

 English
English文献解读|J Hepatol(25.7):FASN介导的脂肪酸生物合成重塑华支睾吸虫感染相关肝内胆管癌的免疫环境
✦ +
+
论文ID
原名:FASN-mediated fatty acid biosynthesis remodels immune environment in Clonorchis sinensis infection-related intrahepatic cholangiocarcinoma
译名:FASN介导的脂肪酸生物合成重塑华支睾吸虫感染相关肝内胆管癌的免疫环境
期刊:Journal of Hepatology
影响因子:25.7
发表时间:2024.03.19
DOI号:10.1016/j.jhep.2024.03.016
背 景
肝内胆管癌(ICC)是第二常见的原发性肝癌,具有高致死率。华支睾吸虫(C. sinensis)感染是ICC的重要危险因素。华支睾吸虫产生的排泄/分泌产物 (ESP)在宿主-寄生虫相互作用中发挥重要作用,从而促进 ICC 肿瘤发生。从机制上讲,ESP 可能通过调节组蛋白乙酰转移酶诱发胆管癌,并参与晚期导管周围纤维化。然而,华支睾吸虫感染对 ICC 患者预后的临床影响以及潜在的分子特征仍不清楚。
实验设计
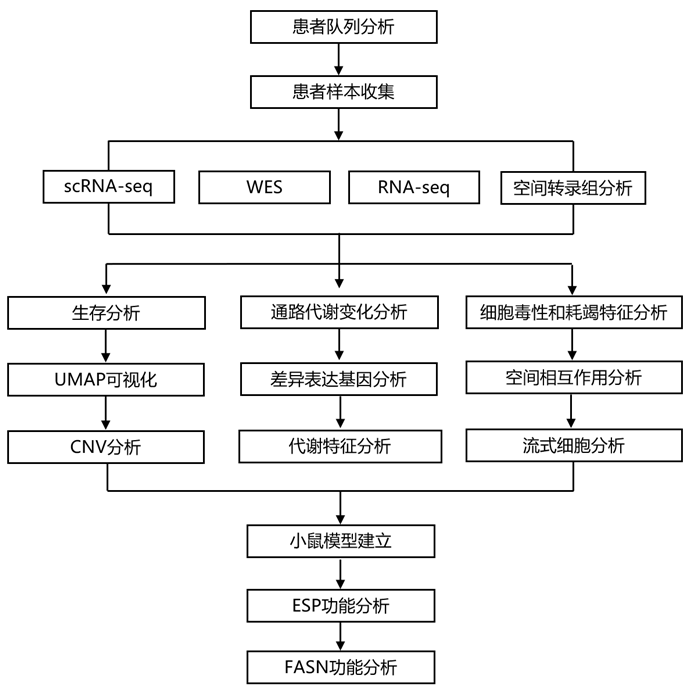
结 果
01

华支睾吸虫感染的 ICC 患者预后较差,对免疫检查点阻断的反应较差
为了评估华支睾吸虫感染对ICC患者预后的临床影响,研究团队纳入了来自三个医疗中心的171例接受根治性手术的原发性ICC患者和47例接受抗pd -1定向免疫治疗的晚期ICC患者。对于接受根治性手术的原发性 ICC 患者,华支睾吸虫感染患者的中位总生存期 (OS) 为 15.5 个月,而非华支睾吸虫感染患者为 25.6 个月(图1A,图S1A)。他们还研究了晚期ICC患者对抗pd -1/PD-L1定向免疫治疗的反应,并观察到华支睾吸虫感染患者的中位无进展生存期(PFS)明显短于未感染的患者(图S1B)。
为了分析华支睾吸虫感染的 ICC的肿瘤微环境 (TME) 和分子特征,他们对 13 名未接受治疗的 ICC 患者进行了单细胞转录组分析(scRNA-seq),这些患者通过手术切除获得了 15 个肿瘤样本,其中包括 8 个华支睾吸虫感染的ICC 。同时,对204例患者进行了全外显子组测序(WES)、bulk RNA测序(bulk RNA-seq)以及空间转录组学分析(图1B-C、图S1A-B)。推断拷贝数变异(CNV)以验证恶性细胞的定义(图1D)。一些众所周知的癌基因,例如MYC和ERBB2发生扩增,这与WES中的CNV结果一致(图S1D)。差异表达基因(DEG)和标记基因显示出具有不同分子特征的异质免疫细胞亚型,证实了细胞身份的准确性(图1E,图S1E)。免疫细胞亚型在华支睾吸虫感染非华支睾吸虫感染之间表现出不同的分布。在华支睾吸虫感染的ICC样本中,CD8_C9_MKI67、CD8_C2_LAG3、Mac_C10_CXCL10、Mac_C8_MKI67、Mac_C0_SPP1等免疫细胞显著富集,构成了华支睾吸虫感染ICC的独特微环境景观(图1F)。
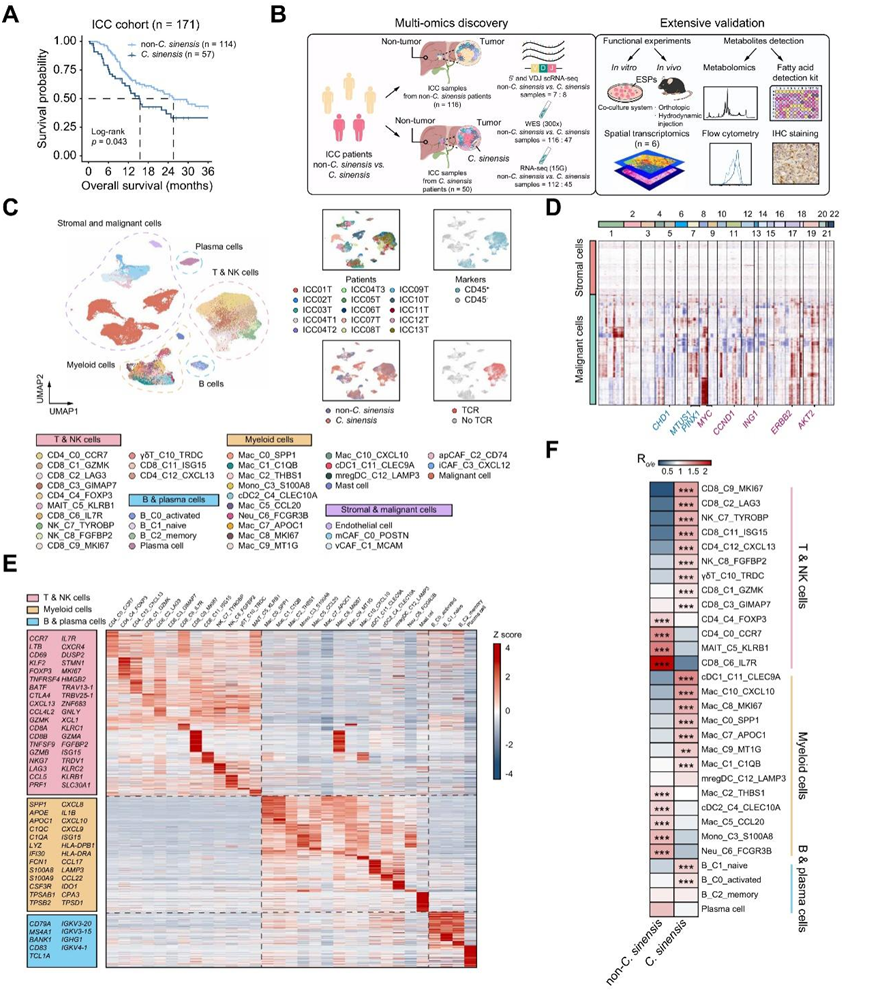
图1. 通过 scRNA-seq 表征感染华支睾吸虫的 ICC 中的 TME。
(A) Kaplan-Meier 分析显示有或没有华支睾吸虫感染的患者的总体生存率。(B) 研究设计的工作流程。(C) 左图中免疫细胞、基质细胞和恶性细胞聚类的统一流形近似和投影 (UMAP) 可视化,右图中相应的患者来源、用于区分免疫细胞的标记、感染状态和 TCR 检测。(D) inferCNV 推断的恶性细胞的拷贝数变化。(F)每个免疫细胞聚类在华支睾吸虫和非华支睾吸虫感染的ICC之间的分布差异。
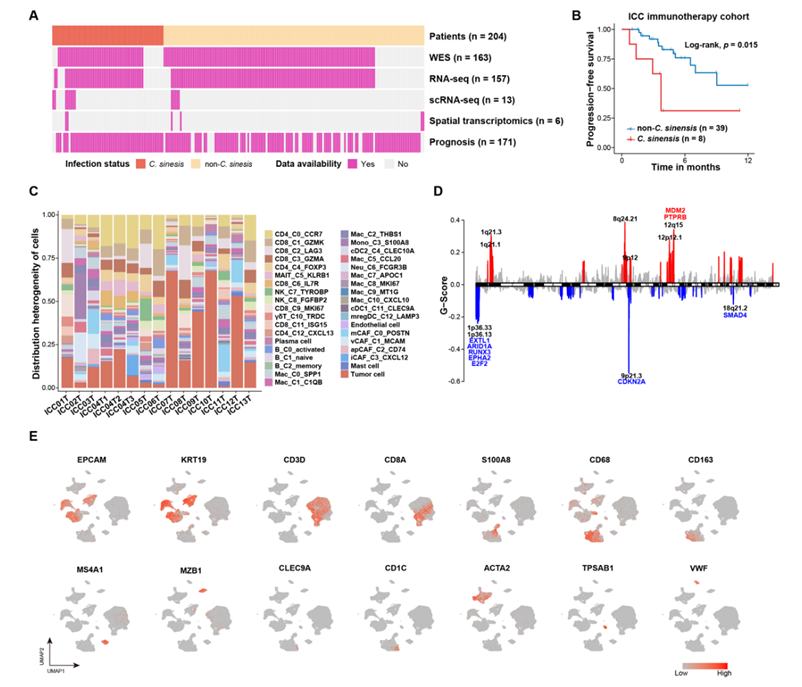
图S1. 华支睾吸虫感染的ICC与非华支睾吸虫感染的ICC的研究设计与转录组学特征。
(A)纳入多组学研究的样本。(B) Kaplan-Meier分析显示,接受免疫治疗的患者有无华支睾吸虫感染的无进展生存期。(C)每个患者的细胞组成图。(D) WES数据的拷贝数概况。(E)显示特定细胞类型表达的典型基因的UMAP。
02
来自华支睾吸虫感染的 ICC 的恶性肿瘤细胞表现出增强的代谢和细胞周期基因表达特征
为了表征华支睾吸虫感染的 ICC的基因组图谱,他们对 163 名初治 ICC 患者的肿瘤和配对非肿瘤肝组织进行了 WES(图 2A,S1A)。在华支睾吸虫感染的ICC与非华支睾吸虫感染的ICC之间反复突变基因没有显著差异(图2A,图S2A)。 与非华支睾吸虫感染的 ICC 相比,在华支睾吸虫感染的 ICC中观察到显著更高的肿瘤突变负荷(TMB),这意味着华支睾吸虫的存在可能伴随着肿瘤遗传不稳定性的增加(图2A)。
华支睾吸虫感染的ICC恶性细胞中有209个基因显著上调,包括代谢相关基因PLTP、肿瘤生长相关基因MET和SERPINE2,以及干细胞相关基因CD44(图S2C)。他们还对恶性细胞进行了共识 NMF (cNMF) 分析,并确定了 ICC 的五个功能元程序 (MP)(图 2B)。
MP2、MP3 和 MP5 中的大多数细胞来自华支睾吸虫感染的 ICC,而 MP4 中的大多数细胞来自非华支睾吸虫感染的 ICC(图 2 C)。细胞周期相关基因(如TOP2A、CDC20和MKI67)、免疫相关基因(如C1QA、CD74和CXCL13)和脂肪酸代谢相关基因(如FABP1)分别在MP2、MP3和MP5中上调(图 2B-C)。MYC、干扰素γ反应和脂肪酸代谢通路在MP2、MP3和MP5中分别显著富集(图 2B-C)。脂肪酸生物合成通路在华支睾吸虫感染的icc恶性细胞中显著富集(图 2D)。
为了探索华支睾吸虫感染的icc中参与脂肪酸生物合成通路的关键基因,他们检测了相关酶的表达水平,发现脂肪酸合酶(FASN)在华支睾吸虫感染的icc中显著上调,免疫组织化学(IHC)染色证实了这一点(图2F)。FASN是脂肪酸生物合成的关键调控因子,在肿瘤的生长和预后中起关键作用。因此,根据FASN的中位表达强度,他们将143例ICC患者分为FASNhigh和FASNlow组,FASNhigh组患者的OS明显短于FASNlow组(图2G)。
已有研究表明,华支睾吸虫分泌的ESP在宿主-寄生虫相互作用中起关键作用,因此,他们在体外用ESP处理ICC细胞和非恶性胆管细胞,模拟华支睾吸虫感染。结果显示,ESP处理显著提高FASN表达(图2H-I)。更有趣的是,代谢检测试剂盒检测到,esp处理细胞的上清液中游离脂肪酸水平显著升高(图2J)。这些结果共同表明华支睾吸虫感染可以促进ICC脂肪酸生物合成。

图2. 源自华支睾吸虫感染的 ICC 的恶性细胞的分子特征。
(A) 所有 163 名 ICC 患者的基因谱和相关临床特征。不同的改变类型和临床特征用不同的颜色代码表示。(B) 热图显示了通过一致非负矩阵分解 (cNMF) 识别的差异表达基因(行,左图),并按单细胞表达模式聚类为 5 个 MP(列,左图)。每个 MP 相应的富集通路列在右图中。(C)不同感染状态的程序的组成变化。 (D) 火山图显示华支睾吸虫 ICC 和非华支睾吸虫感染的 ICC 之间恶性细胞的代谢活性存在差异。(E) 华支睾吸虫感染的 ICC恶性细胞中 FASN 的表达水平上调。 (F) FFPE 组织中 IHC 染色的代表性图像(左)以及华支睾吸虫 ICC 和非华支睾吸虫感染的 ICC 中 FASN 蛋白表达(右)。 (G) Kaplan-Meier 分析显示根据 FASN 蛋白表达从 (E) 分为 FASN高和 FASN低ICC 患者的总体生存率。(H–I) 与对照组相比,ESP 处理的 ICC 细胞系中 FASN 表达水平的 RT-qPCR 和免疫印迹分析。(J) 使用代谢检测试剂盒对 ESP 处理的 ICC 细胞系的条件培养基与对照组进行游离脂肪酸检测。
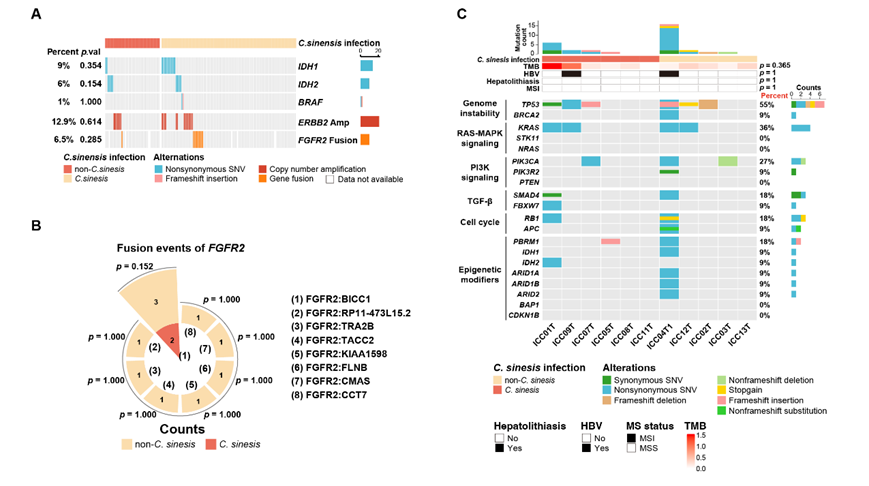
图S2. WES数据分析。
(A)所有163例ICC患者临床批准的可药物靶向基因改变。(B) FGFR2的融合事件。(C)与scRNA-seq数据配对的患者的遗传谱和相关病因。
03
来自华支睾吸虫感染的 ICC 的巨噬细胞表现出肿瘤相关巨噬细胞(TAM)样表型
接下来,他们试图探究ICC患者免疫细胞标记物与代谢相关基因的相关性。他们鉴定了众多免疫细胞,包括Mac_C8_MKI67、CD8_C9_MKI67、Mac_C0_SPP1、CD4_C12_CXCL13、Mac_C1_C1QB和CD8_C2_LAG3,并且它们与恶性细胞的代谢特征密切相关(图3A),表明这些免疫细胞可能受到代谢活动扰动的影响来自华支睾吸虫感染的 ICC的恶性细胞。
Mac_C0_SPP1、Mac_C1_C1QB、Mac_C7_APOC1、Mac_C8_MKI67、Mac_C9_MT1G 和 Mac_C10_CXCL10 在华支睾吸虫感染的ICC中富集,而Mac_C2_THBS1 和 Mac_C5_CCL20 在非华支睾吸虫感染的 ICC中富集(图 1 F)。与募集免疫调节细胞(例如骨髓源性抑制细胞)和促进癌症进展有关的趋化因子(包括CCL2、CCL3和CCL4)也在华支睾吸虫感染的 ICC的巨噬细胞中富集。一组高表达IL-1B和TNF且低表达TAM样基因的巨噬细胞是非TAM样巨噬细胞,其表现出促炎表型(图3B)。因此,Mac_C0_SPP1和Mac_C1_C1QB属于TAM样巨噬细胞,而其他细胞是非TAM样巨噬细胞。为了进一步推断巨噬细胞的动态免疫状态转变,他们使用Monocle3构建了单核细胞和巨噬细胞的发育轨迹(图3D)。结果表明,TAM 样和非 TAM 样巨噬细胞发育成不同的轨迹(图 3 D-E)。SPP1、APOE、C1QB和CD163随着时间的推移逐渐升高,并在轨迹末端达到峰值,伴随着TAM样巨噬细胞的积累(图3E -F)。

图3. 来自华支睾吸虫感染的 ICC的恶性细胞的代谢特征与巨噬细胞极化为 TAM 样细胞有关。
(A) 华支睾吸虫感染的 ICC中代谢相关基因的表达水平上调,在 RNA-seq 数据中显示不同免疫细胞亚群的高水平浸润组。(B) 代表巨噬细胞表型状态的选定功能细胞标记的热图。左图可视化了在非华支睾吸虫感染的ICC中富集的巨噬细胞中所选标记的表达状态,并且右图代表在华支睾吸虫感染的ICC中富集的巨噬细胞中的相同表达。(C)差异表达的巨噬细胞TAM样基因。(D) 按 monocle3 计算的伪时间排序的单细胞轨迹。 (E) 密度图显示巨噬细胞沿伪时间的分布(上图)。显示了指示基因表达沿伪时间动态变化的热图(下图)。(F) 曲线图显示了巨噬细胞转变过程中选定基因的动态表达以及伪时间。
04
来自华支睾吸虫感染的 ICC 的T 细胞具有细胞毒性和耗竭特征
鉴于华支睾吸虫感染的 ICC中 TAM 样特征的显著富集,他们使用 CellChat 进一步计算了 TAM 样巨噬细胞与其他细胞之间的细胞间通讯概率。结果显示 TAM 样巨噬细胞和 CD8 + T 细胞之间的潜在相互作用最高(图 4 A)。因此,他们将T淋巴细胞重新聚类成13个亚聚类(图4B ),在非华支睾吸虫和华支睾吸虫感染中的ICC之间分布不同(图4B)。他们观察到CD8+ T 细胞的高度多样性(图 4 C)。CD8_C1_GZMK高水平表达GZMK、NKG7和IFNG等效应标记物的细胞定义为效应T细胞。CD8_C2_LAG3的特征是耗尽标记物(如LAG3、TIGIT和PDCD1)和效应标记物(如NKG7、GNLY和IFNG)共同表达,它们分别指定为功能障碍前T细胞。CD8_C3_GIMAP7 和 CD8_C6_IL7R 聚类均具有CCR7、SELL和TCF7等幼稚标记基因的表达增加,识别为幼稚 CD8+和记忆 CD8+ T 细胞。CD8_C9_MKI67定义为增殖性的并且以MKI67的表达为特征。CD8_C11_ISG15以CD69、NR4A1和ZNF683为特征,他们将其鉴定为组织驻留记忆T细胞(TRM)。他们观察到CD8_C2_LAG3和CD8_C9_MKI67具有高细胞毒性和耗竭评分,而CD8_C3_GIMAP7和CD8_C6_IL7R具有低细胞毒性和耗竭评分(图4D)。总体而言,华支睾吸虫感染的患者的CD8+ T细胞的细胞毒性和耗竭评分明显高于非华支睾吸虫感染患者。
接下来,他们研究了tam样巨噬细胞(Mac_C0_SPP1和Mac_C1_C1QB)和CD8+ T细胞之间的相互作用。值得注意的是,通过LGALS9/HAVCR2和PDCD1LG2/PDCD1,在tam样巨噬细胞和CD8+ T细胞之间发现了高强度的配体受体(LR)评分,而这两种评分仅存在于华支睾吸虫感染的ICC中(图4E)。LGALS9(编码半乳糖凝集素 9)可以通过与HAVCR2(称为 TIM-3)结合促进效应 T 细胞凋亡。PDCD1LG2 (编码 PD-L2)是PDCD1 (称为 PD-1)的配体,在多种肿瘤中表达,可抑制抗肿瘤免疫。
他们评估了每位患者的 43 种细胞因子活性,并计算了细胞因子活性与 T 细胞增殖之间的中值相关性差异(图 4 F)。有趣的是,一些已知的免疫抑制因子,如FGF2、IL-6、TGFB3、IL-10和LIF,在华支睾吸虫感染的ICC中发挥了更突出的抑制T细胞功能的作用(图4F-G)。而两组间PD-1的表达差异无统计学意义(图4G)。然后他们比较了不同细胞类型中细胞因子表达与相应细胞因子诱导的CD8+ T细胞免疫抑制水平之间的相关性。结果表明,华支睾吸虫感染的ICC中的巨噬细胞可能是参与抑制CD8+ T功能的主要细胞类型(图4H)。总而言之,华支睾吸虫感染的 TME 中的 TAM 样巨噬细胞可能通过免疫检查点分子和华支睾吸虫感染的 ICC中的分泌性免疫抑制细胞因子与效应 CD8 + T 细胞相互作用,从而促进免疫抑制微环境。
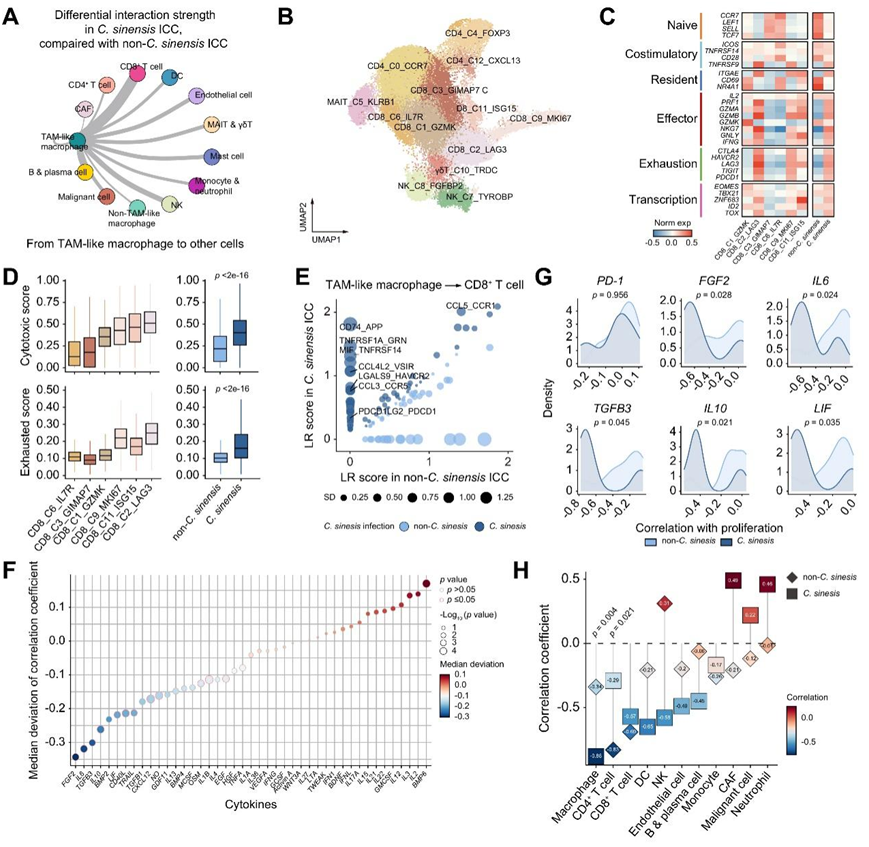
图4. 华支睾吸虫感染的 ICC 中CD8+ T 细胞的转录组特征和潜在的抑制信号。
(A)通过 CellChat 计算,与未感染华支睾吸虫感染的 ICC 相比,TAM 样巨噬细胞与其他细胞亚型之间的差异相互作用强度。(B) T 细胞聚类的 UMAP 可视化。(C) 代表 CD8+ T细胞表型状态的所选细胞标记物的热图。左图可视化CD8+ T细胞聚类中的表达状态,右图代表华支睾吸虫和非华支睾吸虫感染的ICC之间的相同表达。(D) 不同 CD8 + T 细胞组的细胞毒性和耗竭评分(左图),以及按感染状态分组的 CD8 + T 细胞(右图) (E) TAM 样巨噬细胞和 CD8+ T 细胞之间的 LR(配体受体)评分,按不同感染状态着色。点大小代表华支睾吸虫和未感染华支睾吸虫感染的ICC之间LR分数的差异倍数。(F) 气泡图,显示CD8+ T 细胞中增殖水平与 CytoSig 预测的细胞因子信号传导活性之间的相关系数的中值偏差。(G) 密度图显示特定因素与华支睾吸虫和非华支睾吸虫感染的 ICC 中 CD8+ T 细胞的增殖水平之间的相关性。(H) 相关性华支睾吸虫和非华支睾吸虫感染的 ICC中各个细胞类型表达的关键特征细胞因子与 CD8+ T 细胞增殖水平的关系。
05
空间转录组学和体外功能实验揭示了恶性细胞、巨噬细胞和 CD8+ T 细胞之间的相互作用
在空间分辨转录景观中,脂肪酸生物合成、TAM、细胞毒性和耗尽特征在华支睾吸虫感染患者中都有明显的特征(图5A -B)。为了深入了解细胞类型的空间相互作用,他们可视化了 TAM 样点和肿瘤样点之间的空间分辨距离,揭示了华支睾吸虫感染的 ICC 中更紧密的空间组织(图 5C)。华支睾吸虫感染的ICC中广泛观察到TAM样点和耗尽样点的物理邻接(图5D),证实了恶性细胞、巨噬细胞和CD8+ T细胞之间潜在的频繁相互作用以及免疫抑制微环境的形成。
为了进一步验证华支睾吸虫感染引起的恶性细胞代谢变化对巨噬细胞和T细胞的影响,他们建立了体外共培养体系(图5E)。分离从健康供体获得的外周血单核细胞并诱导为巨噬细胞,然后与经 ESP 预处理的 ICC 肿瘤细胞共培养。结果显示,与经ESP预处理的肿瘤细胞共培养,巨噬细胞转变为TAM样表型,其特征是CD163、CD206、IL-10和PD-L1表达较高(图5F)。这些结果表明TAM样巨噬细胞强烈抑制华支睾吸虫感染的ICC中的T细胞功能(图5G)。
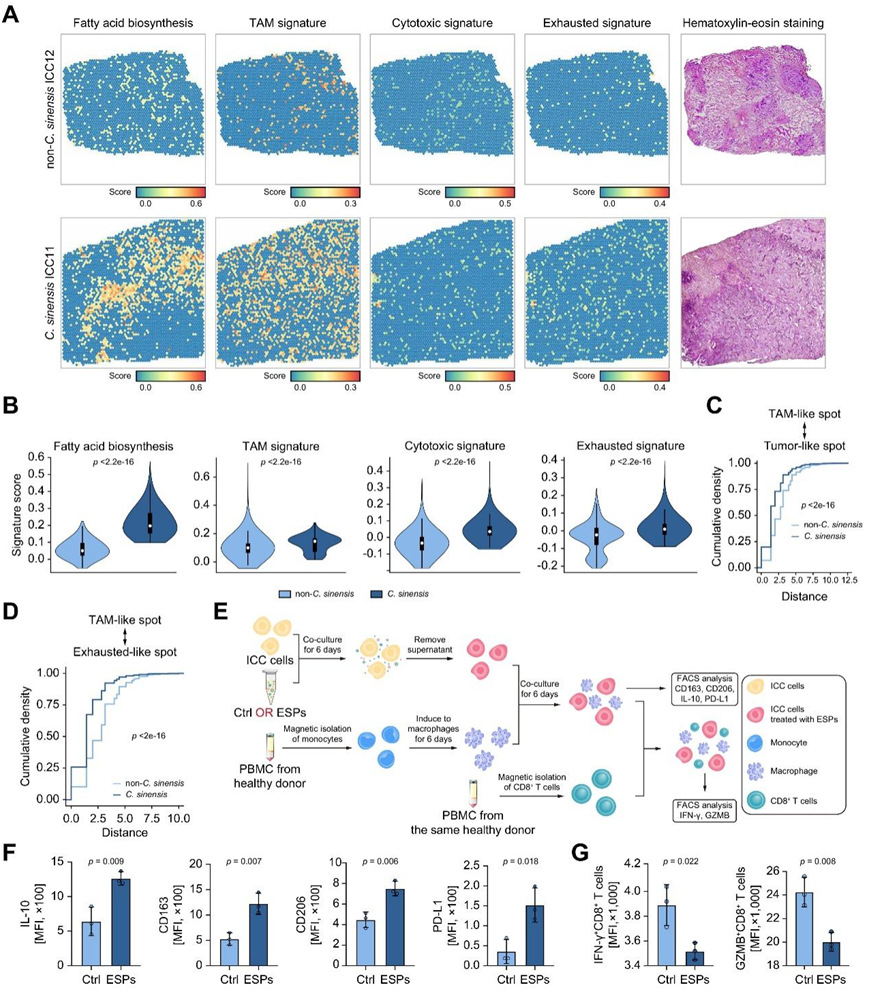
图5. 验证恶性细胞、巨噬细胞和 CD8 + T 细胞的推断特征以及相互作用的关系。
(A) 不同推断特征和相应 H&E 染色的空间特征图。(B) 比较华支睾吸虫和非华支睾吸虫感染的 ICC 患者的推断特征。(C)显示华支睾吸虫和非华支睾吸虫感染的ICC患者中TAM样点和肿瘤样点之间的空间距离的累积概率。(D)显示华支睾吸虫和非华支睾吸虫感染的ICC患者中TAM样点和耗尽样点之间的空间距离的累积概率。(E)共培养系统构建流程图。(F) 与对照组相比,与 ESP 处理的肿瘤细胞 CT1 共培养的人 PBMC 诱导的巨噬细胞的 TAM 样标记物表达的流式细胞分析。(G) 与 (F) 共培养的 T 细胞的 T 细胞功能标志物表达的流式细胞分析。
06
华支睾吸虫来源的 ESP 可通过调节肿瘤代谢和免疫微环境促进肿瘤生长
为了评估华支睾吸虫ESP对肿瘤进展和免疫微环境调节的影响,他们建立了完善的流体动力注射ICC小鼠模型。在该模型中,ESP显著增加了ICC肿瘤负荷,表现为肝重、肝重/体重比和Ki67表达的增加(图6 A-C)。对这些小鼠的肿瘤组织进行了转录组分析(RNA-seq),结果表明,ESP处理可以显著上调FASN表达和脂肪酸生物合成通路(图6D)。与此一致的是,esp处理组FASN蛋白水平明显升高(图6E),同时γ-亚麻酸、顺式-9、12-亚油酸、肉豆酱酸、棕榈酸和硬脂酸等游离脂肪酸显著升高(图6F)。重要的是,在该小鼠模型中,ESP确实可以驱动巨噬细胞极化为tam样亚型并损害T细胞功能(图6G-H)。
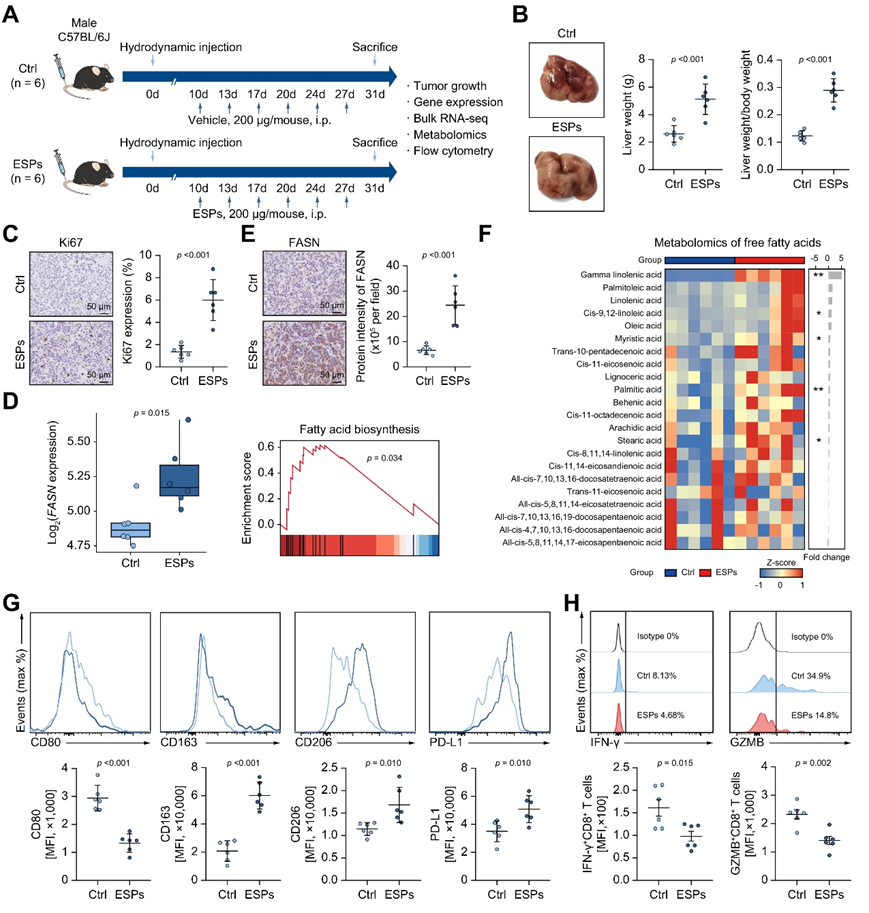
图6. ESP 对肿瘤生长、代谢和体内免疫微环境的调节。
(A) 实验设计。(B) ESP 或对照组的肿瘤、肝脏重量以及肝脏重量与体重的比率的图像。数据显示为平均值±标准差。(C) ESP 或对照组的 Ki67 增殖水平。(D) FASN的转录组差异(左图)和来自 ESP 或对照组的RNA-seq 的脂肪酸代谢通路活性。(E) IHC 染色的代表性图像(左图)和来自 ESP 或对照组的肿瘤组织中 FASN 的蛋白质强度(右图)。(F) 热图显示了通过靶向代谢组学检测到的 ESP 或对照组之间肿瘤组织中游离脂肪酸的成分差异。(G-H) ESP 或对照组之间巨噬细胞 TAM 样和 T 细胞的能标志物表达的流式细胞分析。
07
FASN 抑制剂与 PD-1 阻断剂对 ESP 治疗的 ICC 疗法具有协同作用
接下来,他们采用了水动力注射ICC小鼠模型,其中FASN抑制剂Cerulenin单独给药或与PD-1阻断抗体联合给药于接受或不接受ESP治疗的小鼠(图7A -B)。结果表明,在给予 Cerulenin 后,ESP 处理的肿瘤显著缩小,但对抗 PD-1 阻断表现出轻微反应(图 7 C-D)。此外,在单独使用Cerulenin或与抗PD-1联合治疗时,M1标记物CD80的表达增强,而TAM样标记物如CD206、CD163和PD-L1显著减少(图7E -H)。在 ESP 处理的肿瘤细胞中,T 细胞的细胞毒功能显著受损,单独使用 Cerulenin 可以显著恢复,并且与抗 PD-1 治疗联合使用时进一步增强(图 7 I-J)。这些观察结果共同表明,FASN 抑制剂可以有效抑制巨噬细胞 TAM 样极化,从而恢复细胞毒性 T 细胞的功能,并增强抗 PD-1 治疗在华支睾吸虫感染的 ICC 中的疗效(图 7 K)。
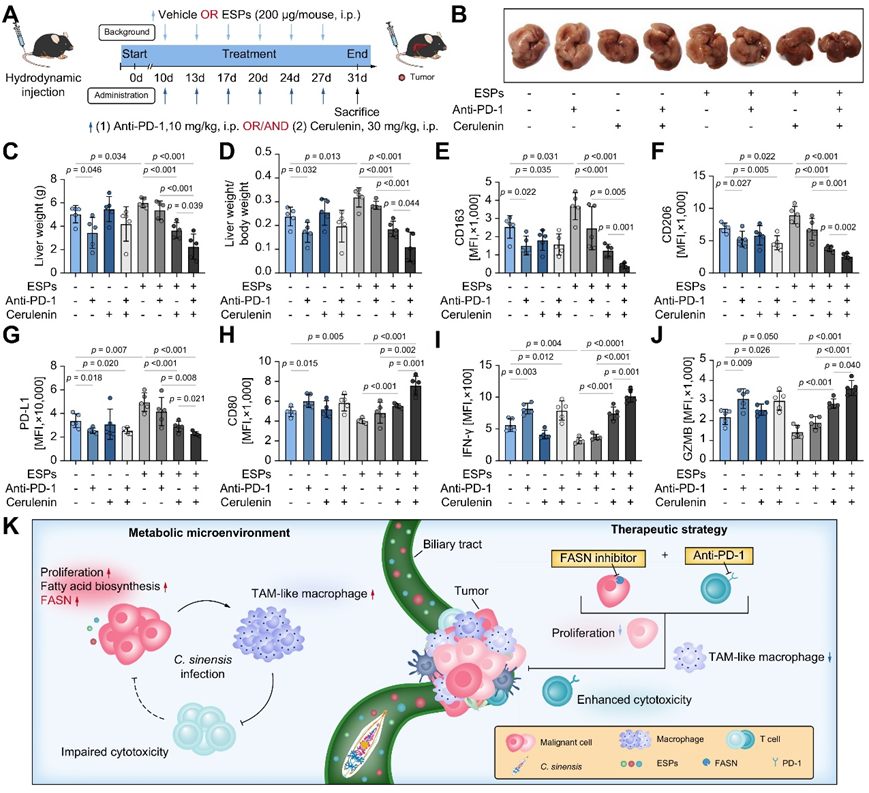
图7. 与 FASN 抑制剂联合使用时,ESP 处理的 ICC 的 PD-1 阻断效果增强。
(A) 实验设计。(B) 八组肝脏的代表性总体图像。(C-D)八组小鼠的肝脏重量以及肝脏重量与体重的比率。(E-J)小鼠肝脏中巨噬细胞的 TAM 样标记和 M1 标记以及 T 细胞的 T 细胞功能标记表达。(K) 图形摘要。
+ + + + + + + + + + +
结 论
本项研究发现华支睾吸虫感染与 ICC 患者的总生存率和免疫治疗反应显著相关。脂肪酸生物合成和FASN(催化长链脂肪酸合成的关键酶)的表达在华支睾吸虫感染的ICC中显著富集。华支睾吸虫的ESP处理的ICC细胞系显示出FASN和游离脂肪酸的升高。肿瘤细胞的代谢改变与肿瘤相关巨噬细胞的富集和T细胞功能的受损密切相关,从而导致免疫抑制微环境的形成和肿瘤的进展。空间转录组学分析显示,与未感染的ICC相比,在华支睾吸虫感染的ICC中,恶性细胞与TAM样巨噬细胞的排列更为紧密。重要的是,在用华支睾吸虫ESP 处理的 ICC 小鼠模型中,FASN 抑制剂显著逆转了免疫抑制微环境并增强了抗 PD-1 功效。
+ + + + +



