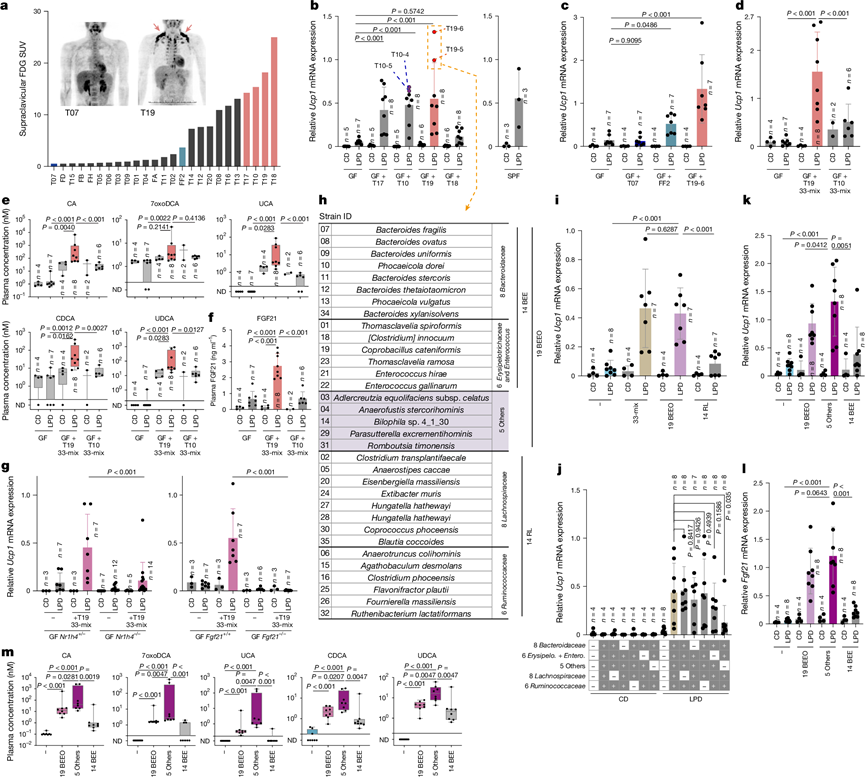
图4. 筛选促进 LPD 介导的褐变的人类来源细菌菌株。
(a) 每位志愿者锁骨上区域的FDG标准化摄取值。(b-d) 通过qPCR测定iWAT中Ucp1的相对表达量。(e-f) 分别通过LC-MS/MS和ELISA定量血浆胆汁酸和FGF21。(g) iWAT中Ucp1的表达。(h) 根据16S rRNA测序确定的33株T19衍生菌株列表。(i-l) iWAT的Ucp1表达和肝脏的Fgf21表达,并以Ppib进行标准化。(m) 小鼠的血浆胆汁酸浓度。
05
微生物氨的产生诱导肝脏FGF21的表达
为了进行额外的菌株剔除实验,他们用5种其他菌株混合物的所有可能的四菌株排列组合接种无菌小鼠,每次剔除一种菌株。值得注意的是,排除Romboutsia timonensis(St.31)阻止了其余四种菌株的定植,并显著降低了米色细胞的诱导(图5a),表明R. timonensis作为支持菌株,促进了其他菌株的定植。排除Adlercreutzia equolifaciens(St.3)、Eubacteriaceae sp.(St.4)或Bilophila sp. 4_1_30(St.14)并未损害其余菌株的定植,但显著影响了它们诱导iWAT褐变的能力(图5a)。相比之下,排除Parasutterella excrementihominis (St.29) 对米色细胞的诱导没有影响(图5a)。因此,虽然Parasutterella菌株并非必需,但其余四个菌株(Adlercreutzia、Eubacteriaceae、Bilophila和Romboutsia;以下统称为“hu4”菌株)对于观察到的 LPD介导的人类相关菌株的褐变效应至关重要。
为了评估hu4菌株组合的代谢效应,研究人员将缺乏棕色化能力的T07号参与者的粪便接种到无菌小鼠体内,并喂以HFD。随后,小鼠接受LPD单独治疗或与两次口服hu4菌株联合治疗。虽然单独使用LPD已有一定的效果,但与hu4菌株联合治疗可显著降低小鼠体重,增加iWAT和BAT中Ucp1和Elovl3的表达,增加肝脏Fgf21的表达,减少iWAT质量,降低血浆胆固醇、甘油三酯和丙氨酸氨基转移酶(ALT)水平,并改善葡萄糖耐量(图5b)。这些效应的发生伴随着肌肉质量的轻微减少和血浆肌酸激酶水平的未升高,表明由特定微生物群成员介导的WAT棕色化增强了LPD喂养的代谢效应,且未造成明显的肌肉损伤。
为了阐明hu4菌株如何促进LPD诱导的脂肪褐变,他们检测了它们的胆汁酸代谢能力。所有四个菌株均表现出胆盐水解酶活性,可在体外将taurocholic acid (tauro-CA)转化为胆酸(CA)(图5c)。此外,支持菌株R. timonensis(St.31)编码7α-羟基类固醇脱氢酶(7αHSDH),并能特异性地将tauro-CA转化为7-oxo-deoxycholic acid (7oxoDCA)(图5c),这可能有助于其他菌株的定植以及iWAT中FXR的激活。
接下来,他们对33株T19衍生菌株进行了测序,并对接种了该菌群并分别喂食对照饮食或LPD的无菌小鼠的盲肠微生物群进行了转录组分析。LPD喂养选择性地上调了19-BEEO和hu4菌株中与氮代谢相关的基因,但未在14-BEE或14-RL菌株中观察到此现象。值得注意的是,在LPD喂养后,Bilophila sp. 4_1_30(St.14)和A. equolifaciens (St.3)中编码亚硝酸盐还原为氨的酶的nrfA基因的表达显著增加(图5d)。与大多数nrfA同源物不同,这些菌株中的nrfA基因独特地含有脂蛋白信号肽 (LSP)(图5e),这一特征在人类微生物组中较为罕见,主要局限于脱硫杆菌门。据预测,这些信号肽可将 NrfA 定位到内膜,从而使其能够进行周质异化硝酸盐还原为铵(图6a)。NrfA 通过其伴侣酶甲酸脱氢酶 (FDH) 以甲酸为电子供体。FDH 的活性位点含有钼蝶呤辅因子,该辅因子可经钨处理抑制。在饮用水中添加钨显著抑制了接种了 19 BEEO 混合物的鼠的 iWAT 褐变和血浆 FGF21 水平,而没有改变菌株移植或血浆胆汁酸水平(图6b-d)。
为了进一步评估 NrfA 在白色脂肪组织褐变中的作用,他们构建了nrfA缺陷型Bilophila sp. 4_1_30 (St.14) 突变株。与nrfA功能正常的(野生型)Bilophila菌株相比,该∆nrfA突变株在体外表现出氨生成缺陷,而后者在蛋白质限制条件下可产生高水平的氨。在体内,将野生型Bilophila菌株与产生胆汁酸的R. timonensis (St.31)菌株共同定植于无菌小鼠体内,可诱导显著的脂肪组织褐变,但其程度远低于完整 hu4 菌株(图6e)。相比之下,尽管ΔnrfA突变株和野生型菌株在肠道中的定植率相当,但与R. timonensis和ΔnrfA Bilophila突变株共定植并未诱导褐变(图6f)。值得注意的是,ΔnrfA Bilophila定植的小鼠肝脏中Fgf21的表达降低(图6g),而肝脏Csad和Cyp39a1的表达以及血浆胆汁酸的诱导则得以保留甚至增强,这表明nrfA介导的氮代谢特异性地促进了Fgf21的诱导,这可能是通过氨的产生实现的。与此一致,LPD喂养增加了SPF B6小鼠和定植野生型Bilophila和R. timonensis的无菌小鼠门静脉中的氨水平,但未增加外周血中的氨水平。相反,在无菌小鼠或定植了∆nrfA菌株的小鼠中未观察到这种增加(图6h)。补充氯化铵可恢复定植了R. timonensis和∆nrfA胆汁酸杆菌的小鼠肝脏中Fgf21以及iWAT中Ucp1和Elovl3的表达(图6i)。此外,用氨刺激人肝细胞类器官可剂量依赖性地诱导FGF21的表达,而不影响胆汁酸生物合成基因(图6j)。这些结果表明,微生物群衍生的氨进入门静脉循环,选择性地诱导肝脏FGF21表达,从而促进WAT褐变。
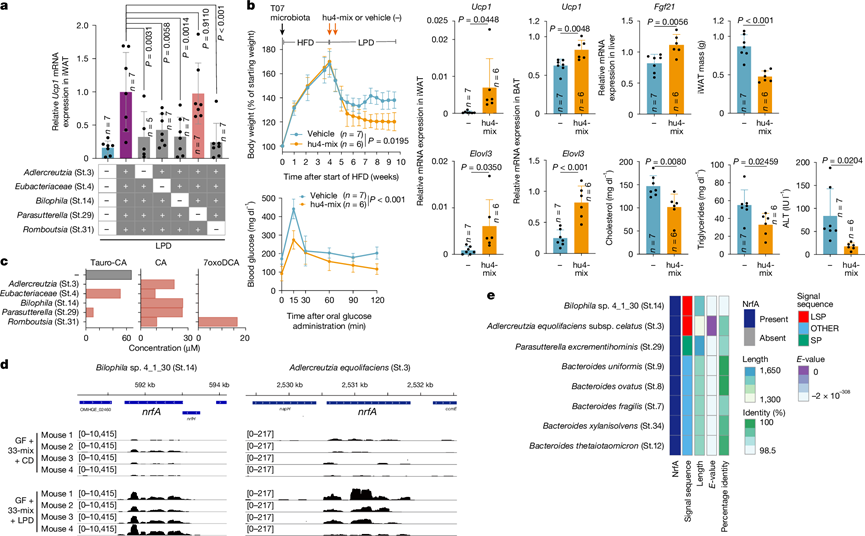
图5. 鉴定出四种促进褐变的人类来源分离物。
(a) 通过qPCR检测iWAT中Ucp1的表达。(b) 评估iWAT和BAT中Ucp1和Elovl3的表达、肝脏Fgf21的表达、iWAT的质量以及血浆胆固醇、甘油三酯和ALT水平。(c) 5种其他菌株混合物中各成员在50 µM tauro-CA孵育后的体外胆汁酸代谢能力。(d) 对定植了T19衍生33菌株混合物并喂食CD或LPD的无菌小鼠盲肠内容物中的细菌RNA进行宏转录组分析。(e) 人源分离株中的NrfA同源物和预测的信号肽。
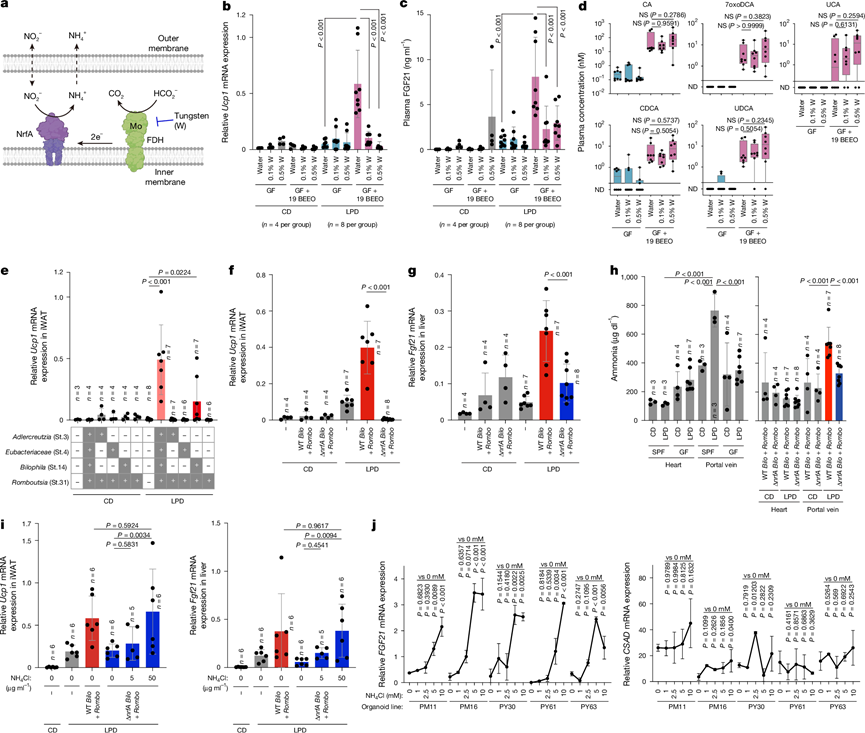
图6. 微生物nrfA依赖的氨产生促进 FGF21 表达。
(a) 示意图展示了NrfA与甲酸脱氢酶(FDH)协同作用还原亚硝酸盐的过程。(b-d) 评估了iWAT中Ucp1的相对表达量、血浆FGF21水平和血浆胆汁酸浓度。(e) 定量分析了iWAT中Ucp1的表达量。(f-g) 采用qPCR检测iWAT Ucp1和肝脏Fgf21的表达。(h) 检测喂食CD或LPD的SPF和GF小鼠以及接种R. timonensis和野生型或ΔnrfA Bilophila的GF小鼠的门静脉血和外周血氨浓度。(i) 评估iWAT Ucp1和肝脏Fgf21的表达。(j) 通过qPCR分析FGF21和CSAD mRNA的表达。
+ + + + + + + + + + +
结 论
本文研究表明,蛋白质的可用性显著调节肠道菌群的功能,并促进WAT的重塑。具体而言,LPD能够显著诱导WAT中棕色化特征基因的表达,其程度与经典刺激(例如冷暴露或β-肾上腺素能受体激活)诱导的表达程度相似。在无菌小鼠中,LPD介导的脂肪褐变显著降低,而通过定植由特定菌株组成的细菌群落可以挽救这一缺陷。这些菌株是从LPD喂养的小鼠或健康志愿者的粪便中分离和筛选出来的,并经18F-氟代脱氧葡萄糖正电子发射断层扫描证实具有棕色或米色脂肪活性。微生物群诱导的脂肪褐变是通过胆汁酸驱动脂肪祖细胞中FXR的激活,以及由编码nrfA的共生菌来源的氨驱动肝细胞中FGF21的表达而介导的。胆汁酸-FXR轴和氨-FGF21轴在促进WAT褐变中均发挥着不可替代的重要作用。这些发现突显了饮食、肠道微生物代谢和脂肪组织重塑之间的机制联系,揭示了宿主对饮食信号做出反应的微生物群依赖性途径。
+ + + + +



