

 English
English文献解读|Gut(24.5):妊娠期间肠道真菌群动态概况及其与宿主代谢和妊娠健康的关系
✦ +
+
论文ID
原名:Landscape of the gut mycobiome dynamics during pregnancy and its relationship with host metabolism and pregnancy health
译名:妊娠期间肠道真菌群动态概况及其与宿主代谢和妊娠健康的关系
期刊:Gut
影响因子:24.5
发表时间:2024.05.09
DOI号:10.1136/gutjnl-2024-332260
背 景
在正常怀孕期间,母体会发生剧烈的生理变化,包括免疫、激素和代谢变化。肠道菌群可视为一个虚拟器官,而怀孕状态与肠道菌群的显著改变有关。有假设认为宿主可以操纵肠道菌群来促进怀孕期间的代谢变化,最终支持胎儿的生长发育。值得注意的是,胃肠道中不仅有细菌,还有真菌。然而,与对肠道细菌的广泛研究相比,对真菌群落的研究仍然很少。作为肠道菌群的主要组成部分,肠道真菌在肠道生态中发挥着至关重要的作用,而肠道生态对宿主的健康至关重要。然而,关于妊娠期间肠道菌群的重塑,肠道菌群的动态及其与肠道微生物功能、宿主代谢、妊娠并发症和不良分娩结果的相互作用尚未得到很好的研究。
实验设计
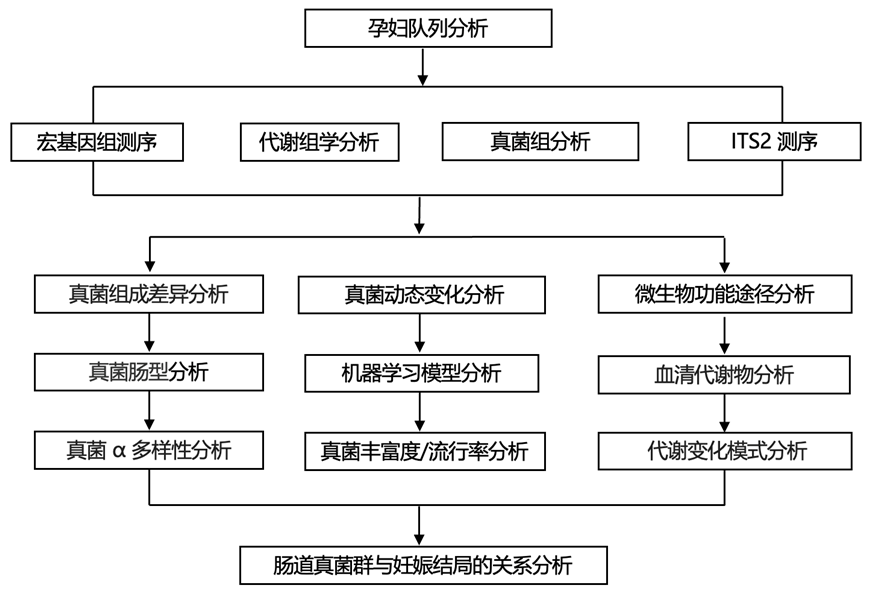
结 果
01

参与者的特征和肠道真菌群组成
本研究基于中国中部地区的一项前瞻性出生队列研究:同济-华西-双流出生队列 (THSBC)。THSBC 招募了在妊娠早期在当地妇幼保健院接受产前护理的孕妇。为了研究妊娠如何随时间影响肠道菌群并研究其与宿主代谢的潜在关联,研究团队建立了一个由 1059 名参与者组成的亚组,其中包括 514 名生育早产、低出生体重或巨大婴儿的女性,以及 545 名随机选择的未经历上述三种不良妊娠结局的参与者。对涉及 4800 名参与者的整个队列进行了 ITS2 测序,而对已建立的亚组中的妊娠前三个月 (T1) 样本进行了宏基因组学测序(图1)。
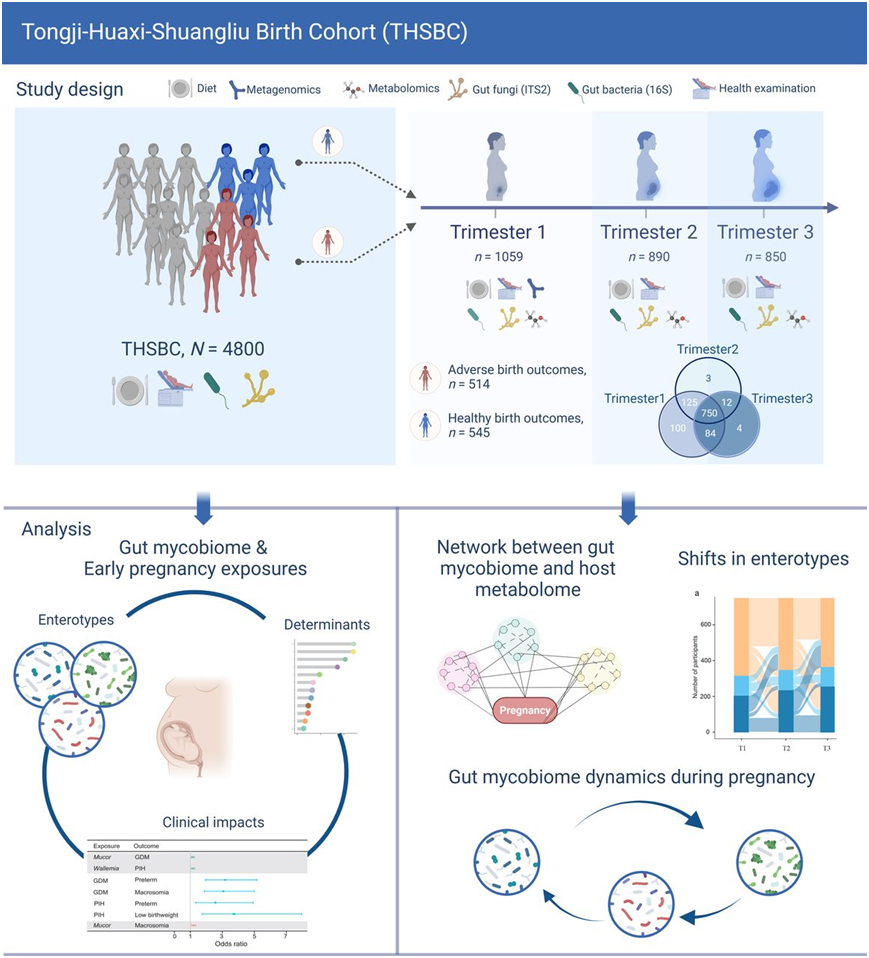
图1. 用于分析妊娠期间的肠道真菌并探索其与宿主代谢和健康的关系的研究流程图。
怀孕前,研究中的 947 名女性体重过轻,666 名女性超重或肥胖,而大多数女性体重正常。年龄和生育次数是导致肠道菌群组成个体间差异最重要的孕前人体测量因素。抗生素的使用和饮食因素(包括馒头、鸡蛋、水果、肉类和茶)也是重要的影响因素(图 2A)。
他们观察到三种真菌肠型,包括以酵母菌为主导的肠型(26.5%)、以念珠菌为主导的肠型(18.8%)和以曲霉菌为主导的肠型(54.7%)(图 2B-C)。生育史和饮食因素(包括馒头和茶)是影响真菌肠型的最重要因素。具体而言,妊娠早期具有以酵母菌为主导的肠型的女性更有可能是初产妇,并且在过去一年内有吃馒头的饮食偏好。另一方面,以念珠菌为主导的肠型的女性在过去一年内更有可能有以喝茶为特征的生活方式。值得注意的是,酵母菌、念珠菌和曲霉菌是三种最常见的真菌,而肠道菌群的总体流行率非常稀少(图 2D-E)。具体来说,他们在 T1共鉴定出 626 个真菌属,其中超过 96%的真菌属的流行率低于 40%。在后续分析中,他们将 40% 以上的参与者中存在的真菌属视为核心真菌属。
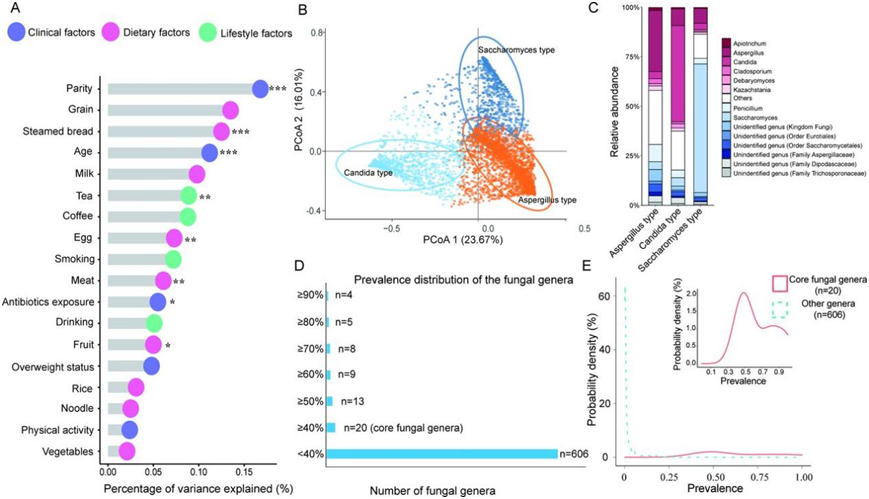
图2. 孕妇肠道真菌组组成和肠型分析。
(a) 真菌组组成的差异。(b) 通过主坐标分析 (PCoA) 可视化真菌肠型的聚类结果。(c) 显示每种肠型中最丰富的属。(d) 显示了在不同截止阈值下基于流行率的过滤中幸存下来的肠道真菌属的数量。(e) 展示了肠道真菌属的流行分布。
02
纵向亚队列中妊娠期间肠道真菌群的组成动态
他们研究了 750 名拥有所有妊娠期 ITS2 测序数据的参与者在怀孕期间的组成变化。结果显示,从 T1 到妊娠中期(T2),微生物群落组成发生了整体变化,但从 T2 到妊娠晚期(T3)并没有发生变化。在 750 名参与者中,多达 68.5% 的参与者在怀孕期间经历了真菌肠型的变化(图 3A)。具体来说,从 T1 到 T3,以酵母菌为主导的肠型比例增加,而以曲霉菌为主导的肠型比例减少,整个妊娠期间念珠菌为主的肠型比例相对稳定。
样本内 α 多样性(包括系统发育多样性和丰富度)从 T1 到 T3 大幅降低,而 Shannon 指数并没有明显动态变化(图 3B-D)。此外,在怀孕期间,生育健康婴儿的妇女和生下早产、低出生体重或巨大儿的妇女中,样本内 α 多样性的变化非常相似。虽然饮食因素导致了不同参与者肠道菌群组成的变化,但经过多重检验校正后,没有观察到从 T1 到 T3食物消耗量的变化与丰富度下降之间存在显著关联。因此,丰富度下降不太可能是由怀孕期间饮食的变化引起的。
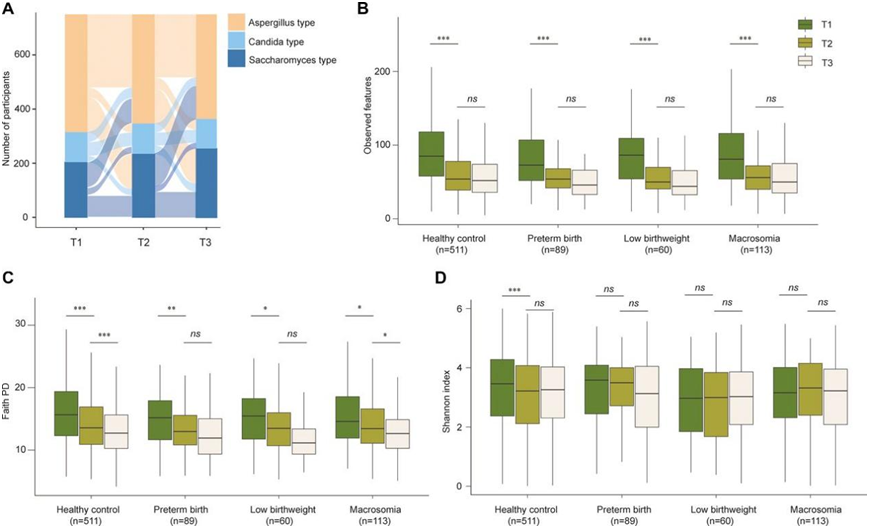
图3. 妊娠期间肠道真菌肠型的变化和肠道真菌 α 多样性的动态。
(a) 桑基图显示了从妊娠早期到妊娠晚期肠道真菌肠型的变化。 (b-d) 不同妊娠期肠道真菌 α 多样性的比较,展示了从妊娠早期到妊娠晚期肠道真菌 α 多样性的动态。
他们进一步在个体间变异的背景下分析了个体内组成的变化程度。从 T1 到 T2 或 T1 到 T3,个体内 Bray-Curtis 距离范围很广,其中大多数甚至大于单个时间点的平均个体间距离(图 4A)。相比之下,怀孕期间的肠道细菌组成更加保守。他们观察到组成变化和肠型转变之间的一致性。此外,他们发现与怀孕前体重过轻的女性相比,怀孕前超重或肥胖的女性从 T1 到 T3 经历的肠道真菌变化要大得多(图 4B)。
在 750 名拥有所有孕期肠道真菌组数据的参与者中,他们在 T1 样本中鉴定出 410 个属。然后,通过计算从 T1 到 T3 的损失率来评估每个属的不稳定性。对于 390 个不太流行的真菌属,平均损失率高达 97.6%,表明极度不稳定。相比之下,20 种核心真菌的平均损失率为 55.7%,其中曲霉菌、假丝酵母和酿酒酵母是最稳定的真菌属(图 4C)。
他们发现 20 个核心属中有 4 个发生了显著改变。具体来说,这些属是曲霉菌、枝孢菌、青霉菌和念珠菌,与妊娠早期相比,所有这些真菌属在妊娠晚期都发生耗尽,这发生在 40.4%–57.6% 的女性中。此外,他们使用机器学习算法(Light GBM)根据肠道真菌组组成区分 T1 和 T3 样本(图4D)。使用可解释的 Shapley Additive exPlanations (SHAP) 值,他们发现核心属占了所有前 10 个可区分的真菌属。然后,他们仅使用核心真菌属进行区分,其性能相当(AUC=0.725)(图 4D)。这些鉴别度最高的属大多在T1中呈现出较高的比例,并且大多属于子囊菌的Saccharomycetales、Eurotiales 和 Capnodiales。他们鉴定了前10个判别属在T1和T3的相对丰度和流行度(图4E)。此外,他们还对T1和T2样本、T2和T3样本进行了分类分析。这些比较的auc分别为0.78和0.61。在区分T1和T2样本的真菌属中,区分度最高的10个真菌属中有8个同时也是区分T1和T3样本的前10个真菌属,这些发现表明T2和T3样本的肠道菌群特征非常相似。
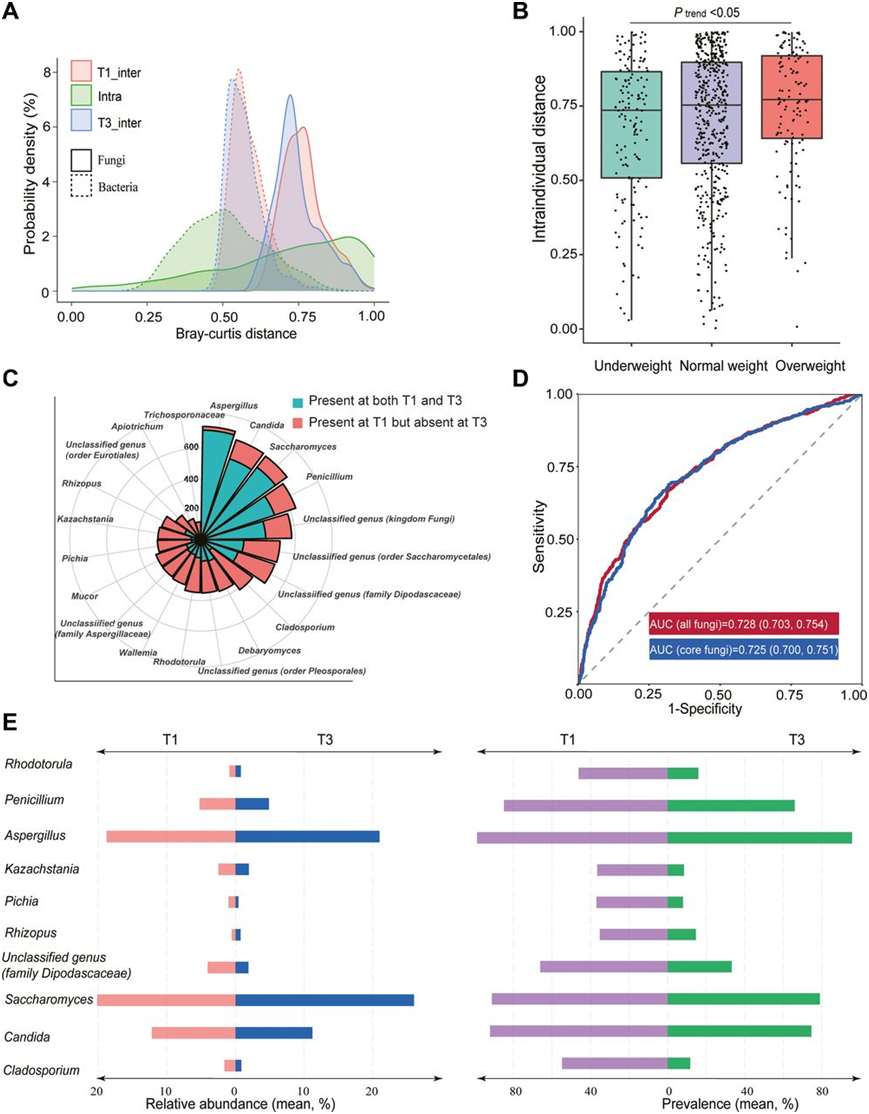
图4. 妊娠早期和妊娠晚期肠道真菌属的鉴别。
(a) 个体内肠道真菌和细菌组成随时间(从 T1 到 T3)变化的分布,以及 T1 或 T3 时个体之间的差异。(b) 按妊娠前超重状况分层,比较个体内肠道真菌组组成随时间(从 T1 到 T3)变化的程度。 (c)南丁格尔玫瑰图可视化了与妊娠早期相比,妊娠晚期核心肠道真菌属丢失的参与者比例。 (d)使用机器学习框架,特别是 LightGBM,对 T1 和 T3 时肠道真菌组组成的妊娠期分类器进行训练。 (e) 该图显示了对 T1 和 T3 妊娠期分类有贡献的前 10 个肠道真菌属的相对丰度(左)和流行率(右)。
03
与肠道真菌群相关的关键微生物功能途径和宿主血清代谢物
为了进一步表征真菌肠型与单个肠道微生物功能途径之间的关系,他们确定了 8 条在不同肠型中分布不同的途径。具体而言,这些途径中的大多数在以酵母菌为主的肠型中相对富集,而在以曲霉为主的肠型中较少富集,包括硫胺素回收IV、蔗糖降解IV、L-赖氨酸生物合成I、L-苯丙氨酸生物合成途径、肽聚糖成熟、C4 光合碳同化循环和硒氨基酸生物合成。同时,蔗糖降解IV和硒氨基酸生物合成在以念珠菌为主的肠型中的富集程度也很高,而 preQ0 生物合成在以念珠菌为主的肠型和以曲霉为主的肠型中的富集程度都很高(图 5A)。此外,进一步的回归分析发现,所识别的途径与 27 种血清代谢物之间存在 95 个显著关联(图 5A)。超过2 / 3的显著代谢物是胆汁酸(如鹅脱氧胆酸、异脱氧胆酸、脱氧胆酸、猪脱氧胆酸和甘胆酸)、氨基酸代谢物(如4-羟基马尿酸和苯乙酰- L-谷氨酰胺)和有机酸衍生物(例如3-吲哚丙酸和吲哚丙烯酸)(图 5A)。
当他们在已建立的亚群中反复测量血清代谢组时,描绘了代谢变化并探索了其与肠道真菌的共变关系,定量了 794 种已鉴定的代谢物,它们属于不同的生化类别,例如氨基酸、脂质、核苷酸和碳水化合物。与肠道真菌组类似,血清代谢组在妊娠不同时期也发生了显著变化,并且 T1 和 T2 之间的变化远大于 T2 和 T3 之间的变化(图 5B)。不同类别代谢物的妊娠时期解释方差比例范围为 1.17% 至 11.89%,其中激素及其相关代谢物排名靠前(图 5C)。单个核心真菌属与单个代谢物之间的相关性分析显示 30 个共变关系,涉及 6 个属和 27 种血清代谢物(图 5D)。鉴定的6种真菌中有4种注释到属水平,包括枝孢霉属、曲霉菌属、根霉菌属和毛霉菌属。具体而言,曲霉、根霉菌和毛霉菌分别与3种、8种和1种代谢物呈正相关,而枝孢霉则与7种代谢物呈负相关。根霉菌相关代谢物多为脂酰,枝孢霉相关代谢物主要为氨基酸或其代谢物。
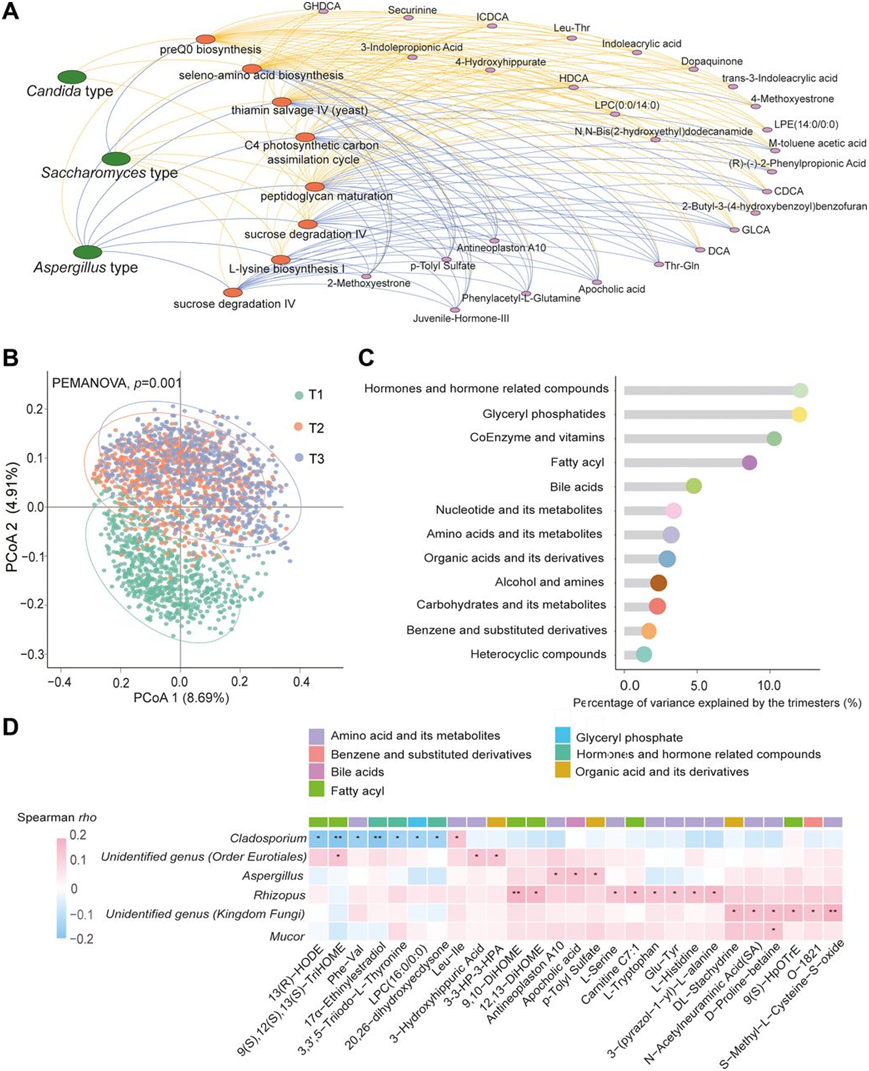
图5. 妊娠期间肠道真菌群与宿主代谢之间的相互作用。
(a) 肠道真菌肠型、微生物功能和宿主代谢组之间的网络分析。 (b) 比较在妊娠不同阶段采集的血清样本的整体代谢模式。(c) 基于堪培拉差异性,不同类别代谢物在妊娠期间随时间发生的代谢改变程度。(d) 从妊娠前三个月到妊娠晚期,各个核心真菌属和各个血清代谢物之间的共变关系热图。对核心肠道真菌的变化和代谢物的变化进行了 Spearman 相关性分析。
04
肠道菌群与妊娠结局的关系
与体重偏轻、正常和超重女性的肠道菌群组成个体内变异不同的发现类似,他们发现怀孕前超重显著影响了从 T1 到 T3 的代谢变化模式(图 6A)。随后,进行了分析,以确定哪些代谢物表现出依赖于怀孕前超重状态的显著变化,研究结果显示,23 种特定代谢物(例如 L-甘氨酸、L-精氨酸、十六烷二酸和肉碱)主要属于氨基酸和脂肪酰,仅在体重偏轻的女性中表现出显著变化(图 6B)。相比之下,24 种主要属于氨基酸和苯衍生物的代谢物(例如 L-缬氨酸、L-谷氨酰胺、2-羟基肉桂酸和 4-羟基苄醇)仅在超重女性中发生显著改变(图 6B)。因此,这些代谢物可能是导致体重过轻或超重女性中观察到的代谢变化模式不同的原因。
通过在整个队列中研究妊娠早期各核心真菌属与妊娠并发症的前瞻性关联,他们发现毛霉菌与新发妊娠期糖尿病(GDM)之间呈显著正相关,Wallemia与妊娠高血压(PIH)呈正相关(图6C)。此外,他们验证了GDM和PIH都是不良出生结局的危险因素,包括巨大儿和早产,GDM与巨大儿和早产的发生风险显著相关。因此,他们进行了中介分析,以检验毛霉菌对胎儿过度生长的影响是否由 GDM 介导,妊娠早期肠道真菌毛霉菌与巨大儿风险相关,与 GDM 无关,这表明毛霉菌和 GDM 可能通过不同的途径影响巨大儿风险(图 6D)。他们还探讨了亚队列中肠道菌群组成改变程度与不良出生结局之间的关联,但未得到显著结果。这一发现可能表明,无论出生结果如何,整体肠道菌群组成的转变是一种由妊娠驱动的广泛现象。然而,这并不排除某些肠道真菌的轨迹可能与妊娠健康有关,因为他们发现早产和非早产婴儿的孕妇之间的毛霉菌轨迹存在显著差异(图6E)。

图6. 按妊娠前超重状态分层的独特代谢变化以及妊娠期间肠道真菌群的临床意义。
(a) 亚组间整体代谢动态比较。(b) 维恩图显示从妊娠前三个月到妊娠晚期发生显著变化的独特和常见代谢物的数量。(c) 核心真菌属与妊娠并发症以及不良出生结局之间的关系。(d) 肠道真菌属Mucor、GDM 和巨大胎儿之间的中介分析。 (e) 曲线显示 LOESS 拟合基于早产(绿色)或否(红色)的已识别分类群的相对丰度。
+ + + + + + + + + + +
结 论
本项研究发现了肠道真菌的组成从妊娠早期到妊娠晚期发生了巨大变化,与肠道细菌的变化相比,其变化程度和个体性更大。多组学数据提供了肠道真菌群、生物功能、血清代谢物和妊娠健康之间的网络图景,精确指出了毛霉菌与不良妊娠结局之间的联系。妊娠前超重状态是影响肠道真菌群组成变化和妊娠期间代谢重塑模式的关键因素。本研究提供了妊娠期间肠道真菌群动态及其与宿主代谢和妊娠健康的关系的概况,为未来开展肠道真菌群研究以实现健康怀孕奠定了基础。
+ + + + +



