

 English
English文献解读|Nat Metab(18.9):运动时间对小鼠骨骼生长的影响不同
✦ +
+
论文ID
原名:Time of exercise differentially impacts bone growth in mice
译名:运动时间对小鼠骨骼生长的影响不同
期刊:Nature Metabolism
影响因子:18.9
发表时间:2024.05.28
DOI号:10.1038/s42255-024-01057-0
背 景
影响骨生长的因素有多种,包括钙、维生素D等营养元素,以及运动或体力活动。特别是,体育锻炼或训练有可能提高骨量和强度,最终改善骨形成。一项随机交叉试验表明,在改善2型糖尿病患者的血糖水平方面,下午运动比早上运动更有效。有规律的晨练对超重或肥胖的人有益,因为它可以提高运动的依从性,并有助于体重管理。然而,运动时间对骨生长的潜在影响和机制尚不明确。
实验设计
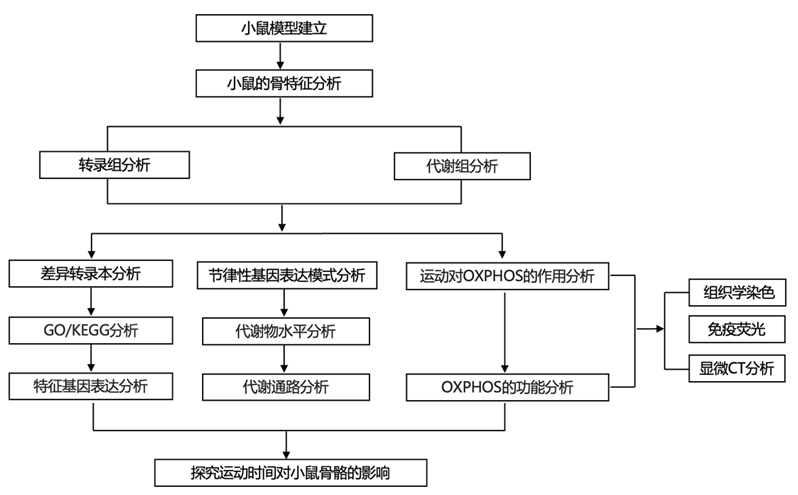
结 果
01

ZT13时的运动可以促进骨骼生长
为了确定运动对骨骼生长是否有特定时间的影响,研究团队建立了小鼠模型,并将雄性小鼠随机分配为7组:在静息期[授时因子时间(zeitgeber time)(ZT) 1、ZT5或ZT9]和活动期(ZT13、ZT17和ZT21)进行30分钟的运动(Exe),以及一个不运动对照组(Sed)(图1a)。让小鼠运动5周后(每周运动5天)或假运动后,雄性小鼠安乐死。微计算机断层扫描(micro-CT)结果显示,与不运动的对照组小鼠相比,ZT1、ZT13、ZT17和ZT21运动组小鼠的四肢骨骼发育显著增强。然而,在ZT5和ZT9运动的小鼠中没有观察到明显的改善(图1b)。特别是,在早期活动期(ZT13)进行运动的雄性小鼠,股骨长度增加了5%,每组织体积骨体积(BV/TV)和小梁厚度(Tb.Th)分别增加了26%和54%(图1b-c)。值得注意的是,无论是在休息阶段还是在活动阶段,运动对老鼠的食物消耗或体重都没有影响。通过检测股骨生长板的形态,发现在ZT1、ZT13、ZT17和ZT21进行运动的雄性小鼠中,增殖层区(PZ)和肥大层区(HZ)的高度显著增加,软骨细胞的数量显著增加(图1d-h)。在早期运动阶段(ZT13)的雄性小鼠中,软骨化中心的EdU+细胞显著增加(图1i-j)。此外,钙黄蛋白染色显示股骨生长板成骨中心的矿物质附着率(MAR)和骨形成率显著增强(图1k-l)。与ZT1阶段相比,在ZT13时运动的雄性小鼠软骨化中心的MAR增加了3倍(图1k-l)。
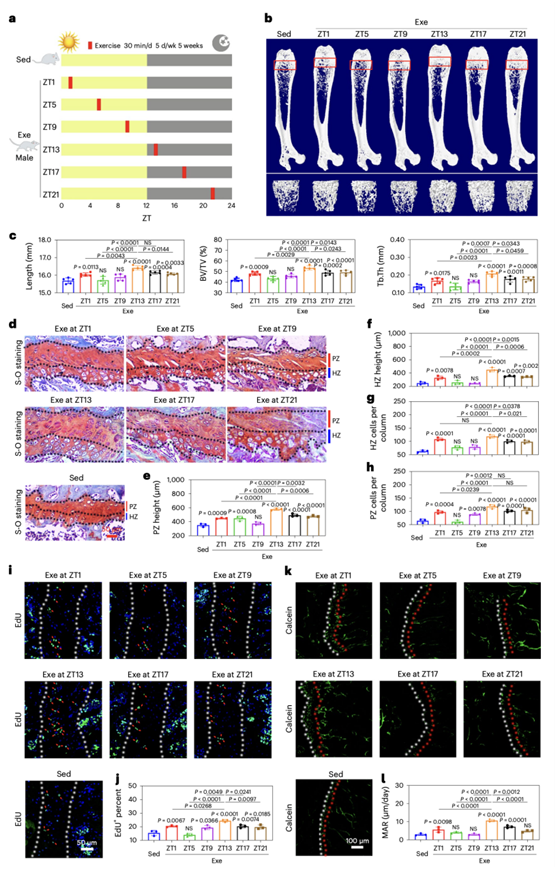
图1. 在早期活动阶段进行锻炼可以有力地促进骨骼生长。
(a) 本研究采用的方案。(b) 久坐和运动的微CT图像的股骨三维可视化。(c) 显微CT结果。(d) 久坐和运动小鼠股骨远端S-O染色的代表性图像。(e-h) 生长板软骨PZ和HZ高度及软骨细胞数。(i-j) 生长板软骨PZ区增殖细胞分析。(k) 久坐和运动小鼠股骨干骺端松质骨钙黄蛋白双标记。(l) 股骨干骺端松质骨矿化附着率(MAR)的定量。
02
在ZT13时,运动促进了骨骼生长的基因表达
白天运动的时间对小鼠的骨骼生长有重大影响,在早期活动期观察到的改善最显著。为了研究运动后骨生长过程的时间特异性反应,他们对软骨成骨组织中转录变异进行了比较分析,这些转录变异来自于在ZT1、ZT13、ZT17和ZT21时进行运动的小鼠。小鼠在固定的跑步机上进行30分钟的运动或假运动5小时后,他们对这些小鼠实施安乐死,然后收集组织样本进行转录组分析(RNA-seq)(图2a)。他们观察到在ZT13的早期活动期运动后,众多转录本和基因的表达发生了显著变化,分别有10.1%和10.3%的总转录本增加和减少。相反,ZT1的早期静息阶段运动导致显著受影响基因的比例较低,分别只有6.4%和3.9%的总转录本增加和减少。在活动期中期(ZT17)和晚期(ZT21)运动后,改变基因的比例介于ZT13和ZT1之间发生的变化(图2b)。他们还发现在ZT1、ZT13、ZT17和ZT21有27个在运动后上调的基因(图2c)。
随后进行GO和KEGG分析,发现上调的基因在运动后早期活动期(ZT13)的骨生长,基质生物合成和能量代谢方面显著增加(图2d-f)。值得注意的是,成骨相关通路如AMPK,在早期活动期(ZT13)的Exe小鼠中发生特异性激活,而HIF-1信号通路在静息期(ZT1)的Exe小鼠中显著富集(图2g)。此外,Exe小鼠在早期活动期(ZT13)表现出与氧化磷酸化(OXPHOS)和三羧酸(TCA)循环相关的基因更明显的富集,而在晚期活动期(ZT21)和静息期(ZT1)表现出与糖酵解相关的基因更明显的富集(图2f)。此外,27个共同上调的基因富集于骨化和基质生物合成相关通路,这些基因在早期活动阶段(ZT13)的表达显著增加(图2h-i)。值得注意的是,热图分析揭示了与基质生物合成和组织相关的基因在运动初期(ZT13)的表达显著增加(图2j)。细胞增殖生物标志物细胞周期蛋白D2 (CCND2)和性别决定区Y(SPY)-盒蛋白9 (SOX9),以及基质合成生物标志物II型胶原(COL2α1)和蛋白聚糖(ACAN)的表达显著升高(图2k-n)。总之,这些结果表明,在早期活动阶段锻炼可以增强与骨生长相关的基因的表达。
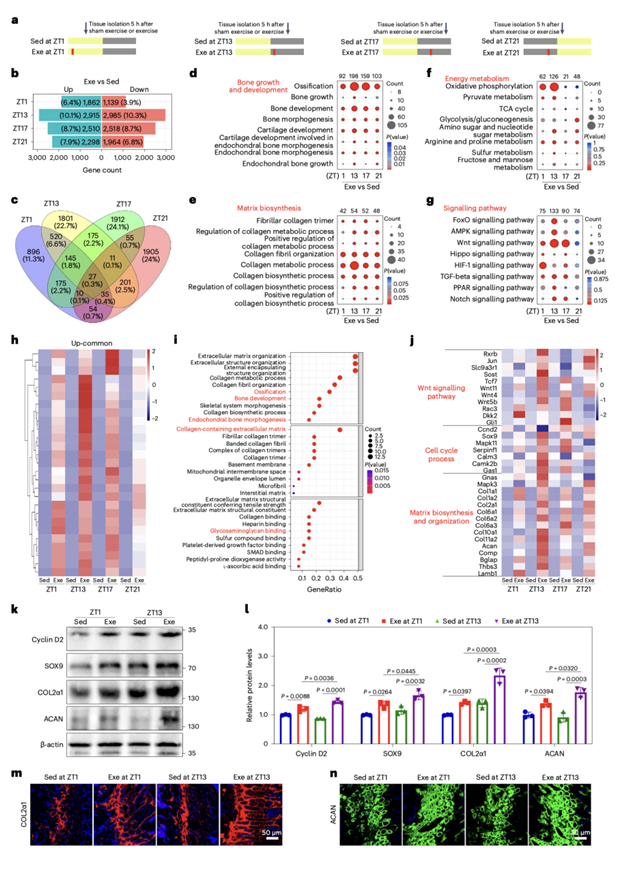
图2. 骨生长相关基因的转录表达在早期活跃期通过运动得到促进。
(a) 假运动治疗后5 h,或在静息期(ZT1)和活动期(ZT13、ZT17和ZT21)运动时分离软骨成骨组织。(b) 运动后软骨成骨组织在早期静息期(ZT1)相对于活动期(ZT13、ZT17和ZT21)的差异表达转录本。 (c) 休息阶段和活动期所共有的与运动相关的上调转录本。(d-e)GO分析。(f-g) KEGG通路分析。(h) 热图显示骨转录本对运动的反应在这两个阶段普遍上调。(i) KEGG分析两个阶段上调转录本。(j) 与WNT信号通路、细胞周期和胶原合成相关的代表性上调基因热图。(k-l)免疫印迹分析和定量。(m-n)免疫荧光分析。
03
ZT13的运动与骨代谢节律相协调
他们在5周运动的最后1小时剧烈运动或假运动后0、6、12、18、24、30、36、42和48小时对小鼠实施安乐死,然后收集组织样本进行分析(图3a),大约1600个基因(占总基因的6.7%)在从Sed小鼠分离的软骨成骨组织中表现出节律性表达。然而,6415个基因在运动早期(ZT13)发生昼夜节律变化,这一反应显著超过了在其他时间点(ZT1、ZT5、ZT17和ZT21)观察到的效应(图3b)。与静息期(ZT1和ZT5)相比,Sed和Exe在活动期(ZT13、ZT17和ZT21)的转录本有更大的重叠。具体而言,ZT13显示Sed和Exe之间有476个重叠转录本,而ZT1和ZT5中Sed和Exe呈现昼夜节律变化的基因数量分别只有183和33个。此外,ZT13的运动保持了与Sed相当的周期分布,而ZT1、ZT5、ZT17或ZT21的运动诱导了节律性基因的周期变化(图3c)。他们进一步根据基因的表达模式在Sed组中鉴定出4个聚类(图3d)。聚类1的基因在ZT6表达最高,在ZT18表达最低。聚类2基因在ZT0后表达增加,在昼夜转换(ZT12)前后达到峰值。相比之下,聚类3基因在光照开始后表达下降,在昼夜转换期间观察到的水平最低,然后在静息期表达减少,在活动期表达增加。聚类4的基因在光照开始后下调,在ZT6达到最低点,在ZT18左右达到峰值,然后在24小时后下降到基础水平(图3d)。在活动期(ZT13、ZT17和ZT21)运动可以保持与Sed对照组相似的表达模式(图3d)。相反,在静息期(ZT1和ZT5),运动小鼠的昼夜表达模式发生显著改变,例如,在聚类1中,它们并没有在ZT18达到最低点,而是从ZT12持续下降到ZT24(图3d)。为了深入了解运动的日常生物学特征,他们进行了GO分析,在ZT13 时,Exe组显示出与骨发育和代谢过程相关的转录本的富集(图3e)。虽然Nr1d1和Rorc在ZT1 Exe组中表现出节律性变化,但核心生物钟基因Bmal1基本没有节律性表达(图3f)。这些观察结果证实了这样一种观点:即在早期活动阶段的锻炼可以增加节律性基因表达和维持昼夜节律稳态,最终协同促进骨骼生长。
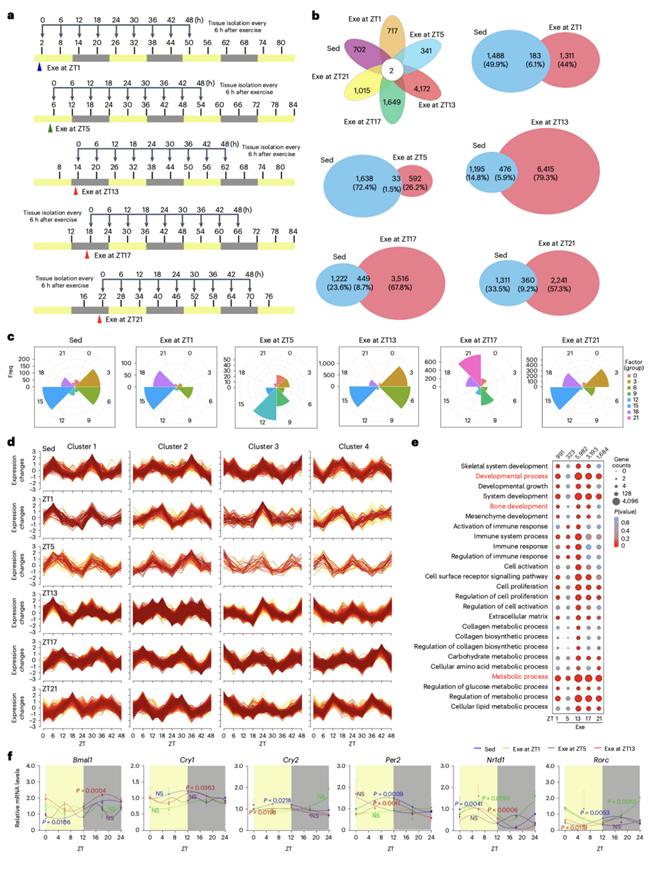
图3. ZT13运动与内在骨代谢节律协同作用激活骨生长相关基因的表达。
(a) 实验设计。(b) 维恩图显示了不运动(Sed)和运动(Exe)小鼠在早期静息期(ZT1)、中期静息期(ZT5)、早期活动期(ZT13)、中期活动期(ZT17)和晚期活动期(ZT21)软骨成骨组织中的基因表达。 (c)不运动(Sed)和运动(Exe)小鼠的基因变化。(d)分析Sed和Exe小鼠节律性基因的表达模式。根据表达模式的相似性检测出4个聚类。(e) GO分析。(f) 定量PCR与逆转录分析核心节律基因的表达。
04
在ZT13时运动显著激活OXPHOS
为了验证运动与成骨组织中OXPHOS的相关性,他们利用微型正电子发射断层扫描(PET)计算机断层扫描仪监测小鼠运动后 2-[18F]-2-脱氧葡萄糖(18F-FDG)的摄取。无论运动的时间如何,运动均导致股骨软骨成骨组织的葡萄糖摄入量显著升高(图4a-b)。然而,他们发现在ZT13的运动导致ATP生成增加和氧化烟酰胺腺嘌呤二核苷酸(NAD+)/还原型烟酰胺腺嘌呤二核苷酸(NADH)的比率升高(图4c-d)。为了研究一天中不同时间运动后代谢通路的变化,他们进行了代谢组学分析。研究发现,与TCA和OXPHOS相关的代谢物,特别是α -酮戊二酸和L-苹果酸的水平,在ZT13运动后增加(图4e)。相反,ZT1时的运动导致糖酵解代谢物(如β -D-果糖6-磷酸和乳酸)水平显著升高(图4e)。
随后,他们对转录组和代谢组数据进行了深入分析,以确定代谢酶和代谢物之间的相关性。在ZT13运动后,九个编码三羧酸循环代谢酶的差异表达基因(DEG)表现出显著上调。这些基因包括4个编码异柠檬酸脱氢酶(ICDH)的基因,2个编码琥珀酰辅酶A合成酶(SCS)的基因,1个编码琥珀酸脱氢酶(SuDH)的基因和2个编码苹果酸脱氢酶(MDH)的基因(图4H)。此外,编码电子传递链的线粒体复合物I、III、IV和V的DEG在ZT13的运动后表达显著增加。相反,编码糖酵解必需调节酶的DEG的表达在ZT1运动后显著增加。这些基因包括1个编码磷酸果糖激酶(PFKP)的DEG、2个编码磷酸甘油酸激酶(PGK)的DEG和1个编码乳酸脱氢酶(LDH)的DEG(图4g)。早期活动阶段的运动增加了OXPHOS基因节律性表达的水平。相应的,编码OXPHOS关键酶的蛋白在ZT13运动后显著上调,而编码糖酵解酶的蛋白在ZT1运动后显著上调(图4h-i)。综上所述,这些结果表明,在早期运动阶段进行运动可引发更强劲的TCA循环、OXPHOS和更多的ATP生成。
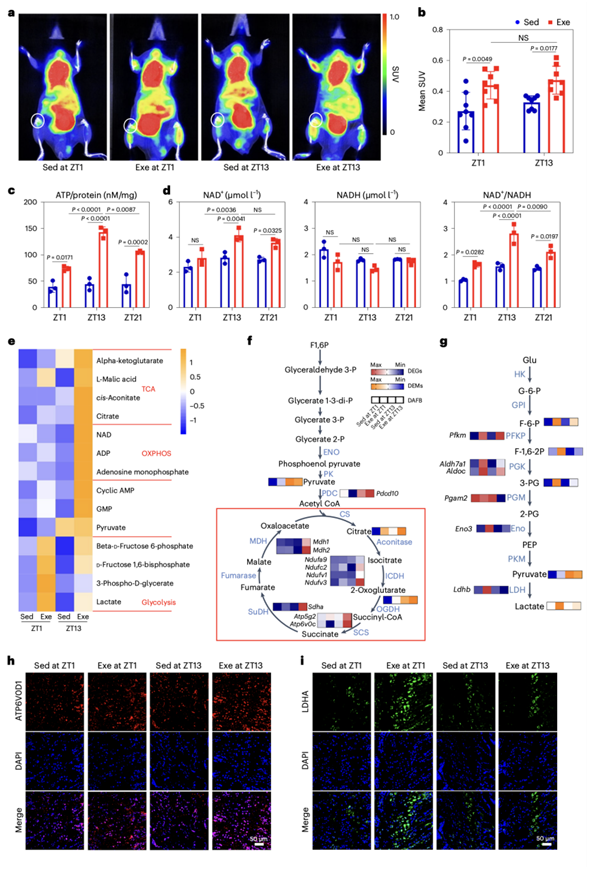
图4. 在早期活跃期,运动显著激活OXPHOS。
(a) 运动后股骨软骨成骨组织在休息早期和活动早期18F-FDG摄取变化的PET分析。(b) 平均标准化摄取值的PET分析。(c) 使用多功能酶标记仪检测股骨在休息早期、活动早期和活动晚期运动后的ATP生成水平。(d) 检测运动后股骨静息早期、运动早期和运动晚期的NAD+、NADH含量及NAD+/NADH比值。(e) 热图显示运动后早期休息期相对于活动期的TCA循环、糖酵解和OXPHOS代谢产物的变化。(f-g) 参与OXPHOS、TCA循环和糖酵解的DEG和差异表达代谢物(DEM)的表达谱。(h-i)免疫荧光分析。
05
ZT13的运动通过OXPHOS刺激骨骼生长
为了进一步研究OXPHOS在运动后早期促进骨生长中的作用,他们在运动前对小鼠进行了髓内或腹腔注射OXPHOS抑制剂寡霉素A (oligomycin A)(图5a)。相比于晚期活动阶段或早期休息阶段,在小鼠的早期活动阶段注射寡霉素A,运动对骨骼长度和质量的增强能力减少得更为显著(图5b-c)。在早期活动期,寡霉素A处理的小鼠生长板软骨高度和软骨细胞数量显著降低(图5d-e)。此外,EdU染色显示,在活动期早期,寡霉素A处理的小鼠PZ软骨细胞的活化增殖能力发生显著逆转(图5f-g)。这些结果表明,在高强度的身体活动中,特别是在早期活动阶段,OXPHOS对ATP的生成和骨形成是必不可少的。
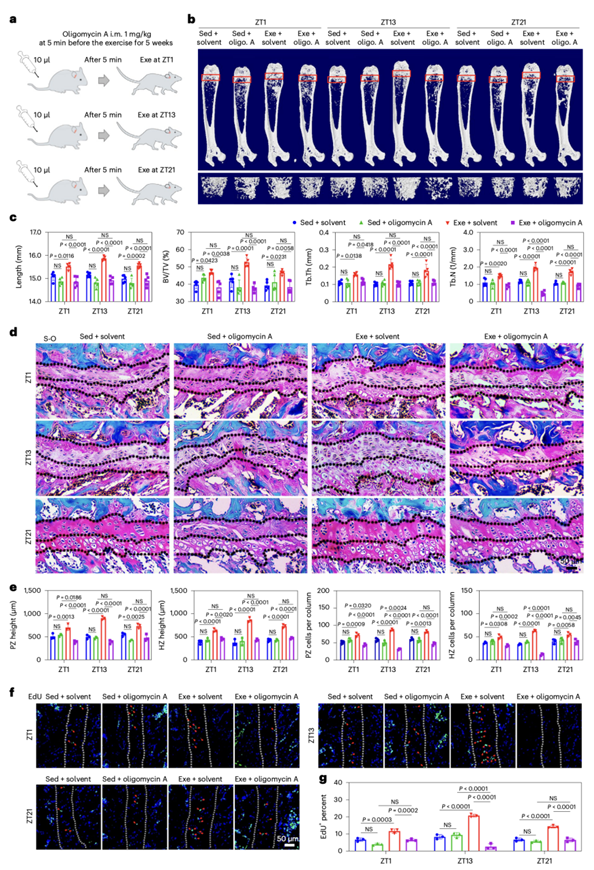
图5. 活动期的运动通过OXPHOS过程刺激骨骼生长。
(a) 实验设计。(b) 代表性显微CT图像。(c)显微CT结果。(d-e)通过S-O染色的股骨远端代表性图像和分析。(f-g) 注射OXPHOS抑制剂或等量溶剂的Sed和Exe小鼠,EdU染色生长板PZ软骨内增殖细胞的分析。
06
在 ZT13时激活OXPHOS可促进骨生长
接下来,他们对野生型小鼠(每日200 mg / kg体重,持续28天)注射5-氨基咪唑-4-甲酰胺核糖核苷(AICAR)(一种OXPHOS激活剂)。在早期活动期腹腔内或髓内注射AICAR后,micro-CT结果表明,AICAR处理的小鼠表现出显著改善的骨生长(图6a-b)。在早期活跃期,AICAR处理的小鼠PZ软骨细胞数量明显增加(图6c-d)。EdU染色和免疫荧光结果显示软骨细胞增殖和基质生物合成增强(图6e-h)。这些结果表明,在早期活跃期对OXPHOS进行药理激活可能是刺激骨骼生长的一种方式。
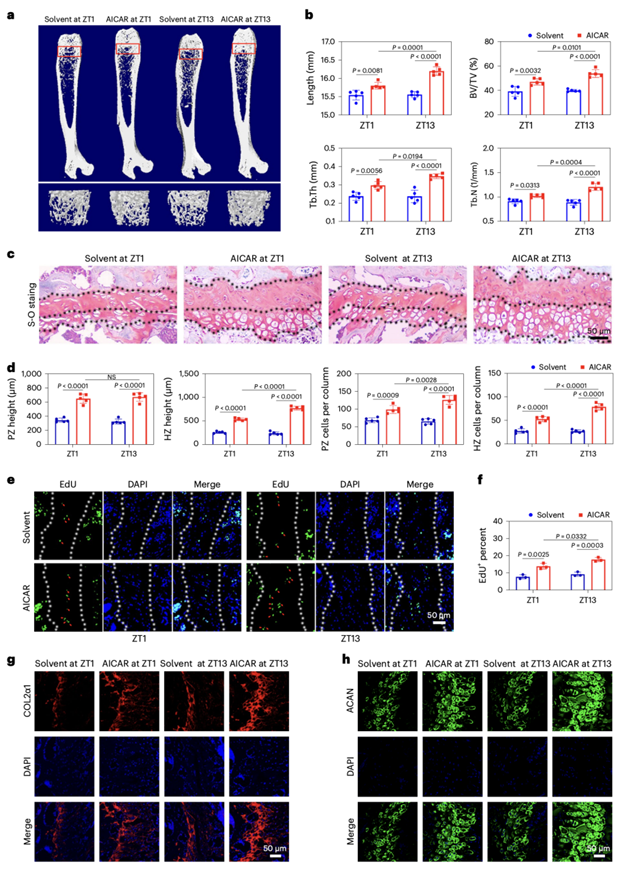
图6. 在早期活跃期通过药物刺激OXPHOS可促进骨生长。
(a) 小鼠股骨代表性显微CT图像。(b) 显微CT结果。(c-d) 在经腹腔注射OXPHOS激活剂或等溶剂的Sed和Exe小鼠中,S-O染色的股骨远端代表性图像和分析。(e-f) 分析腹腔注射OXPHOS激活剂的Sed和Exe小鼠生长板PZ软骨内EdU染色的增殖细胞。(g-h) 免疫荧光分析。
+ + + + + + + + + + +
结 论
通过转录组学和代谢组学分析,本项研究发现在早期活动阶段进行运动可显著上调与骨骼发育和代谢相关的基因。值得注意的是,氧化磷酸化相关基因在软骨化中心表现出节律性表达,在早期活动阶段达到峰值,此时骨代谢中更多节律性基因表达,并且通过影响氧化磷酸化协同促进骨骼生长。
+ + + + +



