

 English
English文献解读|Sci Immunol(24.8):鼻细胞图谱揭示簇状细胞的异质性及其在指导嗅觉干细胞增殖中的作用
✦ +
+
论文ID
原名:A nasal cell atlas reveals heterogeneity of tuft cells and their role in directing olfactory stem cell proliferation
译名:鼻细胞图谱揭示簇状细胞的异质性及其在指导嗅觉干细胞增殖中的作用
期刊:Science Immunology
影响因子:24.8
发表时间:2024.02.02
DOI号:10.1126/sciimmunol.abq4341
背 景
嗅觉神经上皮作为气味的感觉器官,并形成鼻黏膜屏障的一部分。嗅觉感觉神经元由上皮细胞包围和支持。其中,微绒毛细胞(MVC)位于顶端表面,但它们的具体功能是未知的,并且它们与其他特化上皮细胞的关系尚不清楚。
实验设计
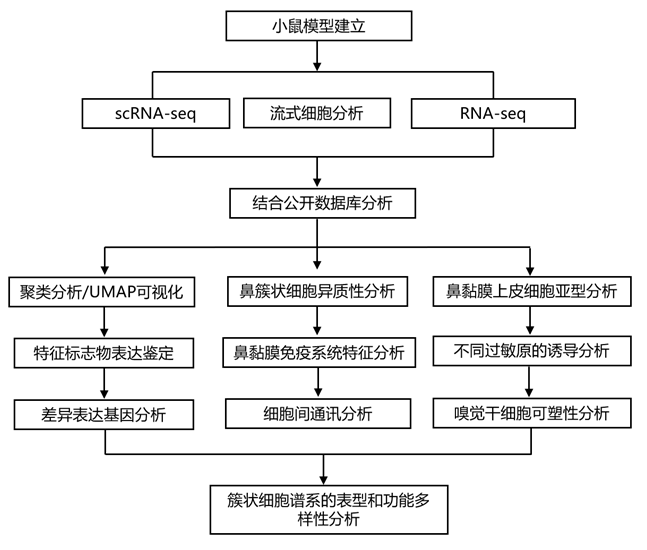
结 果
01

小鼠嗅觉和鼻呼吸道细胞的转录组分析
为了确定小鼠嗅觉和鼻呼吸化学感觉细胞ChAT-eGFP+的转录谱,研究团队沿着嗅鼻甲前部从呼吸黏膜中分离出嗅觉细胞,并从每个隔室中获得单细胞悬液。ChAT-eGFP +细胞在嗅黏膜中大量存在(EpCAM High细胞的 22%),它也在呼吸道黏膜中普遍存在(EpCAM High细胞的 10% 和活细胞的 1%)。然后,他们通过单细胞转录组分析(scRNA-seq)将ChAT-eGFP +细胞与来自嗅觉和呼吸道上皮细胞的EpCAM high细胞进行比较(图1A)。无监督聚类鉴定出TRPM5上皮细胞的一个亚群,其表达已知的簇状细胞标记,与来自呼吸和嗅觉黏膜的所有其他EpCAMhigh细胞明显聚集(图1A)。他们使用随机森林分类器进行全转录组分析,将TRPM5上皮细胞聚类中94%的细胞(本身占所有细胞的44.1%)定位为气管簇状细胞(图1B)。除了簇状细胞外,他们还发现了另一群丰富的分化上皮细胞,这些细胞不表达Chat和Trpm5,但表达Ascl3和Nt5e (CD73),这与瞬时受体电位阳离子通道亚家族M成员5 (TRPM5)- MVC的描述一致。他们将其中96%的细胞映射到肺离子细胞(图1B),明确地将它们鉴定为鼻离子细胞(图1B)。
鼻部的簇状细胞和离子细胞明显多于气管。为了表征它们的实际分布,他们评估了来自鼻子的非富集scRNA-seq数据集中的簇状细胞和离子细胞的数量,并与已发表的气管scRNA-seq数据进行了比较。他们发现,簇状细胞占嗅上皮中EpCAM+细胞的4.7%,是气管中EpCAM+细胞的3.7倍(图1C)。嗅觉上皮中的离子细胞占EpCAM+细胞的5.4%,比气管中的离子细胞多12.3倍(图1C)。本研究鉴定的大量鼻簇状细胞表达了气管和肠道簇状细胞的典型标记物:肌动蛋白结合蛋白advillin、Avil、Trpm5、花生酸类生物合成酶Alox5ap和Ltc4s(图1D)。鼻离子细胞(TRPM5-MVC)表达高水平的Cftr、Coch和Foxi1(图1D)。免疫荧光证实,cochlin+和CFTR+(囊性纤维化跨膜传导调节因子阳性)离子细胞样上皮细胞和ChAT-eGFP+/TRPM5+ MVC在嗅觉上皮中大量存在(图1E),而在呼吸系统上皮中离子细胞数量较少。其余的EpCAMhigh细胞由分化的上皮细胞、基底细胞和少数嗅觉标记蛋白(OMP+) 嗅觉感觉神经元(OSN)组成。
嗅觉和呼吸系统中的 Trpm5+/Chat+ 细胞共有一个核心转录程序,该程序由参与味觉传导的转录本(Trpm5 和 Gng13)、钙信号分子(Lrmp、Pik3cg、Hck、Vav1、Matk 和 Pik3r5)、转录因子(Spib、Pou2f3 和 Pou2af2)以及 CysLT 生物合成酶(Alox5、Alox5ap 和 Ltc4s)定义(图1F)。气管和肠道中的簇状细胞共有这一核心效应谱(图1G)。所有黏膜隔室中的簇状细胞都通过表达必要的机制来区分,以快速产生钙信号依赖的介质-类二十烷酸和乙酰胆碱。
为了确定人类 MVC 是否细分为簇状细胞和离子细胞,他们分析了公开数据集。对标记为MVC的116个细胞聚类进行集中分析,确定了两个亚群(图1H)。其中一个亚群(包含30个细胞)与小鼠簇状细胞高度相似,具有不同的簇状细胞标记物AVIL和LRMP、类二十碳酸途径组分ALOX5AP和PTGS1、IL-25受体IL17RB和簇状细胞转录因子POU2F3的表达。在由NCAM1和OMP标记的嗅神经上皮中,advillin+ 细胞分布于OSN之间(图1I)。这些嗅觉簇状细胞具有类似嗅觉TRPM5+ MVC的球状形状,表明它们具有共同的形态(图1I)。FOXI1+离子细胞也在人嗅觉黏膜中发现,分散在NCAM1+ OSN之间(图1J)。在呼吸道上皮中,adillin和dclk1阳性的簇状细胞呈梭形(图1K-L)。他们还在人类活检标本的呼吸道上皮中发现了FOXI1+离子细胞(图1M)。综上所述,这些结果证实了人类簇状细胞和离子细胞代表了之前描述的嗅黏膜中人类MVC的两个不同的亚群。与小鼠一样,嗅簇状细胞和呼吸簇状细胞具有共同的转录谱,离子细胞也是如此。
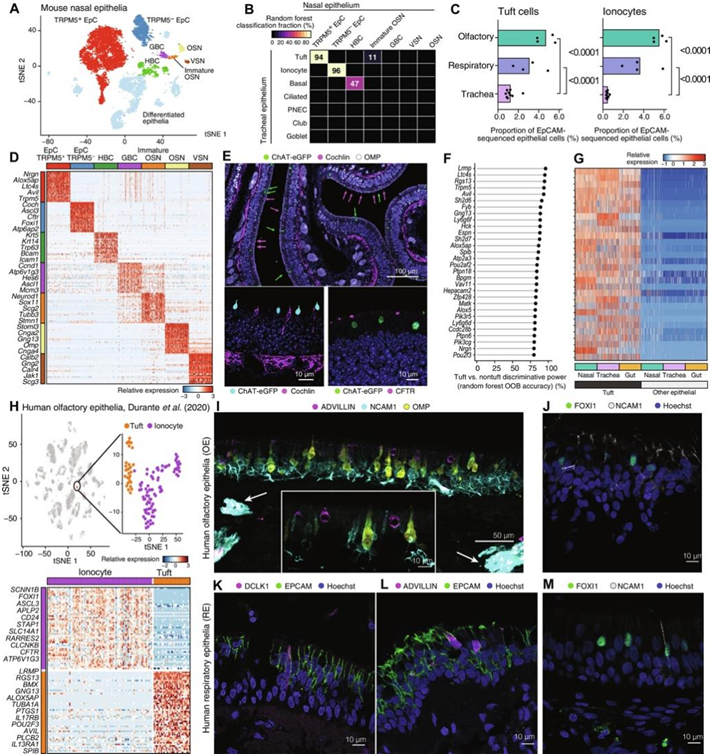
图1. 簇状细胞和离子细胞构成小鼠和人类嗅觉上皮的两种 MVC 类型。
(A) 13052 个细胞的t分布随机邻域嵌入 (tSNE) 嵌入,通过无监督聚类进行着色和标记。 (B) 热图显示使用随机森林分类为每种气管细胞类型(行)的每种鼻上皮细胞类型(列)。(C)来自幼稚鼻黏膜中的EpCAM+细胞的每只小鼠(点)的所有上皮细胞(点)中的簇状细胞(上)和离子细胞(下)的分数(x轴)。(D)数据集中特定于每种细胞类型的前 50 个基因(行)的相对表达。(E)免疫荧光染色。(F) 随机森林分类。(G) 三种组织中簇状细胞和其他上皮细胞中簇状细胞标记基因的相对表达。(H) 3528 个嗅觉上皮细胞、免疫细胞和神经元细胞的 tSNE 嵌入(上),显示了对包含簇状细胞和离子细胞的 116 个 MVC 聚类的新重新分析以及差异表达基因的热图(下)。(I-M) 免疫荧光分析。
02
小鼠鼻簇状细胞的三个亚群分别对应于MVC、SCC和腺簇状细胞群
单细胞分析表明,所有Trpm5+/Chat+鼻上皮细胞聚集在一起,并共有簇状细胞转录谱。簇状细胞的无监督聚类确定了三个亚组(图2A),每个亚组有不同的标记(图2B)。他们将簇状细胞的优势群体定义为TRPM5+ MVC(图2A),因为它们主要来源于嗅觉黏膜,并且不表达孤立化学感应细胞 (SCC)的广泛味觉受体(图2B)。TRPM5+ MVC的特征是簇状细胞标志物TRPM5、Pou2f3和Chat(所有TRPM5+ MVC的原始标志物和泛簇状细胞标志物Avil)的高普遍表达(图2B-C)。这些TRPM5+ MVC在大约50%的细胞中以低水平表达味觉受体相关蛋白Gɑ-gustducin Gnat3(图2C)。它们还表达高水平的类花生酸生物合成酶Ptgs1、Hpgds、Alox5、Alox5ap和Ltc4s和簇状细胞细胞因子Il25(图2B-C)。使用机器学习分类器对簇状细胞MVC和气管簇状细胞亚群进行的比较显示,与之前描述的富含类花生酸的簇状细胞2 (tuft-2)的相似性最高(图2D)。由于TRPM5 MVC共有MVC的位置和形态以及簇状细胞的核心转录谱,他们将它们称为簇状MVC。
他们鉴定了一个之前未描述的簇状MVC标记:Nrgn,它编码神经颗粒素(一种钙调蛋白家族中的突触后神经元蛋白),并在簇状细胞缺陷的Pou2f3-/-小鼠中验证了其在蛋白水平的表达和缺失(图2B-F)。呼吸道上皮内可见散在的罕见神经颗粒细胞(图2E-H)。出乎意料的是,簇状MVC中Dclk1和编码的Dclk1蛋白的水平都非常低,而Dclk1蛋白通常用于识别肠道和气管中的簇状细胞(图2C-I)。在嗅上皮中,DCLK1标记具有导管形态的上皮细胞(图2I),表明DCLK1在鼻腔不是簇状细胞特异性标记物。
第二种更罕见的簇状细胞群仅来源于呼吸道上皮,并且高度富集于定义长形鼻呼吸道SCC的转录本,包括1型(Tas1r1和Tas1r3)和2型(Tas2r104、Tas2r105、Tas2r108、Tas2r118和Tas2r138)味觉受体、琥珀酸受体Sucnr1,以及普遍且高表达的肠收缩素成分Gnat3和Gnb3(图2B-C)。这些细胞具有纺锤形形态,并且也富含核心簇状细胞基因(图2C)。由于鼻呼吸道SCC的这种形态和标志物表达特征,结合簇状细胞的转录谱,他们将其称为簇状SCC。簇状SCC与味觉受体富集的气管簇状-1-细胞最为相似(图2D)。组织学上,簇状SCC大部分为神经颗粒素阴性,但通过ChAT-eGFP、adillin和Gɑ-gustducin进行标记(图2K-L)。
最后,他们鉴定了一个转录中间群体的簇状细胞,该群体高表达Dclk1,并富集了Il13ra1,负责在肠道中2型炎症诱导的簇状细胞扩增(图2B)。它们也富含Ly6d和Ly6e,这是与乳腺癌和前列腺癌相关的转录本。他们在侧鼻腺(LNG)中发现了一种独特的簇状细胞群,这是一种小鼠鼻腔特有的大型浆液腺结构(图2I)。这些ChAT+/advillin+的簇状细胞也表达神经颗粒素,Gɑ-gustducin和DCLK1,因此对应于转录中间簇状细胞群(图2C-J)。Ltc4s mRNA在鼻簇状细胞的所有亚群中普遍表达:嗅觉黏膜的簇状上皮细胞、LNG的簇状腺细胞和呼吸上皮的簇状上皮细胞(图2M-O)。
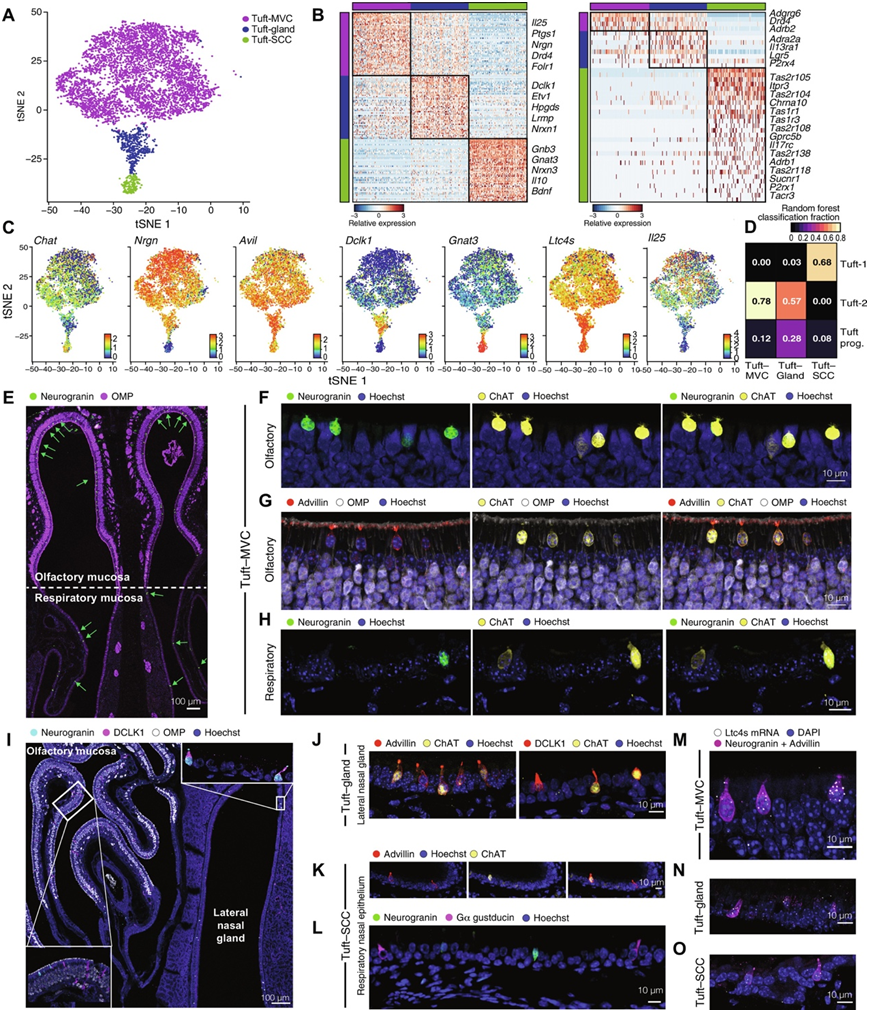
图2. 单细胞分析鉴定小鼠鼻簇状细胞异质性。
(A) 源自 ChAT-eGFP 小鼠的 5594 个簇状细胞的 tSNE 嵌入,通过无监督聚类着色。 (B) 热图显示每个特异性表达的标记物(左)和 G 蛋白偶联受体基因(右)的表达水平。(C) tSNE 图显示共有簇状细胞标记和亚型标记的表达。(D) 热图显示使用随机森林分类为每个气管细胞亚型(行)的每个鼻簇状细胞亚型(列)。(E-L) 簇状细胞亚群的免疫定位。(M-O) 用 RNAscope 探针对Ltc4s进行原位杂交。
03
小鼠鼻黏膜免疫系统在稳态和吸入过敏原反应中的特性
接下来,他们试图绘制鼻黏膜免疫和神经上皮系统在稳态和吸入过敏原反应期间的图谱,在稳态状态下,以及在先天期(单次吸入后36小时)和适应性期(两次吸入后36小时,间隔6天)鼻腔吸入霉菌过敏原alternnaria alternata后进行scRNA-seq实验。他们总共分析了50448个高质量的上皮细胞和免疫细胞以及osn(图3A)。无监督聚类分析检测到11种骨髓细胞亚型和17种淋巴样细胞亚型(图3B-C)。在稳态的髓细胞室中,巨噬细胞最多(13%),其次是组织常驻的常规中性粒细胞(11.7%),另外一组是Siglec F+中性粒细胞(4.3%)(图3B-D)。淋巴细胞集中在鼻呼吸道部分,可能与鼻淋巴组织有关(图3E)。ILC2亚群分别占嗅觉上皮和呼吸上皮造血细胞库的5%和6%。在这项研究中,除了TH1细胞外,他们检测到了所有标准的T辅助细胞(TH)亚群,还观察到一群表达高水平Il17a和Rorc的γδ T细胞,类似于TH17细胞。这两种表达Il17a的细胞亚群在呼吸系统中富集,类似于ILC1和自然杀伤细胞(图3E)。
单次吸入1天后,免疫细胞亚群比例相对稳定(图3F)。在炎症的适应期(第二次吸入Alternaria后36小时),TH2细胞和调节性T细胞增加了一倍(图3F),而几乎所有髓系细胞的相对比例都减少了(图3F),这可能反映了它们的绝对数量稳定。最后,他们在不同类型的鼻黏膜细胞中对气致过敏原诱导的细胞间信号转导进行了研究(图3G)。他们绘制了所有在CellPhoneDB 2.0或FANTOM5数据库中标注的受体-配体基因对,这些基因对Alternaria表现出差异表达。除了经典的细胞因子Il5和Il13外,与2型炎症相关的其他效应信号也由ILC2上调,包括Calca和Il17rb(图3G)。总之,这些数据表征了特定的细胞类型和途径,构成了鼻黏膜对Alternaria的2型极化反应。
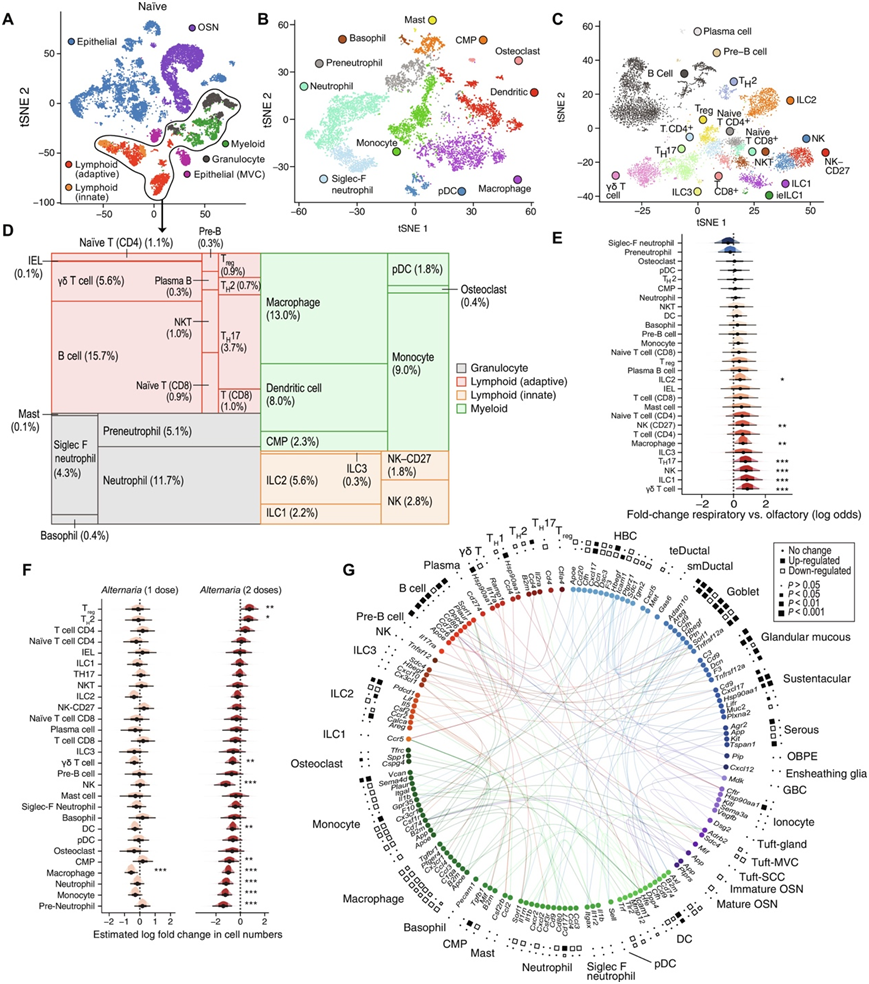
图3. 小鼠鼻黏膜过敏性炎症期间细胞间通讯和成分变化的单细胞分析。
(A) 通过 scRNAseq 评估来自幼稚小鼠鼻黏膜的 19275 个细胞的 tSNE 嵌入;显示了通过无监督聚类识别的细胞类型谱系。 (B-C) 7107 个骨髓和 7213 个淋巴细胞的 tSNE 嵌入(点)。(D) 树状图显示了幼鼠免疫区室的细胞类型组成。(E-F) 组织位置和吸入一剂和两剂Alternaria 对每种免疫细胞类型比例的影响。(G)圆形图显示Alternaria吸入后细胞类型之间差异表达的受体配体相互作用。
04
小鼠鼻黏膜上皮细胞亚型的定位
接下来,他们定义了嗅觉上皮细胞的多样性(图4A-J)。支撑细胞、簇状上皮细胞、离子细胞、导管细胞和球状基底细胞主要来源于嗅觉黏膜(图4B)。嗅觉Cyp2g1+细胞由两大亚群组成,它们具有不同的特征:第一,支撑带细胞由Cyp2g1、Muc2和Il33标记;其次,Cyp2g1+群体Muc2阴性表达Bowman腺导管细胞标志物Aqp4和Aqp5(图4A-G)。Bowman腺导管细胞也意外地呈DCKL1阳性(图4F-G),DCKL1是一种簇状细胞标志物。因为它们位于osn和上皮细胞之间,他们称它们为经上皮导管细胞。Bowman腺导管细胞富含PGD2受体DP1 (Ptgdr),与人类粘液分泌有关。他们发现了一种以前未发现的黏膜下导管细胞群,它们在表达杀菌/通透性增加蛋白(BPI)折叠家族成员bpifb3、Bpifb4和Bpifb6方面存在差异(图4A-G)。
他们鉴定了另外两种类型的粘液细胞:Gp2+杯状细胞和Muc5b+/Tff2+腺状粘液细胞(图4B)。Gp2+杯状细胞大量分布于呼吸道上皮(图4C-E),鼻部后部可见LNG细胞。Muc5b+腺黏膜细胞特异性表达三叶因子Tff2和Bpifa1 (SPLUNC1),Tff2与粘蛋白协同分泌,参与上皮修复,Bpifa1 (SPLUNC1)对鼻上皮防御很重要。他们使用Muc5b蛋白作为腺黏膜细胞的标记物,发现在(i)嗅上皮鲍曼腺的腺泡细胞,(ii)呼吸黏膜下腺和(iii) LNG的腺泡细胞中表达阳性(图4H-J)。
最后,他们鉴定了一个由气味结合蛋白(OBP)编码基因标记的上皮细胞群:Obp1a、Obp1b和Obp2a(图4A)。obp可逆地将气味剂结合到osn中。此外,他们在间隔黏膜下腺中发现了高表达的Obp1a(图4K-L)。然后,他们分析了嗅觉和呼吸黏膜对Alternaria空气过敏原吸入的反应。在一次和两次Alternaria剂量后,上皮中的Muc5b腺黏膜细胞分别增加了1.4倍和3.9倍,两次Alternaria剂量后,Gp2杯状细胞增加了2.9倍(图4M)。差异表达分析区分了细胞类型特异性和细胞类型非依赖性作用,发现Muc5b+腺粘液细胞和Gp2+杯状细胞是反应最强烈的细胞类型,几丁质酶样蛋白4 (Chil4)是多种细胞类型中上调最强烈的基因(图4N-O)。在肺2型炎症中,在il13过表达系统的鼻黏膜中,Chil4增加。此外,吸入Alternaria诱导S100a6、Sprr1a和Sprr2a3(图4N)。
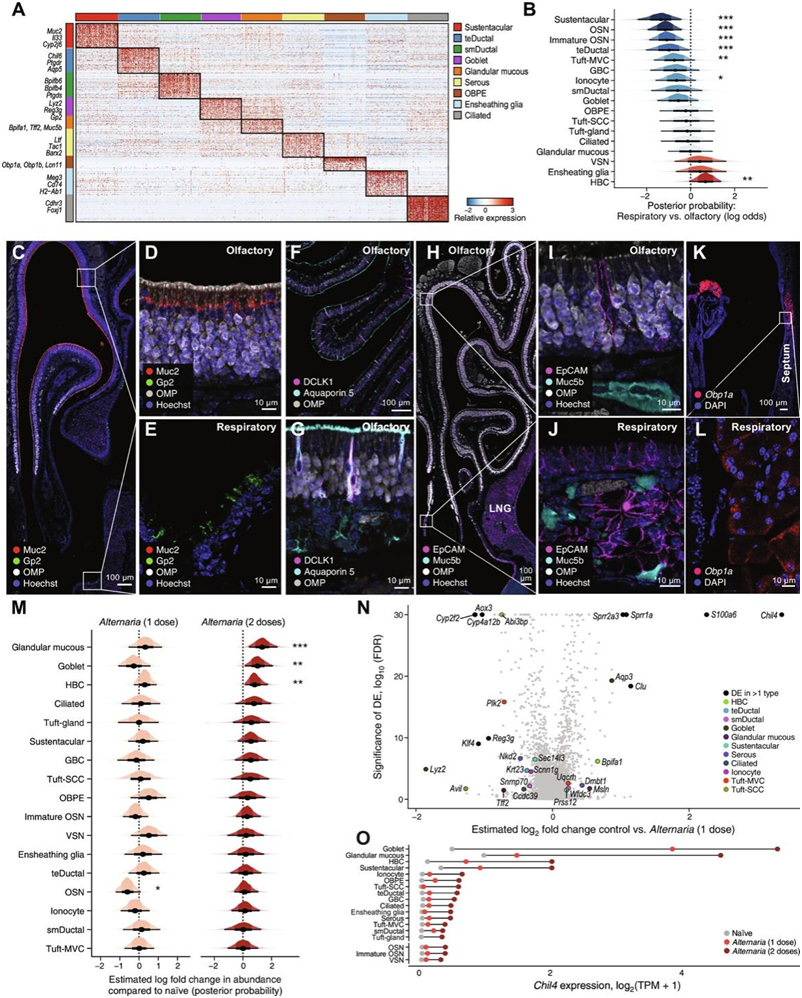
图4. 绘制小鼠鼻黏膜分泌上皮细胞的亚型图。
(A) 热图显示每个非感觉上皮子集特异性表达的前 30 个标记基因(行)的表达水平。(B)组织位置对每种上皮细胞类型比例的估计影响。(C-L)嗅觉和呼吸道鼻黏膜中上皮细胞亚群的免疫定位。(M)吸入一剂(左)和两剂(右)Alternaria对上皮细胞亚群比例的估计影响。(N) 火山图显示了吸入一剂Alternaria后上皮亚群(颜色图例)内基因(点)的差异表达和显著性之间的关系。(O)吸入Alternaria前后几丁质酶样蛋白 4 (Chil4) 的表达水平。
05
吸入过敏原可诱导HBC增殖
吸入过敏原曲霉菌(Aspergillus)导致快速OSN和上皮细胞死亡组织学评估。为了确定Alternaria是否会引起类似的神经上皮结构破坏,他们通过组织学评估了嗅觉和呼吸黏膜的完整性。嗅觉神经上皮呈多层结构,包括覆盖在一到两层未成熟嗅觉感觉神经元(ISN)上的OMP+嗅觉感受神经元(OSN),以及一层增殖的Ki67+球状基底细胞,这种结构在Alternaria作用后得以保留(图5A)。此外,在scRNA-seq中检测到的OMP层厚度、OMP+ OSN和支撑细胞的比例均未发生明显变化,提示没有发生严重的OSN和上皮细胞损失。使用TUNEL(末端脱氧核苷酸转移酶介导的脱氧尿苷三磷酸缺口末端标记)的评估显示,单次吸入Alternaria后,呼吸上皮细胞出现凋亡,而嗅觉上皮细胞未出现凋亡。
在组织学上,最显著的变化发生在嗅神经上皮基底层,吸入Alternaria后Ki67+细胞数量明显增加(图5A-B)。吸入Alternaria后,角蛋白5 (Krt5) +潜伏状态的嗅觉水平基底水平干细胞(HBC)增殖,而增殖的免疫或非免疫细胞(包括Ki67+Kit+球状基底细胞)的数量没有显著变化(图5C-F)。最后,他们还发现,参与细胞增殖的基因表达程序在嗅上皮内的HBC中发生强烈和特异性地诱导。Krt5+增殖细胞在嗅觉上皮中的比例增加了6.1倍,而在呼吸上皮中仅增加了1.8倍(图5G-H)。转录因子p63 (Trp63)通过抑制HBC分化促进嗅觉干细胞自我更新,其缺失先于HBC增殖。与空气过敏原诱导的HBC激活一致,他们发现Alternaria感染后HBC中的Trp63表达显著降低(图5I)。差异表达(DE)分析显示,许多细胞周期基因由HBC上调(图5J),通路分析证实,增殖程序最显著富集(图5K)。综上所述,这些数据表明,吸入Alternaria可在36至72小时内诱导正常静止的HBC增殖,但不会导致osn或支撑细胞凋亡。
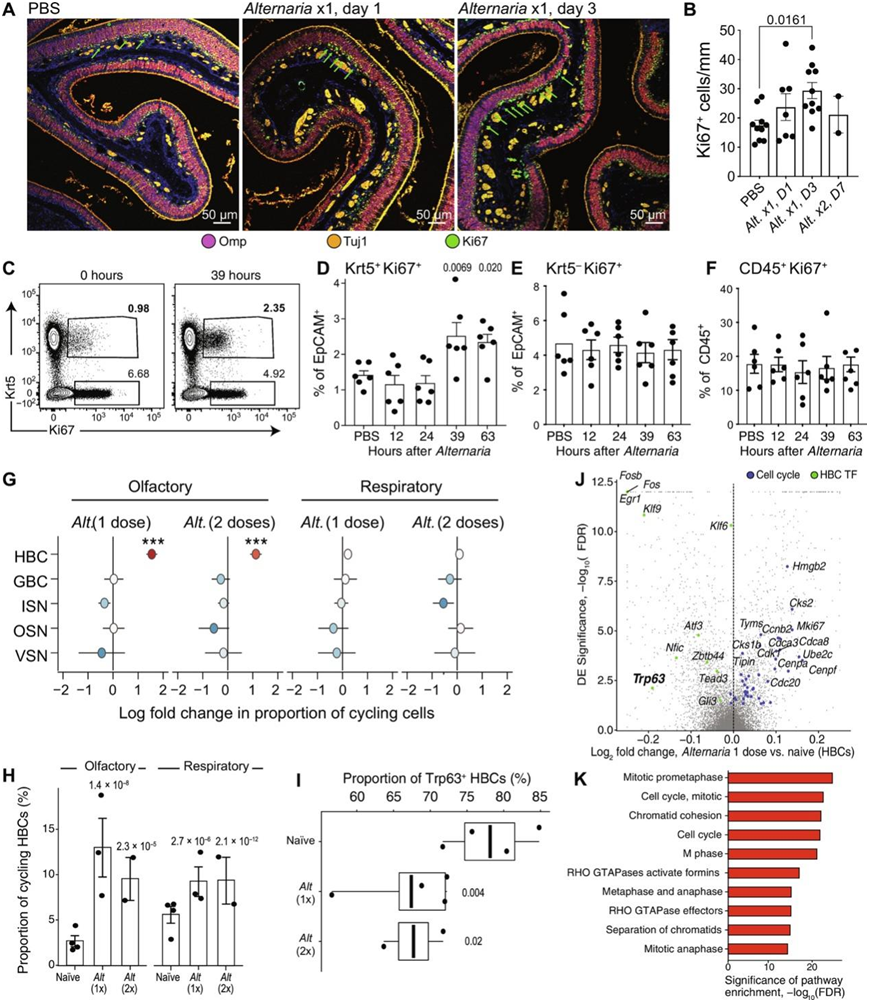
图5. 通过吸入过敏原诱导 HBC 增殖。
(A)组织学评估。(B)增殖定量。(C-F)流式细胞分析。 (G-H)根据 scRNAseq 数据鉴定Alternaria诱导的增殖细胞的细胞类型。(I-K)来自scRNAseq数据显示Alternaria对HBC的转录激活。
06
变应原诱导的嗅觉干细胞可塑性
为了确定嗅觉干细胞生态位在过敏原暴露环境下的可塑性,他们对来自嗅觉上皮的上皮细胞和神经细胞进行了“伪时间”分析(图6A)。推断出的细胞状态轨迹(图6A)表明,HBC产生球状基底细胞,基底细胞依次沿主要途径分化为OSN和次要途径分化为mvc。随着神经发生的进展,早期表达显著变化的基因包括典型的HBC标记物,如Krt5和Trp63,而最新阶段的基因包括成熟的OSN标记物,如Omp、编码嗜铬粒蛋白A的Chga和Olfm1,用于olfactomedin 1(图6B)。正如预期的那样,球状基底细胞标记物Ascl1和Kit在中间阶段短暂激活(图6B)。在向OSN发展的早期,他们观察到一个参与细胞骨架调控的基因模块上调,包括stathin -1 (Stmn1)、α-微管蛋白(Tuba1a和Tuba1b)和β-微管蛋白(Tubb5)。经Alternaria吸入特异性诱导的细胞Mki67/Krt5/Stmn1三种标记呈现阳性,但Mki67和Tubb3未显示阳性(图6C)。他们使用免疫荧光验证了stathmin-1标记早期前体的一个子集,并且很少有增殖细胞对ISN标记TUJ1呈阳性,但有些细胞对stathmin-1呈阳性(图6D)。最后,组织学显示,Alternaria诱导了HBC形态的延伸,表明之前发现的活化HBC“从扁平的静态表型到金字塔型的增殖表型的形态转换”(图6D)。接下来,使用基底细胞粘附分子(BCAM)的组合,干细胞的标记物;NCAM1, osn和ISN的标记物和EpCAM,他们通过流式细胞确定了OSN的不同发育阶段。对EpCAMintBCAM+、EpCAMintNCAM1+和EpCAMlowNCAM1+进行了分类,利用组织学标记和bulk RNA-seq转录分析,分别将其鉴定为HBC、ISN和OSN(图6E)。然后,他们分析了在野生型(WT)小鼠中吸入三种不同的过敏原后,每个亚群的转录和组成可塑性:Alternaria、烟曲霉(Af)或Dermatophagoides pteronyssinus (Dp)。从转录的角度来看,单次吸入这些过敏原中的每一种都会引起HBC的显著变化(图6F)。来自Alternaria-小鼠的HBC中上调的基因主要由细胞周期相关转录本组成(图6G)。
然后,他们比较了Alternaria、Af和Dp在HBC中诱导的基因,以确定常见的转录程序和每种过敏原的特异性转录程序(图6H)。所有三种过敏原诱导的转录本包括单核细胞趋化剂Cxcl17和抗菌肽Lcn2和Ltf,已知它们在过敏性炎症的情况下在上皮中增加。Alternaria对HBC增殖的诱导最为强烈,干细胞激活标记物Mki67、Top2a和Stmn1均特异性上调。此外,Alternaria特异性诱导趋化因子Ccl9、细胞因子Il33和Il4ra,与强烈的2型反应一致(图6G-H)。与Alternaria一样,Af引起HBC细胞周期标记物的表达增加,包括Pcna、Mcm3和Mcm5以及2型相关转录本Chil4和Alox12e,以及嗅觉神经再生相关转录本Sprr2a1和Sprr2a2,但不如Alternaria强烈(图6H)。吸入Dp与强干扰素反应和诱导核因子κB (NFκB)信号传导有关,但没有Alternaria诱导的强增殖反应(图6H)。
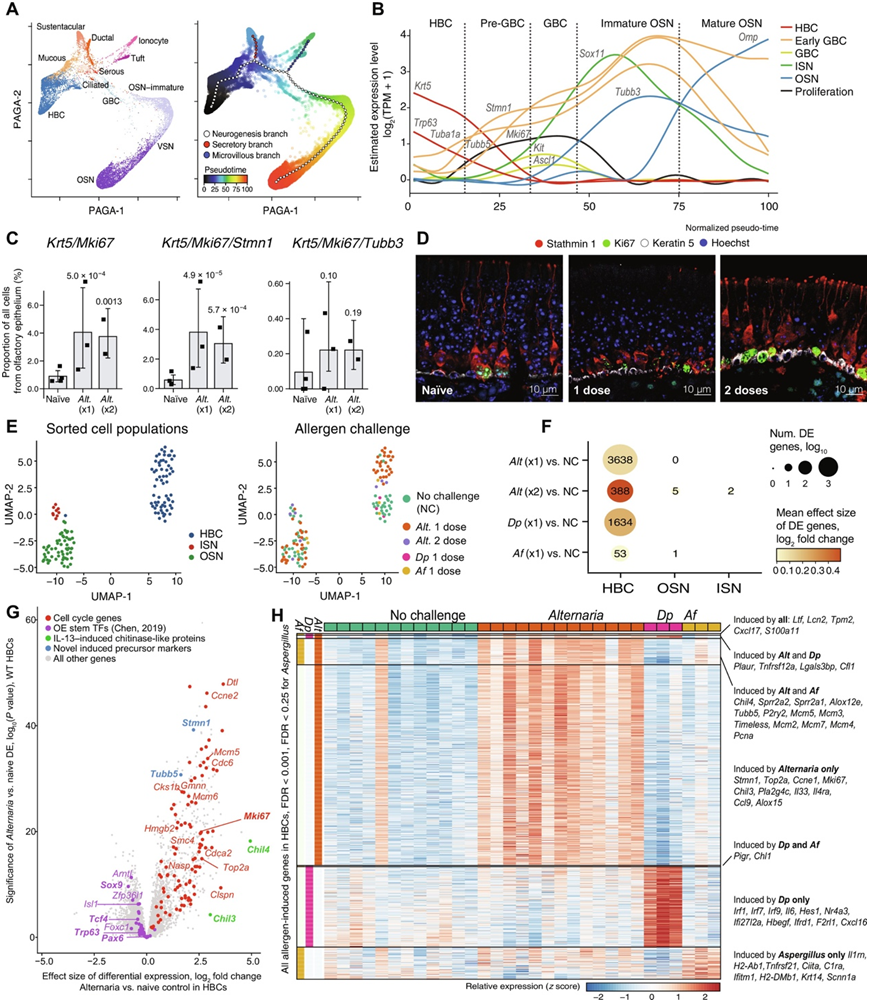
图6. 霉菌和屋尘螨过敏原直接不同的嗅觉干细胞激活程序。
(A) 17318个嗅觉上皮细胞(点)的PAGA可视化,按细胞类型(左)和伪时间着色(右)。(B)经典标记表达水平分析。(C) (C)每只小鼠中每组基因由scRNAseq检测到的细胞平均比例(点)。(D)免疫荧光分析。(E-H)给WT小鼠单次吸入不同过敏原的转录组分析。
07
过敏原诱导的干细胞增殖依赖于簇状细胞
接下来,他们试图确定过敏原诱导干细胞活化的机制。在嗅神经上皮中,干细胞位于几层OSN下,位于腔上皮下60 ~ 100 μm,由TRPM5+簇状mvc、支撑细胞和TRPM5 – mvc(离子细胞)组成。由于簇状mvc直接由Alternaria激活,并且它们位于嗅觉上皮的顶端,与气流相互作用,他们假设簇状mvc介导了过敏原和嗅觉干细胞之间的联系。为了验证这一点,他们评估了缺乏簇状mvc的Pou2f3−/−小鼠对单次吸入Alternaria的反应(图7A)。在簇状细胞缺陷的Pou2f3−/−小鼠中,Alternaria诱导的干细胞增殖减少(图7B-C)。无论是吸入过敏原还是簇状细胞缺乏,Krt5−上皮细胞的增殖、处于稳态或吸入过敏原后的Krt5+干细胞的数量都没有变化。Alternaria诱导的鼻嗜酸性粒细胞和免疫细胞募集和增殖在Pou2f3−/−小鼠中部分减少(图7D)。出乎意料的是,他们发现Alternaria诱导的Krt5+干细胞的增殖在Il7r−/−或Stat6−/−小鼠中都得以保留,这表明先天淋巴细胞和IL-13信号传导并不需要鼻腔干细胞增殖。与先前在肺部的研究结果一致,在Il7r−/−和Stat6−/−小鼠中,Alternaria引起的嗜酸性粒细胞募集、鼻腔炎症和免疫细胞增殖均发生消除,Il7r−/−小鼠的淋巴细胞明显减少。与流式细胞数据一致,分选BCAM+干细胞的RNA-seq表明,Mki67和增殖基因标记在WT、Stat6−/−和Il7r−/−小鼠中均上调,而在Pou2f3−/−小鼠中下调(图7G-H)。依赖于IL-13信号和先天淋巴细胞的转录程序,如Chil4和相关的几丁质酶Chil3,在Stat6−/−中完全消除,在Il7r−/−中强烈减少,而在Pou2f3−/−小鼠中仅受到轻微影响(图7I)。综上所述,这些数据表明鼻丛细胞可以独立于先天免疫细胞和IL-13信号传导,直接诱导过敏原诱导的干细胞增殖。
最后,他们检测了簇状细胞激活是否足以引起干细胞增殖。给小鼠单次吸入5 ' -三磷酸腺苷(ATP)、地那铵或半胱氨酸蛋白酶木瓜蛋白酶,并评估干细胞增殖(图7J)。ATP诱导未分离的鼻腔ChAT+簇状细胞的ChAT+ MVC和CysLT中的钙通量,其中约95%为簇状MVC。地那铵是一种Tas2r配体,可触发ChAT+ MVC亚群中的钙通量。两种潜在的木瓜蛋白酶受体候选物(F2r和F2rl1)均由簇状scc和簇状mvc表达。吸入地那铵和木瓜蛋白酶诱导Ki67+/Krt5+细胞百分比适度增加,Ki67+/Krt5+细胞数量变化更明显(图7K)。ATP诱导干细胞最强劲的增殖,与Alternaria相当(图7K)。在Pou2f3−/−小鼠中,atp诱导的增殖干细胞的绝对数量减少(图7K),表明簇状细胞激活也足以诱导干细胞增殖。
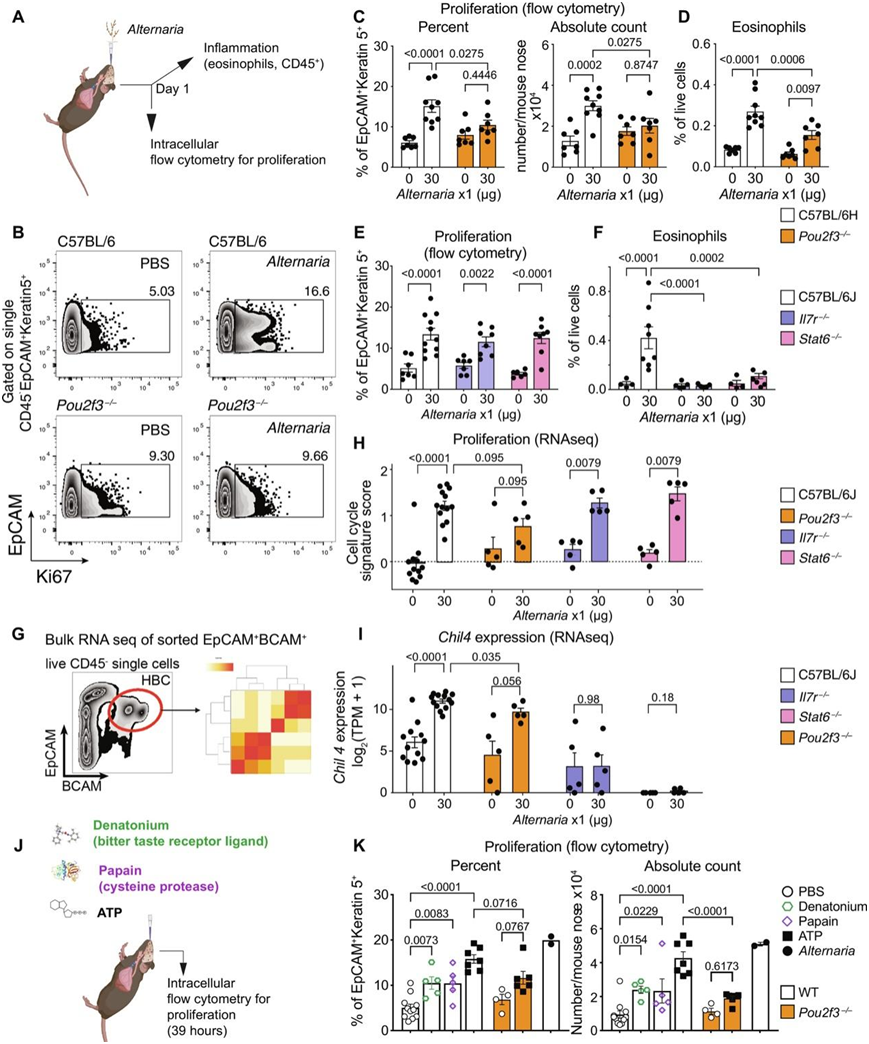
图7. 过敏原诱导的干细胞增殖不依赖于 2 型炎症途径,而是依赖于簇状细胞。
(A-I) 给指定基因型的小鼠鼻内给予单剂量Alternaria,评估鼻黏膜的干细胞增殖情况。(J-K) 流式细胞分析对Ki67核内染色评估Krt5细胞的增殖百分比和数量。
+ + + + + + + + + + +
结 论
本项研究使用综合单细胞转录组分析的方法,发现吸入Alternaria导致黏膜上皮效应分子的表达,包括Chil4,以及引发HBC的增殖的独特途径,这两者都在嗅觉细胞凋亡缺失的情况下触发。Alternaria和ATP诱导的HBC增殖依赖于TRPM5+簇状MVC,确定了这些特殊的上皮细胞作为调节嗅觉干细胞响应的调节器。综合而言,本项研究的数据提供了鼻簇状细胞异质性的高分辨率表征,并确定了TRPM5+簇状MVC在引导嗅觉黏膜对过敏原的反应中的功能。
+ + + + +



