

 English
English文献解读|Nat Aging(16.6):年龄、性别和近远端解析的多组学确定了非人类灵长类动物肠道衰老的调节因子
✦ +
+
论文ID
原名:Age-, sex- and proximal–distal-resolved multi-omics identifies regulators of intestinal aging in non-human primates
译名:年龄、性别和近远端解析的多组学确定了非人类灵长类动物肠道衰老的调节因子
期刊:Nature Aging
影响因子:16.6
发表时间:2024.02.06
DOI号:10.1038/s43587-024-00572-9
背 景
肠道疾病的发病率随着年龄的增长而增加,但控制肠道衰老的机制及其与结直肠癌(CRC)等疾病的联系仍然是未知的。
实验设计
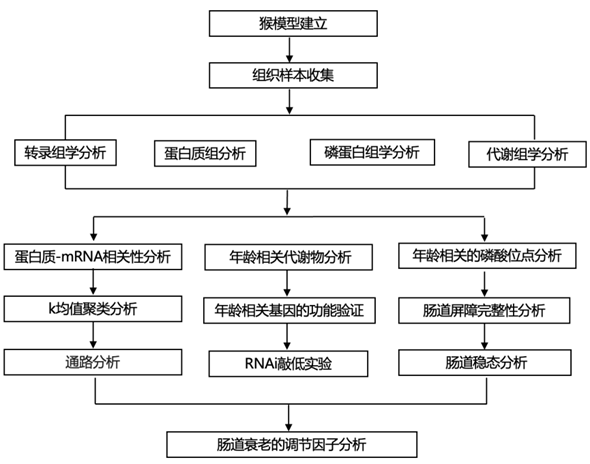
结 果
01

大肠衰老的多组学表征
为了确定大肠衰老的分子景观,研究者团队收集了 26 只束状分枝杆菌动物的 104 块新鲜大肠组织,并进行了年龄解析、位置解析和性别解析的多组学分析,包括转录组学、蛋白质组学、磷酸蛋白质组学和代谢组学(图1a)。在正常大肠组织中观察到蛋白质和mRNA表达之间存在中等相关性(图1b)。并且某些代谢过程,例如缬氨酸、亮氨酸和异亮氨酸降解、脂肪酸降解和色氨酸(Trp)代谢,显示出显著的蛋白-mRNA正相关性(图1c)。
有趣的是,在远端结肠中观察到了更多数量的年龄相关基因产物(图1d)。无监督的k均值聚类分析表明,两个结肠段中的年龄相关蛋白和 mRNA 具有相似的趋势,但变化程度不同(图1e)。一些蛋白质在中年时表现出显著的变化,而在老年时则相反,例如远端结肠中的聚类3和聚类4中的蛋白质(图1e),表明在这一特定生命阶段存在潜在的适应性机制优化功能。血清胆固醇水平往往随着年龄的增长而上升,直到中年,然后下降,并且在肠道胆固醇吸收途径中观察到类似的趋势模式(图1e)。FAO相关转运蛋白和酶,如CPT1A, CPT1B、CPT2、EHHADH和BDH2,在所有年龄阶段的近端结肠中均观察到较高的水平,并且它们的表达随着年龄的增长而快速下降(图1e)。
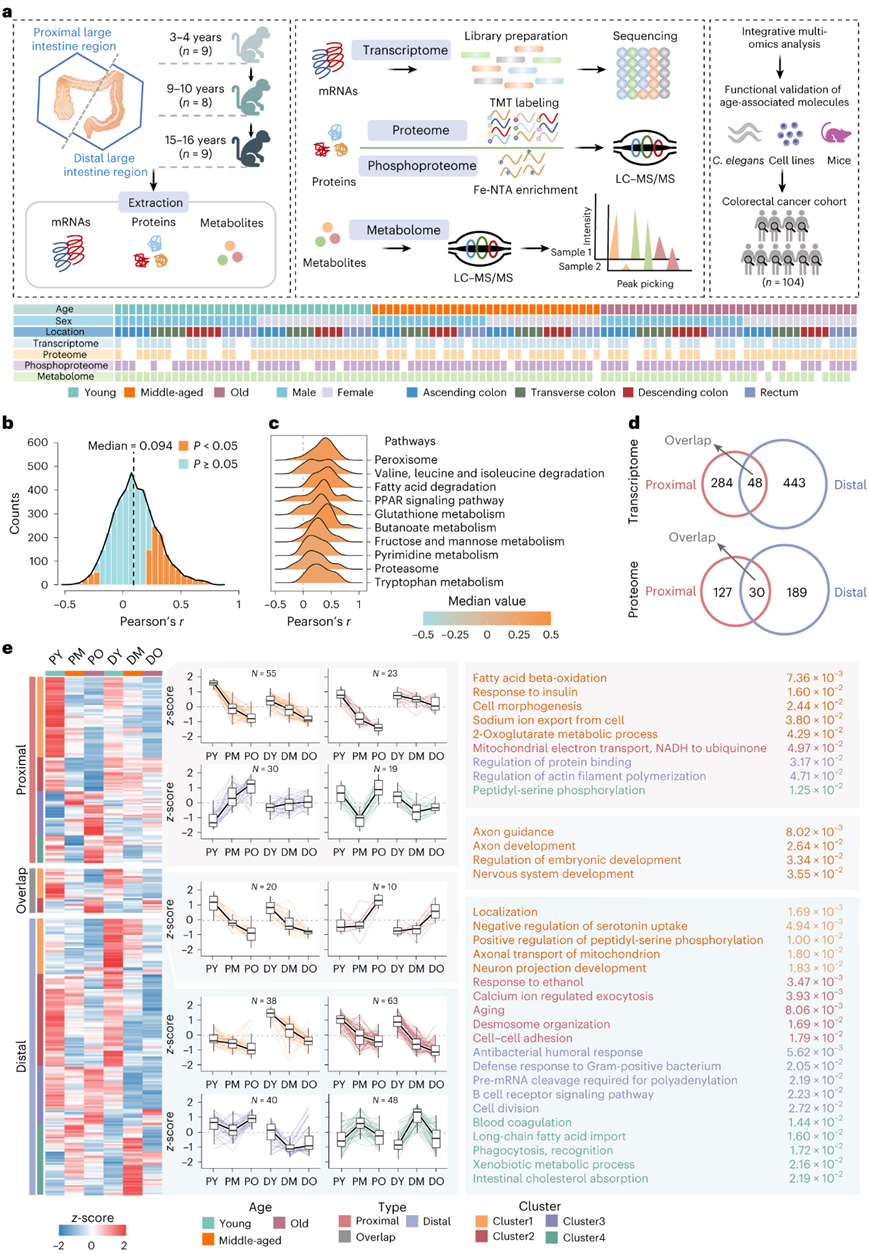
图1. 大肠衰老的位置解析蛋白质组。
(a)动物大肠组织的多组学分析示意图。(b) 蛋白质和 mRNA 之间相关性的直方图。(c) 使用具有 mRNA 与蛋白质正相关性的基因的前 10 个富集 KEGG 通路。(d) 维恩图显示近端和远端结肠之间重叠的年龄相关mRNA/蛋白质。 (e) 年龄相关蛋白的无监督k均值聚类分析。
02
大肠衰老的定位磷酸化蛋白质组分析
蛋白质翻译后修饰 (PTM),例如磷酸化,严格调节衰老过程。引人注目的是,许多磷酸位点在近端和远端结肠中表现出与年龄相关的趋势。在老年组中,他们注意到远端结肠中磷酸位点的减少更多,而近端结肠中磷酸位点的增加数量更多(图2a)。近端和远端结肠中三个年龄阶段的大多数与年龄相关的磷酸位点是不同的(图2b),只有13个位点重叠(图2c)。无监督的k均值聚类分析表明,近端结肠中与年龄相关的磷酸位点在中年时略有减少,随后在老年时增加,而远端结肠中的磷酸位点则随着年龄的增长而一致减少(图2d-f)。带有这些磷酸位点的蛋白质主要与 Rho GTPases 和肌动蛋白丝过程的信号传导相关(图2d-f),表明磷酸化参与细胞-细胞连接调节,强调蛋白质磷酸化和细胞间连接调节之间复杂的相互作用。
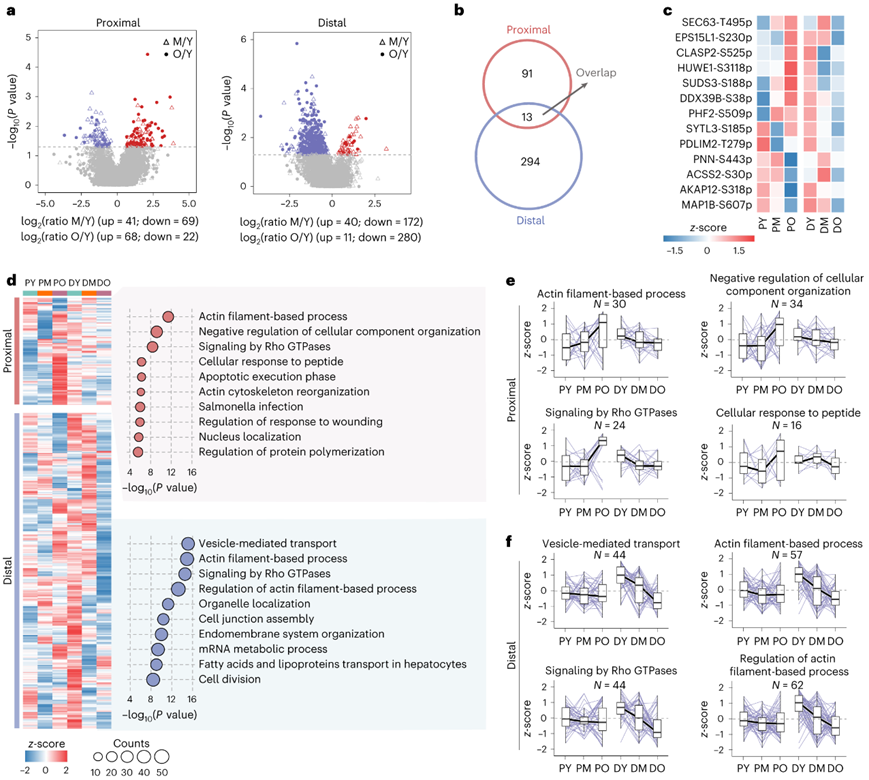
图2. 大肠衰老的定位磷酸化蛋白质组。
(a) 火山图显示近端和远端结肠中中年和年轻个体之间以及老年和年轻个体之间差异表达的磷酸位点。(b) 维恩图显示近端和远端结肠之间重叠的年龄相关磷酸位点。 (c) 热图显示近端和远端结肠中 13 种常见的与年龄相关的磷酸位点的水平。(d)热图显示近端和远端结肠中与年龄相关的磷酸位点的水平。(e-f) 箱线图显示近端和远端结肠中代表性富集途径中磷酸位点的水平。
03
大肠衰老的位置解析和性别解析代谢组学分析
为了深入了解大肠衰老期间的代谢紊乱,他们在近端和远端结肠中鉴定了三个年龄阶段的与年龄相关的代谢物。与转录组学、蛋白质组学和磷酸蛋白质组学的发现一致,远端结肠中出现了更多的与年龄相关的代谢物(图3a)。初级胆汁酸生物合成在远端结肠中表现出与年龄相关的差异(图3a)。他们在近端和远端结肠中发现了性别特异性的年龄相关代谢物(图3b-c)。通路富集分析强调了雄性和雌性近端和远端结肠中与年龄相关的特定代谢途径的不同调节模式(图3d-e)。重要的是,色氨酸代谢仅在雄性远端结肠中富集。色氨酸代谢一般通过犬尿氨酸(KYN)或血清素(5-HT)途径,约90%的色氨酸在大肠中通过KYN途径消耗。令人惊讶的是,他们观察到远端区域的 5-HT 通路活性更高,而近端区域则有利于 KYN 通路(图3f)。
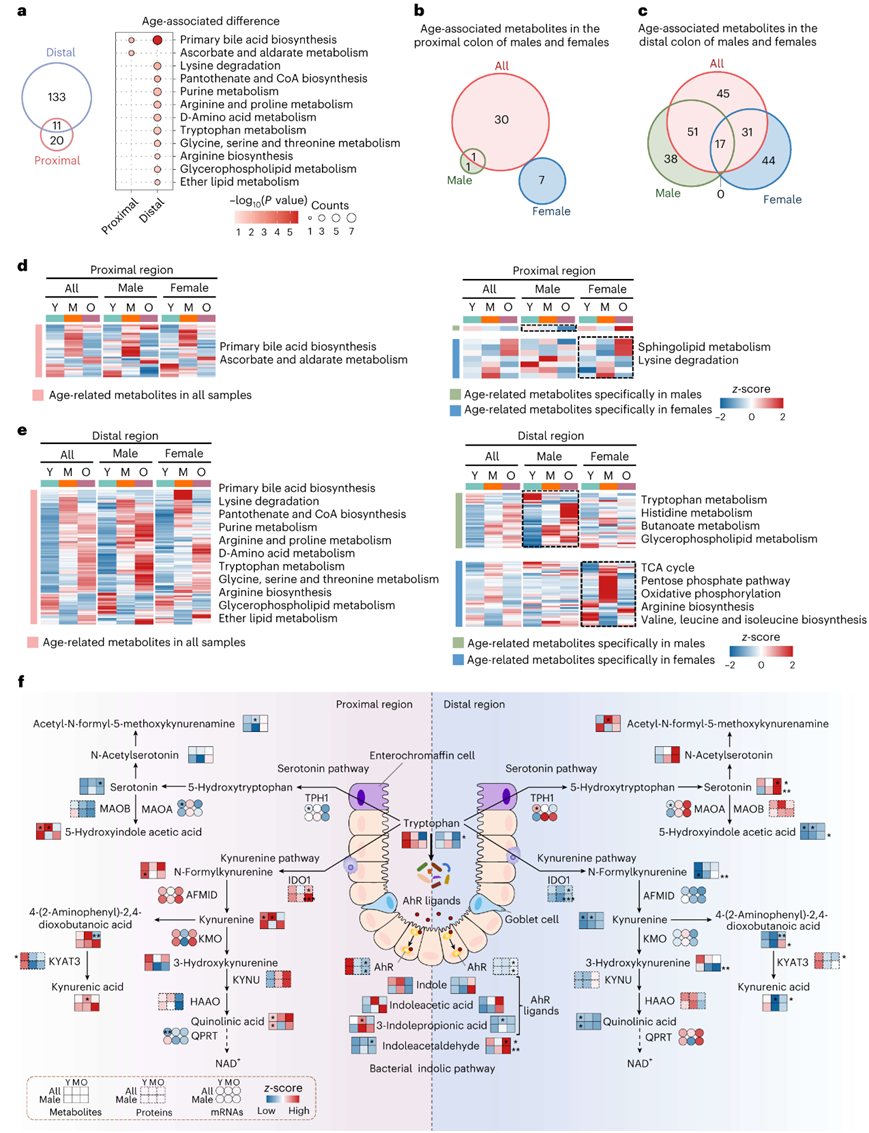
图3. 大肠衰老的位置解析和性别解析代谢组。
(a) 维恩图显示近端和远端结肠中重叠的年龄相关代谢物。 (b-c) 维恩图显示了所有雄性和雌性样本中近端和远端结肠中与年龄相关的代谢物数量。 (d-e) 热图显示每组近端和远端结肠中年龄相关代谢物的表达水平。 (f) 近端和远端结肠中色氨酸代谢途径中的酶和代谢物的水平。
04
年龄相关基因对肠道萎缩的影响
在正常衰老过程中,相对肠道宽度从第 1 天的 0.65 逐渐减小到第 9 天的 0.2。从第3天开始观察到相对肠道宽度明显下降(图4a)。经过适当的筛选后,评估了 37 个年龄相关基因在肠道萎缩中的作用(图4b)。值得注意的是,这些基因在近端和远端结肠中表现出一致的年龄相关变化(图4c)。为了系统地评估它们的影响,用对照或目标基因RNA干扰(RNAi)处理同步线虫,然后在7天后测量管腔宽度(图4d)。在37个基因中,发现23个基因促进肠道衰老,而5个基因阻止肠道衰老(图4d)。在28个有效基因中,11个促衰老基因(ANKRD24、CABP4、DBNL、EPX、GADD45G、GATAD2B、PLA2G2A、HPX、LIPE、SH3BGR和UBAP2)(图4e-f)和4个抗衰老基因(CCT8、HMGCS2、PCK1和SLC9A3)(图4e-g)分别随着年龄的增长而表达增加和减少,表明它们参与了大肠衰老。沉默HMGCS2 (ISC分化的正调节因子)导致管腔宽度变宽(图4e-g)。此外,12个促衰老基因(ANGPTL4、ATP1A3、L1CAM、IYD、NFU1、SCIN、PITX2、RGS13、serpin1、SMDT1、STX1A和CAPN13)随着年龄的增长而减少(图4h),表明在非人灵长类动物中,它们对大肠衰老的贡献减少。值得注意的是,一种抗衰老基因CD82的水平随着年龄的增长而升高(图4i),表明在衰老过程中对肠道萎缩具有潜在的保护作用。
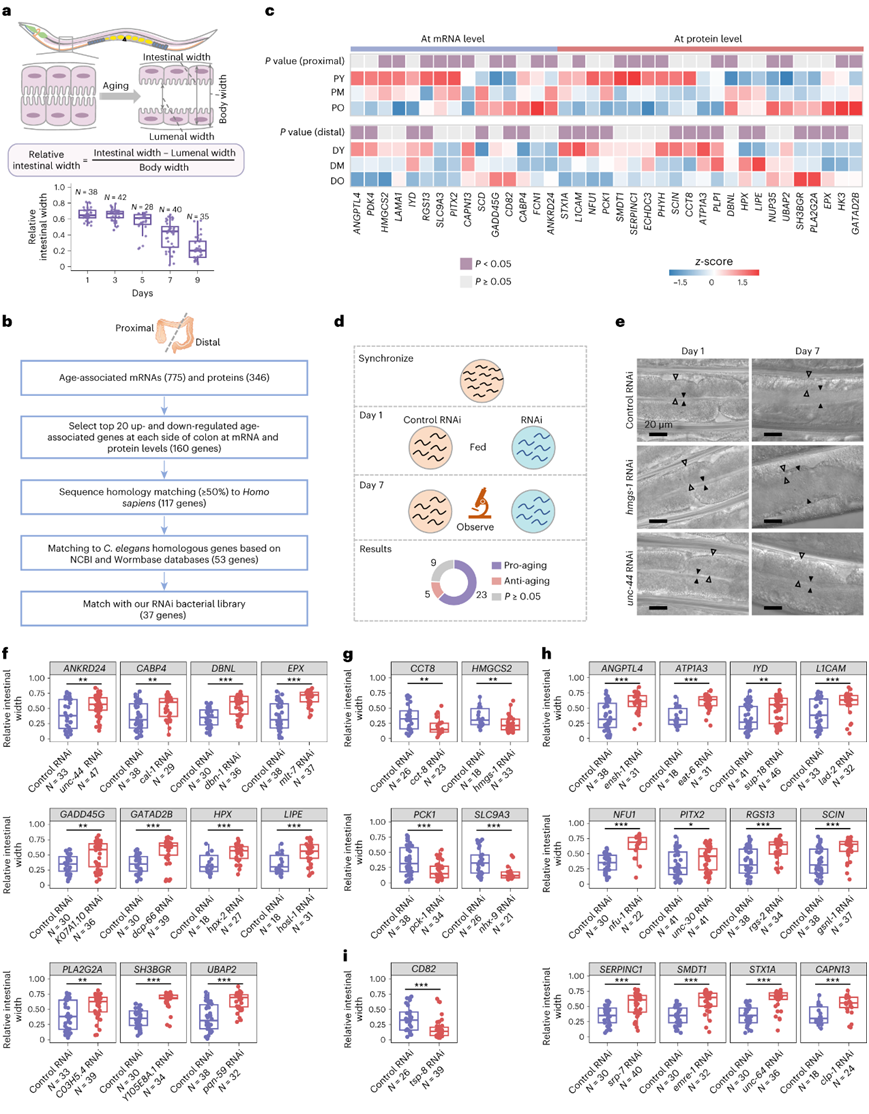
图3. 线虫中年龄相关基因的功能验证。
(a) 肠道萎缩和管腔扩大是线虫肠道衰老的特征。(b) 筛选年龄相关基因进行功能研究的流程图。(c) 热图显示近端和远端结肠中 37 个选定基因的表达。(d) 验证线虫中年龄相关基因的实验流程图。(e) 用RNAi敲低相应基因后的代表性结果。(f-i) 线图描绘了使用 RNAi 敲低目标基因后线虫的相对肠道宽度。
05
年龄相关基因对肠道屏障完整性的影响
有趣的是,在大肠衰老过程中观察到细胞-细胞连接蛋白水平显著下降,细胞-细胞连接蛋白对于维持肠道屏障完整性至关重要。紧密连接(TJ)、粘附连接(AJ)和桥粒蛋白在近端结肠中的表达量高于远端结肠中的表达(图5a-b)。为了确定参与调节肠道屏障完整性的年龄相关基因,他们采用了线虫肠道渗漏模型,随着肠道通透性的恶化,更多的蓝色染料从管腔渗入体腔(图5c)。正如预期的那样,衰老促进了肠道渗漏(图5d-e)。沉默与细胞-细胞连接相关的基因,包括CTNNA1、CTNNB1和DSG2,通过增加蓝色染料的浸润来验证该模型(图5f-g)。接下来,他们研究了可能涉及调节特定细胞-细胞连接蛋白的年龄相关蛋白(图5h)。
他们将人类和秀丽隐杆线虫的同源基因进行匹配,并进一步与RNAi细菌文库进行重叠分析,以鉴定出33个感兴趣的基因(图5h-i)。通过使用RNAi单独沉默线虫中的33个基因,他们发现了8个对肠屏障具有显著保护作用的基因,包括IMPA2、SLC25A20、CPT1A、NFU1、CCT8、 TMPP、LIPE和HARS,以及10个对肠屏障具有破坏性作用的基因,包括DTD1、HPX、 SPATA5、DBNL、NUP35、SH3BGR、ATP1B1、L1CAM、SERPINC1和STX1A(图5j-m)。
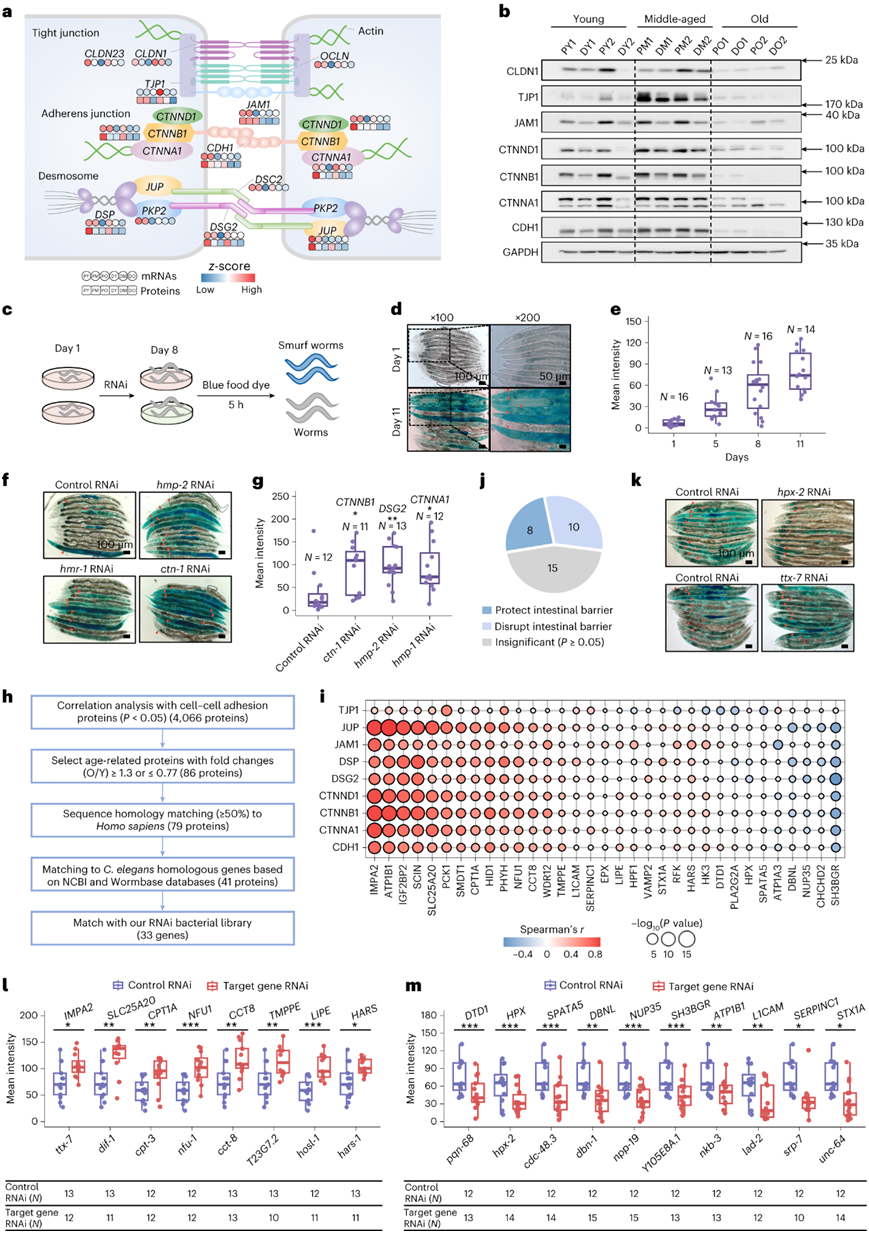
图5. 年龄相关基因对肠道屏障完整性的影响。
(a) 近端和远端结肠中三个年龄组细胞-细胞连接基因的蛋白质和 mRNA 表达量。 (b) 免疫印迹分析。(c) 评估线虫肠道屏障完整性的实验流程。 (d) 线虫的代表性 DIC 显微镜照片,显示染料随着年龄的增长从肠腔渗漏到体腔中。(e) 野生型线虫在 D1、D5、D8 和 D11 上与年龄相关的 Smurf 表型的量化。(f-g) 代表性 DIC 图像和箱线图(显示沉默细胞-细胞粘附相关基因对肠道屏障功能的影响)。 (h) 用于Smurf检测的年龄相关基因筛选流程图。 (i) 气泡图显示 33 种选定的蛋白质和 9 种细胞间粘附蛋白之间的相关性。(j) 饼图显示了表型检测的总体结果。 (k) 在成年第 8 天在蓝色染料中浸泡 5 小时的hpx-2 RNAi 处理、ttx-7 RNAi 处理和对照 RNAi处理的代表性 DIC 图像。 (l-m) 箱线图说明用RNAi敲低相应基因后的平均染料渗漏强度。
06
年龄相关磷酸位点对肠道屏障完整性的影响
通过通路分析获得的见解强调了磷酸化在细胞间粘附调节中的潜在作用,下一步是检测可能影响肠道屏障完整性的特定磷酸位点。基于类似的假设来识别细胞-细胞粘附相关蛋白(图5h),他们获得了944个磷酸位点,这些磷酸位点与九种细胞-细胞连接蛋白中的至少一种表现出显著相关性(图6a)。在年轻组和老年组之间识别出 127 个位点(图6a)。在这些位点中,45个磷酸位点表现出与两个以上细胞粘附相关蛋白显著相关(图6b)。值得注意的是,所有 45 个磷酸位点在人类和猴子中都完全保守。此外,43 种磷蛋白中有 29 种在这两个物种之间具有超过 95% 的同一性(图6c)。在45个磷酸位点中,CTNND1在S349处的磷酸化(CTNND1-S349p)在两个结肠切片中均表现出与年龄相关的减少,并在远端结肠中更为明显(图6d-g)。引入S349E突变来模拟磷酸化的效果,显著增加了细胞-细胞连接蛋白的表达,例如OCLN、CLDN1和JAM1(图6h-i),强调了与年龄相关的CTNND1-S349p 失调导致屏障完整性破坏。
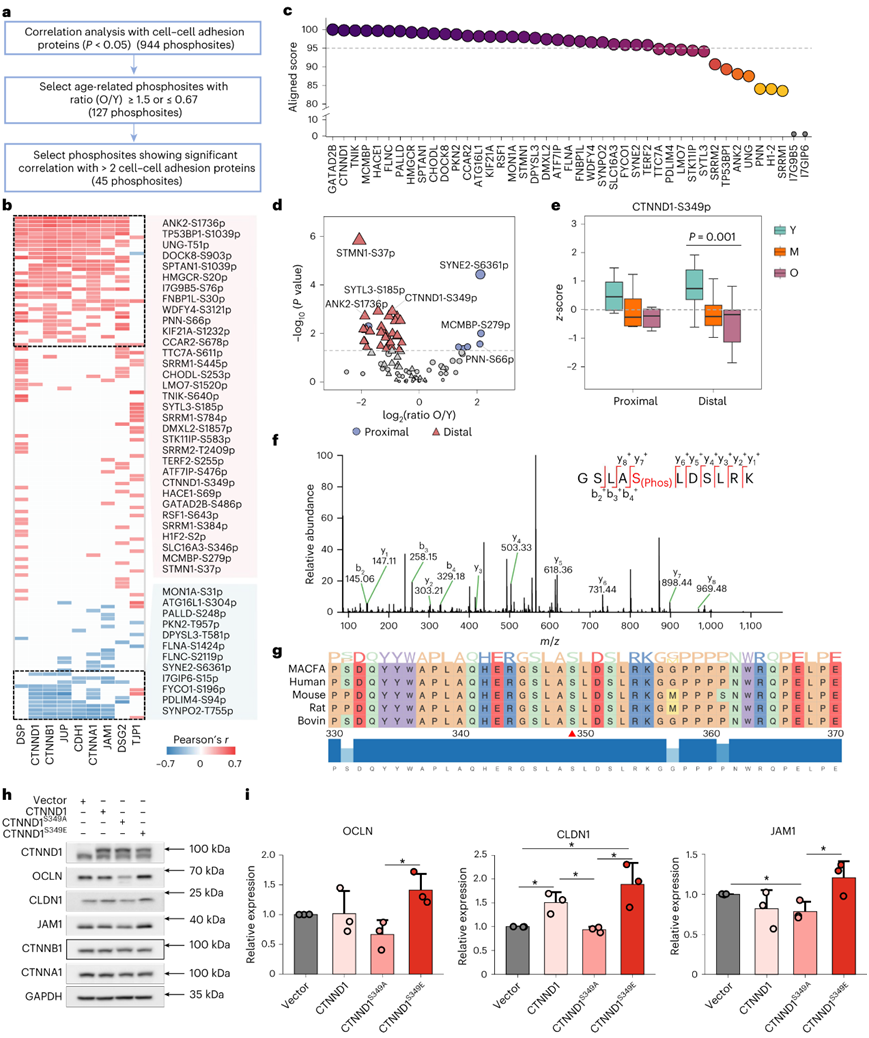
图6. 与年龄相关的磷酸位点对肠道屏障完整性的影响。
(a) 筛选可能影响细胞间粘附蛋白水平的年龄相关磷酸位点的流程图。(b) 热图显示 127 个年龄相关磷酸位点和 9 个细胞-细胞连接蛋白之间的相关性。(c) 通过CLUSTALW对猴子和人类之间对应于45个磷酸位点的43个蛋白质进行序列比对。 (d) 火山图显示年轻和年老猴子近端和远端结肠中 45 个磷酸位点的表达差异。(e) 箱线图显示近端和远端结肠中三个年龄组的CTNND1-S349p的水平。 (f) 在束状分枝杆菌样品中鉴定出的含有 CTNND1-S349p 的磷酸肽的 MS/MS 谱。(g) 不同物种间含有S349残基的CTNND1的部分氨基酸序列的比对。 (h) 免疫印迹分析。(i) 条形图显示通过密度测定法定量的 OCLN、CLDN1 和 JAM1 水平。
07
色氨酸代谢产物对肠道稳态的影响
鉴于近端和远端结肠中色氨酸代谢途径的不同偏好,他们评估了色氨酸及其代谢物的影响,包括5-HT、5-羟基吲哚乙酸(5-HIAA)、KYN、喹啉酸(QA)、犬尿酸(KYNA)和微生物群衍生的3-吲哚丙酸(IPA)对衰老过程中秀丽隐杆线虫肠道形态、肠核和屏障完整性的影响(图7a)。用这些化合物处理显示出对肠萎缩的保护作用(图7b-c),肠核荧光强度显著改善,但5-HT除外,它表现出相反的效果(图7d-e)。此外,在有渗漏的肠道模型中,只有5-HT处理增加了肠道渗漏(图7f-g),并伴有较小的线虫大小(图7f),这可能是由于5-HT的促炎作用。
鉴于到大肠老化的异质性,他们假设5-HT对左侧和右侧结肠炎的影响是不同的。为了验证这一假设,用雄性小鼠建立了葡聚糖硫酸钠(DSS)诱导的结肠炎模型(图7h)。该研究包括未处理的小鼠和DSS、DSS色氨酸、DSS 5-HT或DSS Tph1抑制剂(LP-533401)处理的小鼠(图7h)。Tph1是5-HT生物合成途径中的关键酶,抑制Tph1可减少5-HT的产生。结果表明,5-HT加重了结肠炎,而抑制5-HT生物合成或色氨酸处理可减轻这种作用(图7i-l)。结肠近端和远端H&E染色显示,5-HT或LP-533401治疗对右侧结肠炎影响不大。相比之下,左侧结肠炎的严重程度随着5-HT处理而增加,但通过使用LP-533401抑制5-HT的产生而减轻(图7m)。这一发现强调了大肠的异质性,左侧结肠炎对5-HT波动更敏感。
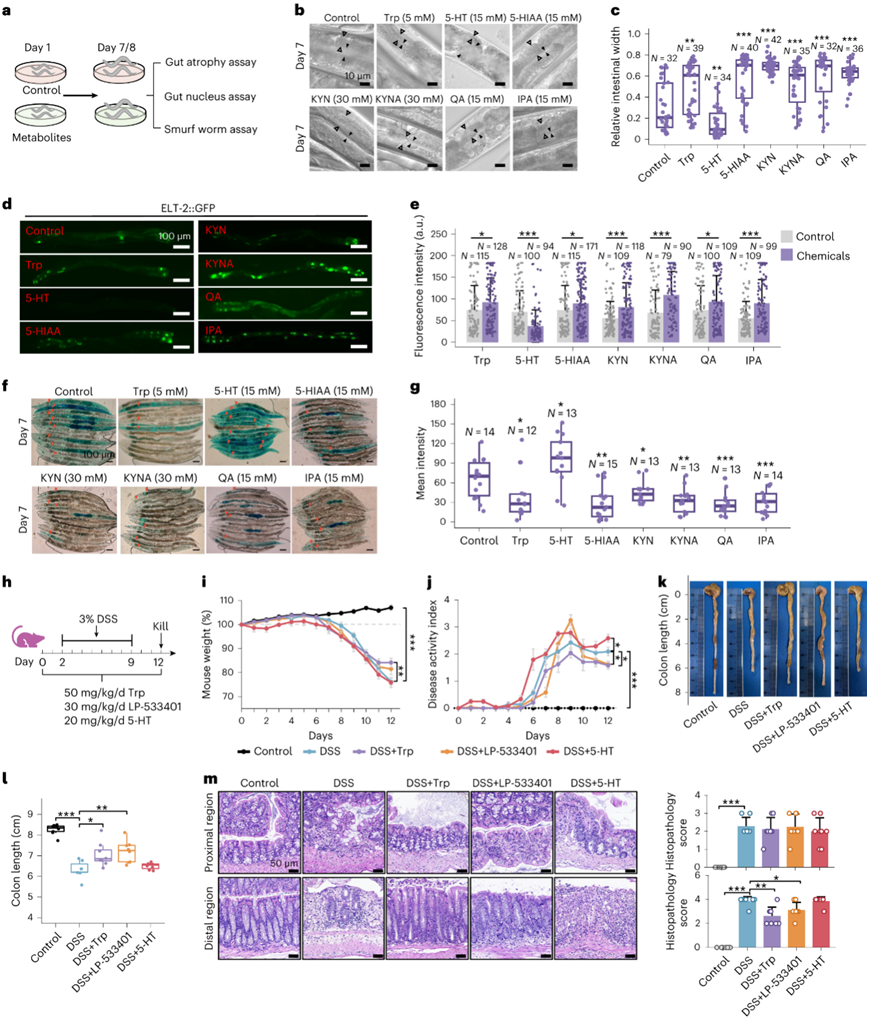
图7. 色氨酸代谢物对肠道稳态的影响。
(a) 探索色氨酸代谢功能的工作流程图。 (b) 代表性 DIC 显微图像显示 D7 上的肠萎缩。(c) 说明D7上相对肠道宽度的统计分析的箱线图。(d) D7 上elt-2 ::gfp 线虫的代表性显微照片。(e) 显示平均 GFP 强度的条形图。(f) D8 上蓝色食用染料中浸泡 5 小时的线虫的代表性 DIC 图像。(g) 代表平均染料渗漏强度的箱线图。(h)实验流程。 (i) 每日小鼠体重。(j) 由体重减轻、粪便稠度和出血确定的疾病活动指数。(k) 小鼠结肠长度的代表性图片。(l) 结肠长度的统计分析。(m) 升结肠和降结肠的 H&E 图像。
+ + + + + + + + + + +
结 论
本项研究探讨了几种调节因子(包括色氨酸代谢调节因子)在秀丽隐杆线虫肠道功能和寿命中的作用。通过犬尿氨酸和血清素 (5-HT) 途径的色氨酸代谢在近端和远端结肠之间有所不同,这表明区域特异性的保守性,并且使用小鼠结肠炎模型,观察到远端结肠炎对 5-HT 治疗更敏感。此外,通过对人类 CRC 样本进行蛋白质组学分析,确定了肠道衰老与 CRC 之间的联系,高 HPX 水平预示着老年 CRC 患者预后不良。总之,这项工作为预防肠道衰老和相关疾病提供了潜在的目标。
+ + + + +



