

 English
English文献解读|Cell(64.5):单细胞图谱揭示了高认知功能、痴呆和阿尔茨海默症病理恢复力的相关性
✦ +
+
论文ID
原名:Single-cell atlas reveals correlates of high cognitive function, dementia, and resilience to Alzheimer’s disease pathology
译名:单细胞图谱揭示了高认知功能、痴呆和阿尔茨海默症病理恢复力的相关性
期刊:Cell
影响因子:64.5
发表时间:2023.09.28
DOI号:10.1016/j.cell.2023.08.039
背 景
阿尔茨海默病 (AD) 是最常见的痴呆症。它是一种缓慢进展的神经退行性疾病,从轻度记忆丧失开始,最终导致广泛的执行和认知功能严重受损。AD 的病理特征包括一系列临床表现和细胞病理学,包括 β-淀粉样蛋白积累、配对螺旋丝 (PHF) tau 神经原纤维缠结、神经胶质细胞激活以及神经元和突触损失,但目前对于认知障碍的分子和细胞机制仍然知之甚少。
实验设计
结 果
01

老年人类前额皮质 230 万个细胞核的转录图谱
研究者团队对427 名参与者死后脑组织的前额皮质中分离出的超过 230 万个细胞核进行了单细胞核转录组分析(snRNA-seq)(图1A-B)。427 名研究参与者的样本包括处于不同 AD 进展阶段的个体,从非 AD 和早期 AD(Braak 阶段 0、I 和 II)到中期(阶段 III-IV) AD 晚期(V-VI 阶段)(图 1 A-B)。根据死亡时认知状态的临床共识诊断,146 名研究参与者诊断为无认知障碍,102 名患有轻度认知障碍,144 名患有 AD 痴呆症状(图1B)。在427名研究参与者的队列中,AD和其他病理的分类和定量指标,以及定量临床表型的特征密切反映了整个队列的分布情况(图1C)。最终数据集中总共保留了 2359994 个细胞(图 1 D), 注释了12 个主要组中的 54 种高分辨率细胞类型,其中包括14 种兴奋性神经元亚型(Exc)、25种抑制性神经元亚型(Inh)、少突胶质细胞、少突胶质细胞前体细胞(OPC)、3种星形胶质细胞亚型、5 种免疫细胞类型[小胶质细胞、中枢神经系统 (CNS) 相关巨噬细胞 (CAM) 和 T 细胞]以及多种血管细胞类型[内皮细胞(End)、平滑肌细胞(SMC)、成纤维细胞(Fib)和周细胞(Per)亚群](图 1 F-1J)。
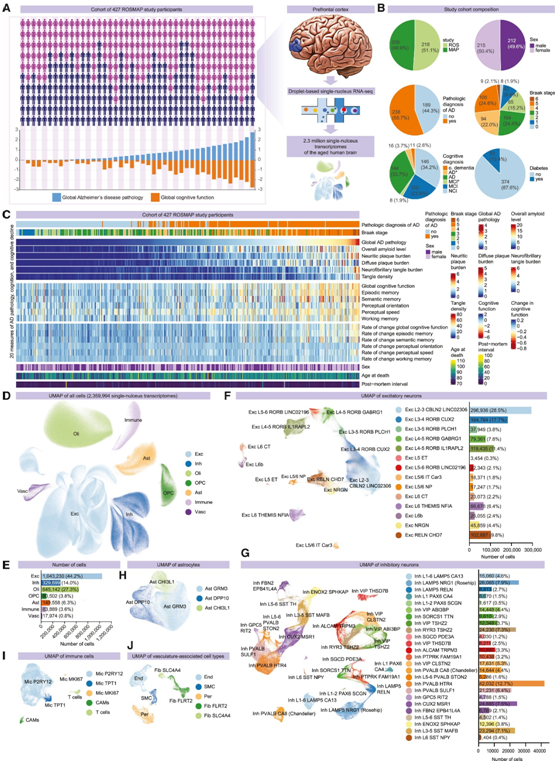
图1. 对 427 名 ROSMAP 研究参与者的前额皮质组织进行单核 RNA-seq 分析。
(A) 队列和 snRNA-seq 分析摘要。(B) 研究队列组成。(C)队列元数据概述。(D) 7 个主要细胞类别的 230 万个细胞的联合 UMAP,包括兴奋性神经元 (Exc)、抑制性神经元 (Inh)、少突胶质细胞 (Oli)、少突胶质细胞前体细胞 (OPC)、星形胶质细胞 (Ast)、免疫细胞 (Immune)、以及血管和上皮细胞 (Vasc)。(E) 条形图显示每个主要细胞类别的细胞数量。(F–J) 兴奋性神经元亚型 (F)、抑制性神经元亚型 (G)、星形胶质细胞亚型 (H)、免疫细胞亚型 (I) 以及血管和上皮细胞亚型 (J) 的 UMAP。条形图显示兴奋性 (F) 和抑制性 (G) 神经元亚型的细胞数量。
02
基因表达的系统差异分析
为了确定不同的临床和生活方式变量如何影响个体细胞类型,他们分析了 54 种细胞类型中每个变量的差异表达基因(DEG) (图 2 A)。考虑了 AD 病理学、路易体和 TDP-43 病理学、血管病理学、医疗状况以及认知、身体和社会生活方式变量的多种因素(图 2 A)。
他们随后将分析重点放在 AD 病理变量上,因为大量基因与其相关(图 2 A)。在几乎所有细胞类型中均发现了与 AD 病理学显著相关的DEG(图2A)。当比较与不同类型 AD 病理相关的 DEG 时,他们观察到与整体 AD 病理、神经炎斑块负荷、神经原纤维缠结负荷和许多神经元亚型以及星形胶质细胞和少突胶质细胞中缠结密度呈正相关或负相关的基因显著重叠(图 2 B-F)。他们还发现不同兴奋性神经元亚型之间与 AD 病理相关的 DEG 存在显著重叠(图 2G-H)。
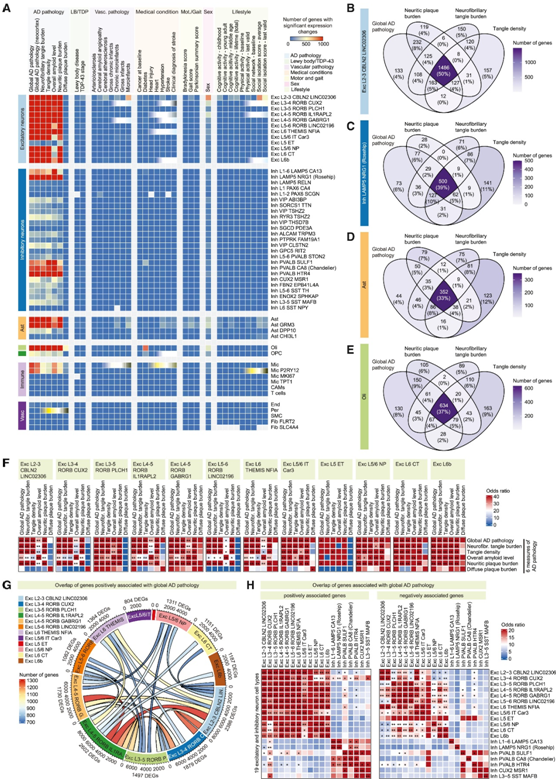
图2. 兴奋性神经元亚型共有与 AD 病理相关的基因表达变化。
(A) 热图显示 54 个细胞类型(行)和 36 个变量(列)中显著差异表达的基因的数量。(B–E) 维恩图显示兴奋性神经元亚型 Exc L2–3 CBLN2 LINC02306 (B) 和抑制性神经元亚型 Inh 中与整体 AD 病理学、神经炎斑块负荷、NFT 负荷和缠结密度呈显著正相关的基因数量。(F) 热图显示与所示变量显著正相关的基因重叠。(G) Circos 图显示兴奋性神经元亚型之间与整体 AD 病理呈正相关的基因重叠。(H) 热图显示与指示的兴奋性神经元亚型之间的整体 AD 病理学显著正相关(左图)或负相关(右图)的基因重叠。
03
AD 病理相关变化在兴奋性神经元亚型中保守
为了鉴定在多个兴奋性神经元亚型中反复改变的基因、通路和生物过程,他们在至少三种不同的兴奋性神经元亚型中定义了与整体 AD 病理学显著相关的基因的共识特征。与整体 AD 病理学正相关的共识特征富含参与神经元投射形态发生、mRNA 代谢过程(包括异质核核糖核蛋白HNRNPA2B1和HNRNPH3)、染色质组织和突触信号传导的基因(图 3A-C)。 与整体 AD 病理学负相关的共识特征涉及 tRNA 代谢过程和脂质代谢的基因(图 3 B)。在前额叶皮质(PFC) 中也观察到的与突触信号、RNA剪接、脂质代谢和线粒体功能相关的基因表达差异(图3D)。
多种细胞类型的 AD 病理学多种指标呈负相关的基因包括脂质代谢基因 BDH1、GPCPD1、MECR 和 TM7SF2,这些基因参与胆固醇生物合成(图 3 E)。虽然共识特征是基于兴奋性神经元的基因表达差异,但星形胶质细胞中脂质代谢和胆固醇生物合成基因的失调也很明显(图3F)。脂质代谢的改变伴随着编码线粒体电子传递链成分的基因的广泛变化,患有高水平 AD 病理学的个体表现出编码线粒体膜间空间桥接复合物 (MIB) 成分的几个基因的表达减少,MIB 是一种复合物这对于维持线粒体内膜结构至关重要(图 3G-H)。

图3. 与AD病理相关的基因表达变化。
(A-B)在与整体 AD 病理显著正相关或负相关的基因中富集的通路。(C,E,G) 基因表达水平(行)与所示细胞类型的整体 AD 病理学之间的关联(列)。(D) 差异表达基因 (DGE) 的结果分析。(F) 星形胶质细胞中胆固醇生物合成基因的表达水平与整体 AD 病理学之间的负相关性。(H) MIB 复合体基因的表达水平与第 4/5 层兴奋性神经元 (Exc L4-5 RORB IL1RAPL2) 中的整体 AD 病理学之间的关联。
04
AD 中粘连蛋白复合物表达和 DNA 损伤反应的协调增加
为了评估 AD 是否影响 MIB 复合物之外的其他蛋白质复合物,他们计算了 CORUM 2417 种人类蛋白质复合物的模块分数,并测试了它们与整体 AD 病理学的关联。粘连蛋白是一种调节姐妹染色单体凝聚、同源重组和 DNA 循环的蛋白质复合物,在患有高水平 AD 病理的个体 PFC 的兴奋性神经元亚型中,粘连蛋白的升高最为一致且显著,也与 PFC 少突胶质细胞中的整体 AD 病理呈正相关(图 4 A)。在其他大脑区域的兴奋性神经元亚型子集中也观察到粘连蛋白复合物表达增加(图4B-C)。
为了探索 AD 中粘连蛋白表达增加的背景和可能的功能,他们计算了粘连蛋白复合物与兴奋性神经元细胞类型的所有表达基因之间的相关性。有趣的是,许多与粘连蛋白复合物呈正相关的基因,包括染色质调节因子 ATRX、ZBTB1、SIRT1、CHD1、BPTF 和 ZBTB7A,在细胞对 DNA 损伤的反应中发挥着关键作用(图 4 D)。其他与内聚蛋白复合物正相关的蛋白汇集到DNA损伤位点(NIPBL),调节碱基切除修复(USP47),促进DNA损伤恢复(BAZ1B),调节DNA损伤反应(CDKN2AIP),并在DNA损伤修复(MACROD1)中发挥关键作用(图4D)。在少突胶质细胞中也观察到 AD 中粘连蛋白复合物和 DNA 损伤反应基因表达的协调增加(图 4 E-G)。这些结果表明,AD 相关的粘连蛋白复合物表达增加伴随着大量染色质调节因子和 DNA 损伤反应因子的表达升高,这些调节因子和 DNA 损伤反应因子促进基因组完整性。

图4. AD中粘连蛋白复合物表达和DNA损伤反应的协调升高。
(A) 14 种蛋白质复合物的表达(行)与所示细胞类型(列)和大脑区域的整体 AD 病理学之间的关联。(B-C) 基于来自 638 名 ROSMAP 研究参与者背外侧前额皮质的bulk RNA 测序数据和定量蛋白质组学,确认 AD 中粘连蛋白复合物表达增加。(D) 左:粘连蛋白复合物和共有特征基因之间的相关性(关联分数)与兴奋性神经元亚型的整体 AD 病理呈正相关。右 :基于左图所示兴奋性神经元关联评分的皮尔逊相关性的基因 k 均值聚类。(E) 与整体 AD 病理学正相关的基因(在少突胶质细胞中)和少突胶质细胞中所示的蛋白质复合物(列)之间的相关性(关联分数)。(F) 左:少突胶质细胞中所示的蛋白质复合物(列)和基因(行)表达水平之间的相关性。中:总体 AD 病理学与所示细胞类型(列)中与粘连蛋白复合物(行)共同调节的基因表达水平之间的关联。右:少突胶质细胞中与粘连蛋白复合物共同调节的基因表达水平(行)与所示变量(列)之间的关联。(G) 四种神经胶质细胞类型和人脑四个不同区域的粘连蛋白-SA1 复合物的表达水平与所示基因(行)之间的相关性。
05
AD进展多个阶段基因表达的时间变化
为了加深对与 AD 进展相关的分子变化的分析,他们根据AD 病理变量将整个队列分为三组,并确定 DEG,将第二组与第一组 AD 病理水平最低的个体进行比较,并确定 DEG。第三组的 AD 病理水平高于第二组(图 5 A)。与不同类型 AD 病理学相关的基因表达变化的趋同性,以及在疾病进展的早期和晚期阶段兴奋性神经元亚型之间的 DEG 显著重叠。为了在分子水平上检查 AD 进展,他们将整个队列中与 AD 病理相关的基因,与根据疾病阶段特异性差异表达分析确定的 DEG 进行交叉分析(图 5 A)。在队列中发现的许多 DEG 在疾病早期或晚期阶段也显示出显著变化。在多种细胞类型中,与 AD 病理学正相关的早期变化富含参与突触组织、细胞投射组织调节、染色质组织和 DNA 损伤反应的基因(图 5B)。星形胶质细胞、兴奋性和抑制性神经元、少突胶质细胞和 OPC 中与 AD 病理负相关的早期变化中,参与自噬的基因显著富集(图 5 C)。与 AD 病理学呈正相关的晚期变化,涉及神经元中 mRNA 代谢过程、染色质组织、DNA 损伤反应、细胞投射组织、突触信号传导和蛋白质分解代谢过程的基因富集(图 5B)。在疾病进展后期与AD病理负相关的基因在星形胶质细胞、兴奋性和抑制性神经元以及少突胶质细胞中的脂质代谢基因富集(图5C)。他们还研究了 PFC 和颞中回(MTG) 中多种细胞类型中显著的基因子集,发现其中许多与疾病进展后期多种神经元和神经胶质细胞类型的整体 AD 病理学显著相关(图 5 E)。与这些观察结果一致,他们发现粘连蛋白复合物的表达往往随着 AD 的进展而增加,并且在总体 AD 病理水平最高的个体中显著升高(图 5D)。最后,他们通过比较 PFC 和 MTG 数据集之间的基因表达变化来探索 AD 的疾病进展。早期的变化在数据集之间有相当大的重叠(图 5G),有趣的是,在 PFC 疾病进展后期显著的相当大一部分基因在 MTG 早期也发生了显著改变(图 5E -F)。

图5. 阿尔茨海默病进展多个阶段基因表达的时间变化。
(A)差异表达分析方法的示意图。(B-C) GO分析。(D) 箱线图显示指定细胞类型中每个个体的平均粘连蛋白复合物表达(模块得分)的分布。(E-F)分别在前额叶皮层和中颞叶皮层中涉及生物过程的选定基因的表达水平(行)与整体AD病理学和连续伪进展评分之间的关联。(G) PFC 中疾病进展的不同阶段正相关的基因与 MTG 中疾病进展的不同阶段正相关的基因的重叠部分。
06
老年人大脑认知障碍的分子相关性
为了确定老年人大脑认知障碍的分子相关性,他们分析了个体细胞类型中的差异基因表达,包括总体认知功能的汇总测量以及个体认知领域的测量和个体特定的变化率。随着时间的推移这些变量。在几乎所有细胞类型中都发现了与认知显著相关的 DEG(图 6A)。与不同认知变量相关的基因在大多数兴奋性神经元细胞类型、抑制性神经元亚型的子集、星形胶质细胞和少突胶质细胞中显著重叠。这些基因还与 AD 病理相关基因显著重叠,表明在特定细胞类型内,与认知障碍相关的基因表达变化密切反映了 AD 病理相关变化(图 6 B)。
负相关基因在突触后的表达最为显著,而与整体认知功能正相关的基因,包括CACNG3、PAK1、NPTX2、RPH3A、SVOP和BDNF,都映射到突触前和突触后的区室(图6C-E)。使用RNA原位杂交(图6F)和免疫组织化学(图6G)证实NPTX2在认知障碍个体中的表达显著降低。RT-PCR实验也证实其他与整体认知功能正相关的突触基因包括NUMB和VGF(图6D-E)。最后,他们观察到参与成纤维细胞生长因子 (FGF) 信号传导的几个基因与多种兴奋性神经元亚型的多种认知功能呈正相关(图6D-E)。总之,这些结果强调了与老年人大脑中的高认知功能和痴呆症相关的常见和细胞类型特异性基因、通路和细胞成分。

图6. 老年人大脑认知障碍的分子相关性。
(A) 热图显示显著差异表达基因的数量。(B) 热图显示与 L4-5 兴奋性神经元 (Exc L4-5 RORB IL1RAPL2) 中指示的变量显著相关的基因的重叠。(C) 旭日图显示与整体认知功能(基因数量)正相关的基因突触定位。(D) 突触基因表达水平(行)与所示细胞类型(列)的整体认知功能之间的关联。(E) L4-5 兴奋性神经元 (Exc L4-5 RORB IL1RAPL2) 中突触基因(行)和所示变量(列)之间的关联。(F) 使用 RNAscope 探针对无认知障碍个体(左)和有认知障碍个体(右)的 PFC 灰质中的 NPTX2(红色)进行 RNA 原位杂交。(G)免疫组织化学分析.
07
随着 AD 病理积累,神经元和神经胶质的组成发生变化
细胞死亡是细胞功能障碍的最终形式,也是包括阿尔茨海默氏病在内的神经退行性疾病的标志。他们研究了 AD 病理进展如何影响细胞类型组成,以确定特别脆弱的细胞类型。首先评估了主要细胞类型的比例是否随着 AD 病理的进展而变化,发现大多数主要细胞类型的相对丰度没有显著变化(图 7A -B)。一个例外是小胶质细胞群,在总体 AD 病理、神经炎斑块负荷和总体淀粉样蛋白水平最高的个体中,其相对丰度显著增加(图 7 A-B)。相比之下,随着 AD 病理学的进展,兴奋性神经元的相对丰度呈微妙下降的趋势。
接下来他们在高分辨率细胞类型水平上评估了细胞类型组成随 AD 病理进展的变化,确定了 SST 亚类的三种选择性易受攻击的抑制性神经元亚型(Inh CUX2 MSR1、Inh ENOX2 SPHKAP 和 Inh L3-5 SST MAFB),这些亚型在总体 AD 病理、神经炎斑块负担水平最低、弥漫性斑块负荷、总淀粉样蛋白水平、神经原纤维缠结(NFT)以及没有 AD 病理诊断的个体的个体子集中显著过高(图 7C)。有趣的是,抑制神经元的统一流形近似和投影(UMAP)表示显示,三种易感的SST亚型彼此相邻聚集,表明这些亚型具有高度的转录组相似性,可能解释了AD病理的共同易感性(图7D)。与过度代表性分析的结果一致,在AD病理、NFT负担和缠结密度最高的个体子集中,SST抑制神经元的整个亚类的相对丰度以及选择性易感SST神经元组的个体亚型的相对丰度显著降低,这一发现通过准二项回归模型得到了进一步证实(图7E-F)。
接下来,他们在高分辨率细胞类型水平上探讨了与认知衰退相关的细胞类型组成的差异,确定了多种抑制性神经元亚型,包括脆弱的 SST 神经元,这些亚型在具有最高水平的整体认知功能、情景记忆和语义记忆的个体子集中的水平明显过高(图 7 G)。在具有高认知功能的个体中过多代表的其他抑制神经元亚型包括RELN阳性的LAMP5亚类抑制神经元(Inh LAMP5 RELN),以PTPRK和FAM19A1表达为标志的亚型(Inh PTPRK FAM19A1),以及以SORCS1和TTN表达为标志的亚型(Inh SORCS1 TTN)(图7G)。抑制神经元的UMAP表征显示,三种抑制神经元亚型Inh LAMP5 RELN, Inh PTPRK FAM19A1和Inh SORCS1 TTN(以下简称LAMP5 RELN组)彼此相邻聚集,表明这些亚型之间具有高度的转录组相似性(图7D)。
在整体认知功能、情景记忆、知觉定向和语义记忆测试中得分较低的个体中,整个 LAMP5 RELN 组抑制性神经元和整个 SST 抑制性神经元亚类的相对丰度显著降低(图 7 H-J)。
在病理诊断为 AD 的个体中,与诊断为 AD 痴呆的个体相比,无认知障碍的个体中抑制性神经元的相对丰度显著更高(图 7 K-L),无认知障碍的个体中易受影响的 SST 神经元的相对丰度显著更高(图 7 M)。关注抑制性神经元的LAMP5 RELN组,他们发现整个组及其两个亚型(Inh LAMP5 RELN和Inh PTPRK FAM19A1)的相对丰度在整体AD病理、神经斑块负担、NFT具有高CR的个体中显著更高(图7N)。此外,在病理诊断为AD的个体中,与诊断为AD痴呆的个体相比,无认知障碍的个体中LAMP5 RELN组个体亚型的相对丰度显著更高(图7O)。
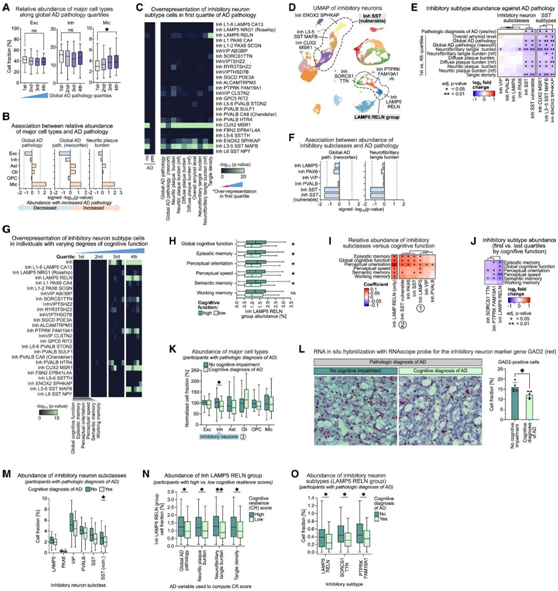
图7. 与 AD 病理学和认知障碍相关的细胞类型组成变化。
(A) 箱线图显示了疾病进展过程中主要细胞类型的相对丰度。(B) 主要细胞类型的相对丰度与 AD 病理学测量值之间的关联(关联评分)。(C-G) 从具有不同程度 (C) AD 病理学或 (G) 认知功能的个体分离的细胞的每种抑制性神经元亚型中的过度表达。(D) 按亚型着色的抑制性神经元的 UMAP。(E) 比较具有高和低 AD 病理学的个体的抑制性神经元亚类和 SST 抑制性神经元亚型的相对丰度。(F) 抑制性神经元亚类的相对丰度与 AD 病理学测量之间的关联。(H) 比较具有高认知功能和低认知功能的个体的 LAMP5 RELN 抑制性神经元组的相对丰度。(I) 抑制性神经元亚类的相对丰度与认知功能测量之间的关联。(J) 比较具有高认知功能和低认知功能的个体的抑制性神经元亚型(LAMP5 RELN 组)的相对丰度。(K,M,O) 具有 AD 病理诊断的研究参与者中主要细胞类型、抑制性神经元亚类或 抑制性神经元亚型(LAMP5 RELN 组)的相对丰度。(L) 使用 RNAscope 探针进行 RNA原位杂交。(N) LAMP5 RELN 组抑制性神经元的相对丰度。
+ + + + + + + + + + +
结 论
本项研究采用单细胞转录组学技术分析了老年人前额叶皮层组织,覆盖了427名患有不同程度AD病理和认知障碍的人死后大脑样本中的230万个细胞,确定了AD病理相关的改变在兴奋性神经元亚型之间共有,揭示了兴奋性神经元和少突胶质细胞中内聚蛋白复合物和DNA损伤反应因子的协调增加,并揭示了与高认知功能、痴呆和AD病理恢复相关的基因和途。此外,还鉴定了选择性易损的生长抑素抑制性神经元亚型,发现两组不同的抑制性神经元在晚年保持高认知功能的个体中富集程度更高,并揭示了抑制性神经元与AD病理恢复能力之间的联系。
+ + + + +



