

 English
English文献解读|Immunity(32.4):对 BCG 疫苗接种的先天和适应性反应的多组学分析揭示了可预测训练后免疫力的表观遗传细胞状态
✦ +
+
论文ID
原名:Multi-omics analysis of innate and adaptive responses to BCG vaccination reveals epigenetic cell states that predict trained immunity
译名:对 BCG 疫苗接种的先天和适应性反应的多组学分析揭示了可预测训练后免疫力的表观遗传细胞状态
期刊:Immunity
影响因子:32.4
发表时间:2024.01.09
DOI号:10.1016/j.immuni.2023.12.005
背 景
卡介苗(BCG)是一种 100 多年前开发的减毒活结核病疫苗。它是世界上使用最广泛的疫苗之一。单剂量给药,它可以增强抗分枝杆菌宿主反应,从而预防结核病,尽管不同个体和地理区域的功效不同。BCG还可以增强先天免疫系统的反应能力,并可能提高发展中国家儿童的总体生存率。在接种卡介苗后,无论是特异性适应性免疫反应还是经过训练的免疫反应,都观察到个体间的高度变异性。这种变异性在很大程度上仍然无法解释。
实验设计
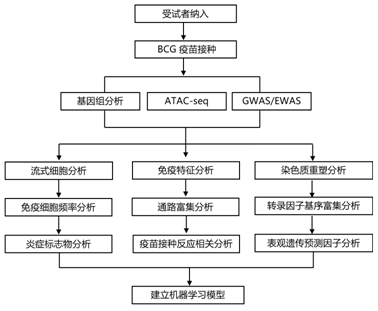
结 果
01

健康个体基线的个人免疫特征
为了评估基线和体内刺激后免疫功能的个体间差异,研究者团队在一项介入性临床研究中让 323 名健康志愿者接受了 BCG 疫苗接种(图 1 A)。他们在疫苗接种前(第 0 天)、预计会出现主动免疫反应的疫苗接种后早期阶段(第 14 天)以及在急性反应消退但随后的时间点收集了血液样本和纯化的外周血单核细胞 (PBMC)。
他们通过流式细胞分析免疫细胞频率、测量 73 种循环炎症标志物的血浆浓度以及评估对四种微生物刺激(白色念珠菌[ C. albicans ]、大肠杆菌脂多糖 [LPS]、金黄色葡萄球菌[ S. aureus ] 和结核分枝杆菌[ M. tuberculosis ]),并进一步建立了个人基因型(使用单核苷酸多态性 (SNP) 微阵列)和染色质可及性图谱(使用 ATAC-seq 测定转座酶可及染色)(图 1 A)。对于免疫细胞频率(图1B),较高的实际年龄与较低的CD8+ T细胞频率和较高的非经典单核细胞(CD14++CD16+)和自然杀伤(NK)细胞频率相关,与先前的结果一致,他们还观察到性别特异性差异,最明显的是男性NK细胞的频率更高。循环炎症标志物浓度反映了采集样本时的免疫状态,随着年龄的增长而增加(“炎症”)(图1C)。此外,他们在男性和报告使用口服避孕药的女性中检测到炎症标志物浓度升高。相比之下,最近饮酒(过去24小时)、BMI和昼夜节律通常与炎症标志物水平升高或降低无关,尽管有一些统计学上显著的关联。
细胞因子生产能力(图1D),评估细胞应对免疫挑战的潜力,与pbmc中单核细胞的百分比相关。在对免疫细胞频率进行统计校正后,较高的实足年龄与白色念珠菌诱导的CXCL9和CXCL10水平下降以及LPS和结核分枝杆菌应答中IL-6产生减少有关,这与免疫衰老的概念一致。
对染色质可及性与年龄相关的差异进行富集分析,突出显示了参与 IL-10 产生的基因,而昼夜节律相关区域则与免疫调节和细胞因子反应相关的途径相关(图 1E)。对于细胞因子的产生能力,他们的 GWAS 分析检测到了 6 个显著相关的基因组位点(图 1 F),其中几个与免疫基因相关。他们还发现了TLR10-TLR1-TLR6位点的一个变体(rs5743618)与影响细胞因子产生和抗炎细胞因子IL-10浓度之间的关联,提示存在负反馈机制。通路分析发现,与“革兰氏阴性细菌的防御反应”、“细胞因子分 泌的积极调节”和“糖酵解过程”相关的基因富集(图1G)。平行EWAS分析发现CCL3L1启动子染色质可及性与细胞因子生产能力之间存在关联,包括与白色假丝酵母菌诱导的CCL4生产呈正相关(图1H)。CCL3L1编码一种促炎细胞因子,吸引单核细胞和淋巴细胞到炎症和感染部位。他们还发现结核分枝杆菌诱导的IL-12产生与位于PGS1基因内的染色质可及性之间存在关联,该基因编码参与脂质代谢的磷脂酰甘油磷酸合成酶。该区域位于SOCS3基因上游约50kb处,该基因抑制细胞因子信号转导和IL-12信号传导。通路分析表明,具有较高细胞因子生产能力的个体在与中性粒细胞和粒细胞活化和募集相关基因相关的区域具有较高的染色质可及性(图1L)。计算细胞因子生产能力的方差百分比,发现免疫细胞频率和遗传有关,宿主因素和表观遗传变异的作用较小(图1J)。
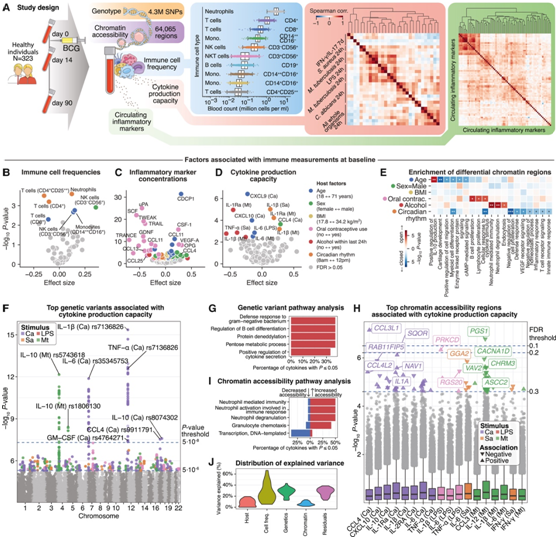
图1. 多组学分析描绘了基线免疫和对 BCG 疫苗接种反应的群体差异特征。
(A) 300BCG 队列和 BCG 疫苗接种时间过程的概述。(B–D) 基线时宿主因素与个人免疫特征之间关系的火山图。(E) 具有宿主因子相关染色质可及性的基因组区域的通路富集。(F) 曼哈顿图显示遗传变异与细胞因子产生能力显著相关。(G) 与细胞因子产生能力相关的遗传变异的途径富集。(H) 箱线图显示具有与细胞因子产生能力相关的染色质可及性的基因组区域。(I) 具有与细胞因子产生能力相关的染色质可及性的基因组区域的通路富集。(J) 由宿主因素、免疫细胞频率、遗传变异、染色质可及性概况和残留的无法解释的方差解释的细胞因子产生能力的估计方差百分比范围。
02
卡介苗接种后的个人免疫特征
卡介苗疫苗接种不仅能诱导结核病特异性的适应性免疫,还能诱导先天免疫记忆(经过“训练”的免疫),从而提高对各种病原体的免疫反应。他们研究了它对免疫细胞频率、循环炎症标志物、细胞因子产生能力和染色质可及性特征的影响(图 2A)。
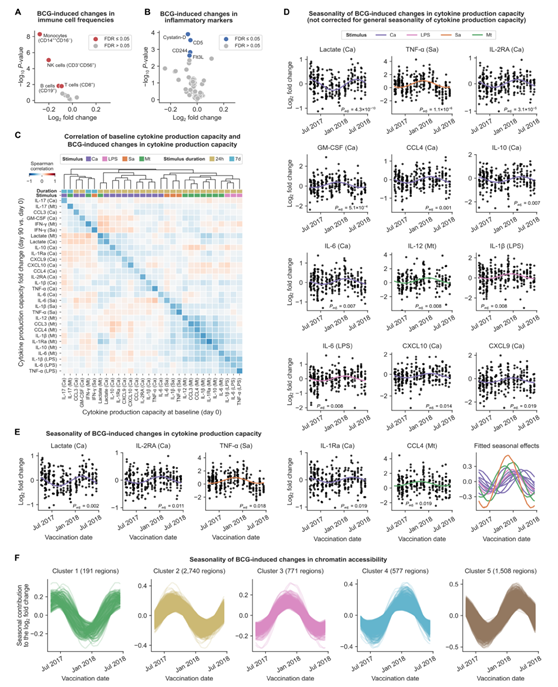
在接种 BCG 疫苗后第 90 天,经典单核细胞 (CD14+ +CD16 - )、NK 细胞、B 细胞和 CD8 + T 细胞的免疫细胞频率略有下降(图 S2 A)。在循环炎症标志物中,半胱氨酸蛋白酶抑制剂 D、CD5、CD244 和 Flt3L 的浓度在 BCG 疫苗接种后 90 天下降(图 S2 B)。Flt3L是一种造血生长因子,参与免疫细胞发育,它的减少与循环免疫细胞频率的降低相一致。比较第 90 天和第 0 天之间的细胞因子产生能力(图2B),他们观察到用结核分枝杆菌刺激 PBMC 后 IFN-γ 产生的预期增加,这与卡介苗接种后病原体特异性适应性免疫记忆一致。然而,他们还观察到与结核分枝杆菌无关的变化,这可能是卡介苗疫苗接种诱导的非特异性先天免疫记忆的结果。
他们观察到个体之间的适应性和先天免疫记忆存在显著差异,对结核分枝杆菌的病原体特异性(适应性)反应比对其他微生物刺激的非特异性(先天)反应的群体差异更明显(图2C)。比较第 90 天和第 0 天之间刺激和细胞因子的差异倍数(图 2D),反应曲线主要按刺激而不是细胞因子聚集。例如,白色念珠菌刺激显示出与LPS刺激截然不同的细胞因子。他们观察到BCG诱导的适应性免疫记忆(通过对结核分枝杆菌的IFN-γ和IL-17的产生来测量)和先天免疫记忆(通过对异源刺激的各种细胞因子来测量)之间存在微弱且通常为负相关(图2D)。对于结核分枝杆菌的刺激,先天免疫细胞因子产生的变化似乎与患者特异性先天免疫记忆的相关性比适应性免疫记忆更强(图2D)。此外,结核分枝杆菌引起的乳酸释放增加(表明糖酵解率增加)与先天免疫记忆标志物呈正相关(图2D),与适应性免疫记忆标志物负相关(图2D)。
疫苗接种前(第 0 天)的免疫细胞频率确实与 BCG 诱导的细胞因子产生能力的变化相关(图 2 E)。非经典单核细胞(CD14 + + CD16 +)的基线频率与先天免疫记忆负相关,而经典单核细胞(CD14 + + CD16 -)和非经典单核细胞的基线频率与适应性免疫反应正相关。基线时的细胞因子产生能力与 BCG 诱导的先天性和适应性免疫记忆呈负相关(图 S2C)。BCG 诱导的细胞因子产生能力与任何研究的宿主因素均无关(图 2F)。相反,他们发现与每年季节变化存在显著关联,通常与冬季 BCG 疫苗接种引起的更强的先天免疫记忆反应有关(图 S2 D)。对于金黄色葡萄球菌诱导的TNF-α和白色念珠菌诱导的IL-2RA和乳酸生成,即使他们对细胞因子产生能力的基线季节性进行统计校正,卡介苗效应与季节性之间的关联仍然存在(图2G,图S2E)。
他们确定了5787个基因组区域,其中bcg诱导的染色质可及性变化遵循季节性模式,产生了五个不同的聚类(图2H,图S2F)。其中3个聚类富集了特定的生物学功能,包括与RNA剪接和加工相关的通路(bcg诱导的变化高峰区域在10月至1月)和与中性粒细胞介导的免疫和自噬相关的通路(bcg诱导的变化高峰区域在7月至9月)(图2I)。
在本项研究的数据集中,没有观察到疤痕大小和bcg诱导的细胞因子生产能力变化之间的显著关联。相反,他们发现疤痕大小与IFN-γ的产生之间存在强烈的正相关,这是对结核分枝杆菌刺激的反应(图2J)。这些结果为疤痕发展与低死亡率之间的关联提供了另一种解释:疤痕大小可能反映NK细胞和T细胞免疫应答在基线时的强度,而不是反映卡介苗接种诱导的免疫记忆的强度,并且具有强基线应答的个体可能更好地保护免受卡介苗接种之外的感染。
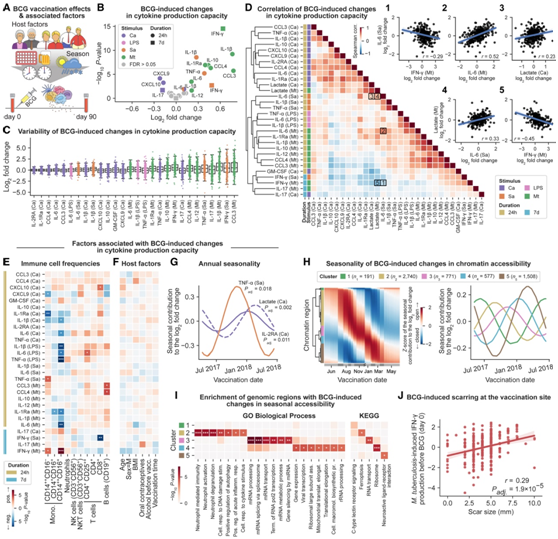
图2. BCG 疫苗接种会引起细胞因子产生能力的变化,这种变化是季节性的,并且与基线时的免疫功能相关。
(A) 研究卡介苗疫苗接种对细胞因子产生能力影响的研究设计示意图。(B) 火山图显示接种疫苗 90 天后,BCG 诱导的细胞因子产生能力变化,以响应白色念珠菌(Ca)、LPS、金黄色葡萄球菌(Sa) 和结核分枝杆菌(Mt) 刺激。(C) 箱线图显示 BCG 诱导的细胞因子产生能力的变异性。(D) 聚类热图(左)显示 BCG 诱导的细胞因子和乳酸生产能力变化的成对相关性。对于选定的细胞因子和刺激组合,BCG 引起的个体变化(每个点是一个个体)的散点图(右)。(E-F) 基线时免疫细胞频率 (E) 和宿主因子 (F) 与 BCG 诱导的细胞因子产生能力变化的关联。 (G) 季节性与 BCG 诱导的金黄色葡萄球菌诱导的 TNF-α 变化和白色念珠菌诱导的 IL-2Ra 和乳酸产生变化之间的关联。(H) 对 BCG 引起的染色质可及性变化的季节性贡献的层次聚类(左)和五个已识别聚类中每个聚类的平均季节性趋势(右)。(I) BCG 诱导的季节性染色质可及性变化导致基因组区域的通路富集。(J) BCG 诱导的疤痕大小与结核分枝杆菌诱导的 IFN-γ 基线产生的关联。
图S2. bcg诱导的免疫反应和季节变化。
(A-B)火山图显示接种后90天bcg诱导的免疫细胞频率和炎症标志物浓度的变化。(C)基线时(第0天)细胞因子生产能力的层次聚类和接种后90天bcg诱导的细胞因子生产能力变化。(D) bcg诱导细胞因子生产能力变化的年度季节性变异性。(E) 校正了细胞因子生产能力的一般季节性后,bcg诱导的细胞因子生产能力变化的年季节性变化。(F) bcg诱导的染色质可及性变化的年季节性变化,分为五个聚类。
03
BCG 疫苗接种反应的遗传和表观遗传关联
利用基线的基因型数据和染色质可及性概况,他们对 BCG 诱导的细胞因子生产能力的变化进行了并行 GWAS 和 EWAS 分析(图 3 A)。rs9595087 变异与 BCG 诱导的 LPS 诱导的 TNF-α 产生变化密切相关。这种遗传变异与TSC22D1的表达水平有关,TSC22D1是一种编码tgf-β应答转录因子的基因(图3B-C)。他们进一步观察到rs57438517变异与bcg诱导的结核分枝杆菌诱导的IL-1Ra产生的变化之间的关联。这种变异与NIPA1的表达有关,NIPA1编码一种镁转运蛋白,在结核分枝杆菌感染时受到抑制。
他们对外源刺激下bcg诱导的先天细胞因子产生扩增相关的遗传变异(表明免疫训练)进行了通路分析,并观察到与细胞代谢相关的生物过程的富集(图3D)。他们还发现了4个基因组区域,其染色质可及性在基线时与异源刺激下bcg诱导的先天细胞因子产生的变化相关(图3E-F)。bcg诱导的白色念珠菌诱导的GM-CSF产生的变化与UCP3基因有关,UCP3基因在氧化代谢中起关键作用。这与先前的研究一致,表明氧化磷酸化对bcg诱导的训练免疫至关重要。通路富集分析确定了免疫和代谢过程,如“中性粒细胞介导的免疫”、“炎症反应”、“戊糖和葡萄糖酸盐相互转化”和“糖胺聚糖生物合成”(图3G)。具体而言,基线可及性与先天免疫记忆负相关的区域在促炎途径中富集,表明在接种疫苗前这些基因上具有更多抑制染色质的个体在接种疫苗后表现出更强的训练免疫反应增强。
通过将观察到的bcg诱导的细胞因子生产能力的群体差异划分为可归因于不同因素的部分(图3H),他们发现宿主因素和昼夜节律的作用可以忽略不计;季节性(接种日期)和基线时的免疫细胞频率与一些刺激因子-细胞因子组合相关,基线时的基因型和染色质可及性谱具有更强的影响,基因型比染色质可及性解释的差异更多。
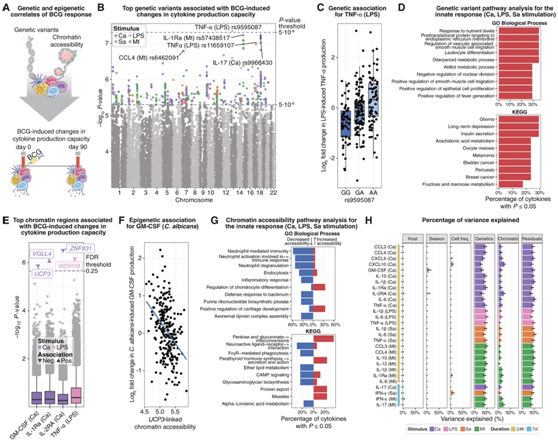
图3. 基线基因型和染色质可及性概况与 BCG 疫苗接种反应相关。
(A) BCG 诱导的细胞因子产生能力变化与遗传变异 (GWAS) 和染色质可及性谱 (EWAS) 的匹配关联分析示意图。(B) 曼哈顿图显示遗传变异与 BCG 诱导的指定细胞因子和刺激的变化显著相关。(C) 箱线图显示 SNP rs9595087 的 LPS 刺激的 TNF-α 产生中 BCG 诱导的变化。(D) 与 BCG 诱导的先天细胞因子产生能力变化相关的遗传变异的通途径富集。 (D) 与 BCG 诱导的先天细胞因子产生能力变化相关的遗传变异的途径富集。(F) 散点图,显示UPC3连锁基因组区域中的染色质可及性与 BCG 诱导的白色念珠菌诱导的 GM-CSF 产生变化之间的关联。(G) 基因组区域的途径富集,其染色质可及性与 BCG 诱导的先天细胞因子产生能力的变化相关。(H)由宿主因素、季节、免疫细胞频率、遗传变异、染色质可及性谱和BCG诱导的细胞因子生产能力变化的估计方差百分比。
04
应答者与无应答者中 BCG 诱导的染色质重塑
与静态基因型相反,染色质可及性谱随着卡介苗接种而发生动态变化。因此,他们比较了免疫应答者和无应答者之间卡介苗诱导的染色质重塑,分别针对先天性(即经过训练的免疫力)和对卡介苗接种的适应性反应。根据在治疗后第 90 天非特异性先天细胞因子产生能力(即用白色念珠菌、LPS 或金黄色葡萄球菌刺激后)的预定义阈值,将 213 名个体确定为免疫应答者,78 名个体确定为无应答者(图 4 A)。然后,他们研究了 BCG 诱导的经过训练的免疫反应者随时间推移的染色质可及性变化,以及基线时反应者和无反应者之间染色质可及性的差异(图4 B)。在经过训练的免疫应答者中,接种卡介苗在第14天导致4123个区域染色质可及性增加,3110个区域染色质可及性降低(图4B)。在第90天,当他们预计急性反应已经消退时,仍然检测到54个区域染色质可及性增加,20个区域染色质可及性降低(图4B)。在这个时间点,染色质可及性增加最强烈的基因组区域是GABBR1的启动子,GABBR1是g蛋白偶联受体家族的成员,在免疫过程中发挥多种作用,包括诱导pi3k -激酶- akt通路。该途径通过推动代谢向更高的糖酵解率转变,有助于体外bcg诱导的训练免疫。
在基线(第0天),免疫应答者与无应答者的区别在于,31个区域的染色质可及性较高,32个区域的染色质可及性较低(图4B)。应答者中几个染色质可及性较低的区域位于参与训练免疫的基因附近,包括MAPK和PI3K通路的成员。无应答者和应答者在基线时染色质可及性差异区域与应答者在第90天和基线时染色质可及性差异区域在统计学上有显著重叠(图4C),这表明在基线时无应答者中已经存在一些bcg诱导的应答者的变化。此外,基线时染色质可及性与细胞因子生产能力负相关的区域(图1H)与无应答者和第90天染色质可及性较低的区域显著重叠(图4D)。
他们使用bootstrapping对这些结果进行了敏感性分析,发现上面突出显示的示例始终位于排名靠前的区域中(图4E)。总之,他们的分析表明,一些个体在基线时携带的表观遗传细胞状态类似于bcg诱导的先天免疫记忆,这可能为进一步诱导训练免疫留下了很少的空间,从而解释了它们作为无应答者的状态。
在接种后第90天,根据bcg诱导的适应性细胞因子生产能力(即结核分枝杆菌刺激后的IFN-γ和IL-17)的增加,他们确定了217个个体为适应性免疫应答者,74个个体为无应答者。他们观察到卡介苗接种对3709个差异区域的适应性免疫应答者有很强的初始效果(第14天)(图4B),但这些差异在接种后第90天没有保留(图4B)。此外,在基线时,适应性免疫应答者和无应答者之间的染色质可及性没有统计学上的显著差异(图4B)。这些结果表明,表观遗传细胞状态与bcg诱导的先天免疫记忆的关系大于适应性免疫记忆。
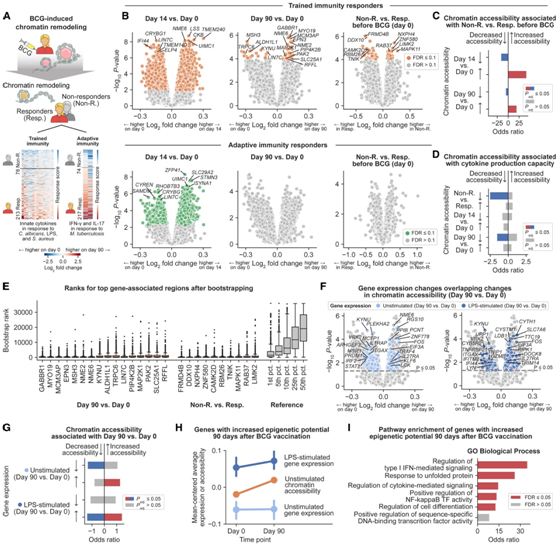
图4. BCG疫苗接种诱导与先天免疫记忆相关的染色质重塑。
(A) 疫苗接种后 90 天通过 BCG 诱导的细胞因子产生能力变化识别的应答者 (Resp.) 和无应答者 (Non-R.) 的示意图,分别针对先天性和适应性免疫刺激。(B) 火山图显示,在 BCG 疫苗接种后第 14 天(左)和第 90 天(中),受过训练的免疫应答者(上)和适应性免疫应答者(下)的基因组区域具有差异染色质可及性,以及无应答者与基线响应者(右)。(C) (1) 与基线时免疫应答者相比,无应答者染色质可及性较高(红色)或较低(蓝色)的基因组区域与 (2) 疫苗接种后 BCG 诱导的染色质可及性变化的区域之间存在重叠。(D) (1) 与基线时细胞因子产生能力最正相关(红色)或负相关(蓝色)的基因组区域和 (2) 与免疫应答者和 BCG 诱导的染色质重塑相关的区域之间的重叠。(D) (1) 与基线时细胞因子产生能力最正相关(红色)或负相关(蓝色)的基因组区域和 (2) 与免疫应答者和 BCG 诱导的染色质重塑相关的区域之间的重叠。(F) 火山图显示,在接种疫苗 90 天后,在没有(左)和有(右)LPS 刺激的情况下,BCG 诱导的 PBMC 基因表达发生变化,这些变化与 BCG 诱导的染色质可及性变化相关。(G) (1) 与疫苗接种后 90 天 BCG 诱导的染色质可及性变化最正相关(红色)或负相关(蓝色)的基因连锁区域和 (2) 与 BCG 诱导的基因表达变化相关的基因之间的重叠。(H) 卡介苗接种后 90 天,基因和基因连锁区域的平均染色质可及性和基因表达显示出表观遗传潜力增加。(I) BCG 疫苗接种 90 天后表观遗传潜力增加的基因的途径富集。
05
BCG 诱导的染色质重塑的生物学意义
为了将 BCG 诱导的染色质可及性变化与细胞功能联系起来,他们对差异区域进行了通路富集分析(图 5 A)。对于免疫反应者,在第 14 天和第 90 天,染色质可及性增加的区域与“mRNA 加工”、“mRNA 剪接”、“糖异生”、“葡萄糖代谢过程”和“典型糖酵解”相关(图 5 B) 。此外,与基线时的应答者相比,免疫非应答者的可及性较低的区域富含“T细胞激活”、“EGF激活受体活性”和“鞘糖脂生物合成过程”,表明这些过程在免疫中的潜在作用。他们通过使用 bootstrapping 的敏感性分析证实了这些观察结果的可重复性,并且上面突出显示的所有生物过程始终处于排名靠前的富集之中(图 5 C)。[1]
在免疫反应者中,BCG 疫苗接种后染色质可及性增加的区域富集了与 mTOR 信号传导相关的转录因子结合位点,包括 E2F1、AHR、NRF1、ARNT、HIF1A 和 MYC(图 5D)。在基线时的免疫应答者中染色质可及性较高的区域富含参与抑制炎症的转录因子(例如 NKX61)的结合位点。他们还对 PBMC 中的组蛋白标记图谱进行了富集分析,发现在接种 BCG 后染色质可及性增加的区域中与开放染色质相关的组蛋白标记同样富集(图 5D)。
BCG 诱导的染色质可及性增加的区域优先与先前在各种模型和数据集中与训练免疫有关的基因和基因组区域重叠(图 5 E)。例如,他们观察到人甲羟戊酸处理的巨噬细胞中的 H3K27 乙酰化和 BCG 处理的单核细胞中的 H3K4 三甲基化呈正相关。相比之下,之前描述为干扰训练免疫的基因显示出更封闭的染色质结构,这些基因的表达与染色质谱之间总体呈负相关(图 5 F)。
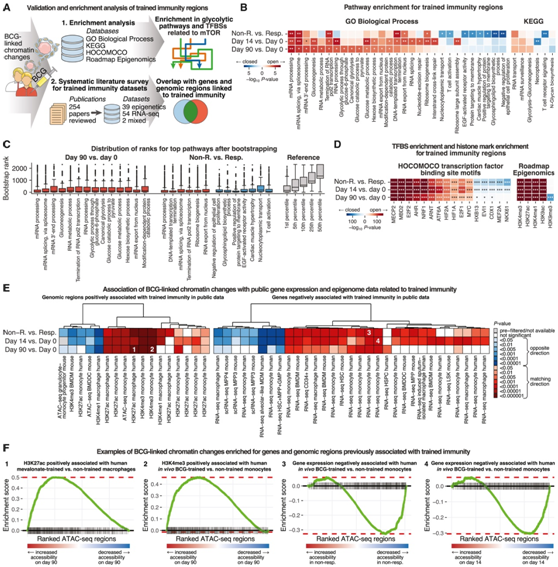
图5. BCG 疫苗接种后的染色质反应验证并扩展了小鼠和人类经过训练的免疫力的先验知识。
(A) 针对四个注释数据库(上)和文献整理的先前与训练免疫有关的基因和基因组区域的集合(下)进行富集分析的示意图。(B)免疫无反应者和反应者之间差异染色质可及性的基因集富集热图(上)以及反应者之间的 BCG 疫苗接种后(中和下)。(C) 箱线图显示了使用 bootstrapping 进行的敏感性分析中来自 (B) 的排名最高的基因本体 (GO) 生物过程富集的可重复性。(D) 基因组区域集富集的热图,类似于 (B) 中的基因集富集。(E) 本研究与之前的训练免疫研究之间的一致性热图。将经过训练的免疫无应答者和应答者(上)以及应答者中的 BCG 疫苗接种后(中和下)之间染色质可及性存在差异的区域与文献整理的基因组区域进行比较,这些区域在经过训练的免疫中染色质可及性增加或相应的组蛋白标记(左)以及文献整理的基因在经过训练的免疫中基因表达降低(右)。 (F) 与当前研究相比,先前报告中选定的基因组区域和与训练免疫相关的基因的基因集富集图。
06
BCG 疫苗接种反应的表观遗传预测因子
为了将患者分为免疫反应者和无反应者,他们将机器学习模型应用于多组学数据(图 6 A)。仅基于宿主因素的分类导致交叉验证的“受试者工作特征曲线下面积”(AUROC) 值为 0.54,仅比随机好一些(图 6 B)。添加季节性(疫苗接种日期)将性能提高到 0.64,可能反映了病原体暴露和免疫功能的季节性变化。当在基线处包含染色质可及性特征时,获得了最具预测性的模型,其 AUROC 为 0.76,并且比其他模型有统计上显著的增加。
染色质图谱的预测能力特定于经过训练的免疫反应,并不适用于适应性免疫反应:当他们对适应性免疫反应者与无反应者进行类似的机器学习分析时,AUROC 值没有超过随机机会,表明基线时免疫细胞的表观遗传状态与 BCG 疫苗接种的适应性免疫反应的强度无关,因此无法预测其强度(图 6 C)。
为了研究将表观遗传细胞状态与经过训练的免疫反应联系起来的生物过程,他们分析了经过训练的机器学习模型的回归系数,确定染色质可及性的主要成分 2 是经过训练的免疫反应的最强预测因子,其次是疫苗接种的季节性和时间(图 6 D)。富集分析将该主要成分与先天免疫途径联系起来,包括“中性粒细胞激活”和“补体和凝血级联”,其方向性使得基线水平较高预测成为免疫应答者的可能性较低(图6E)。总之,他们观察到对 BCG 疫苗接种的反应存在广泛的人群差异(图 6 F),并且确定了具有 BCG 诱导的训练免疫表型的个体子集,这是通过其基线染色质图谱预测的。
本项研究结果强调了在开发未来的治疗方法时进行患者分层的可行性和重要性,这些治疗方法可以通过表观遗传学增强对感染和癌症的免疫警觉。
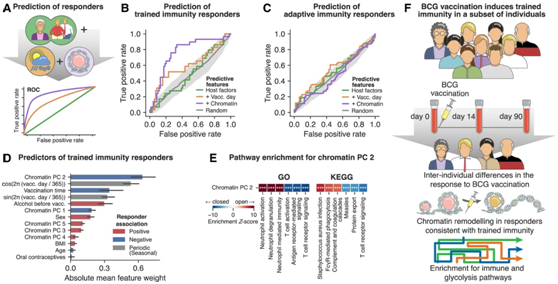
图6. 染色质可及性概况可预测经过训练的免疫应答者,但无法预测适应性免疫应答者。
(A) 基于机器学习的特征组(宿主因素、季节性、染色质可及性)评估的示意图,以评估其预测适应性免疫应答者的能力。(B-C) ROC 曲线比较具有不同输入特征集的模型,用于预测免疫应答者和适应性免疫应答者。(D)免疫反应者预测中的特征权重(重要性)条形图。(E) 有助于染色质可及性数据的主成分 (PC) 2 的基因组区域的途径富集。(F) 该研究的概要,该研究分析了基线时的个人免疫特征以及对卡介苗疫苗接种的反应,并确定了经过训练的免疫的表观遗传预测因子。
+ + + + + + + + + + +
结 论
本项研究用BCG对健康个体进行了免疫接种,这种减毒活疫苗不仅能诱导针对结核病的适应性免疫反应,还能触发先天免疫激活和记忆,这表明经过“训练”的免疫力。在 323 名个体的 90 天 BCG 疫苗接种过程中建立了个人免疫谱和染色质可及性图,揭示了基线免疫和免疫反应的遗传和表观遗传预测因子。BCG接种增强了先天免疫反应,特别是在基线时处于休眠免疫状态的个体中,而不是提供普遍的先天免疫增强。这项研究增进了对卡介苗异源免疫刺激作用和人类训练免疫力的理解。此外,它强调了表观遗传细胞状态对于将免疫功能与基因型和环境联系起来的价值。
+ + + + +



