

 English
English文献解读|Bone Res(12.7):人类肌腱粘连的多组学分析表明 ACKR1 调控的巨噬细胞迁移参与再生
✦ +
+
论文ID
原名:Multi-omics analysis of human tendon adhesion reveals that ACKR1-regulated macrophage migration is involved in regeneration
译名:人类肌腱粘连的多组学分析表明 ACKR1 调控的巨噬细胞迁移参与再生
期刊:Bone Research
影响因子:12.7
发表时间:2024.05.07
DOI号:10.1038/s41413-024-00324-w
背 景
肌腱粘连是肌腱损伤后的常见并发症,若没有有效的抗纤维化疗法,则会形成积聚的纤维化组织,从而导致严重残疾。巨噬细胞是肌腱粘连形成过程中的纤维化触发因素。然而,不同种类的巨噬细胞具有不同的功能并接受多重调控,这些都仍是未知的。
实验设计
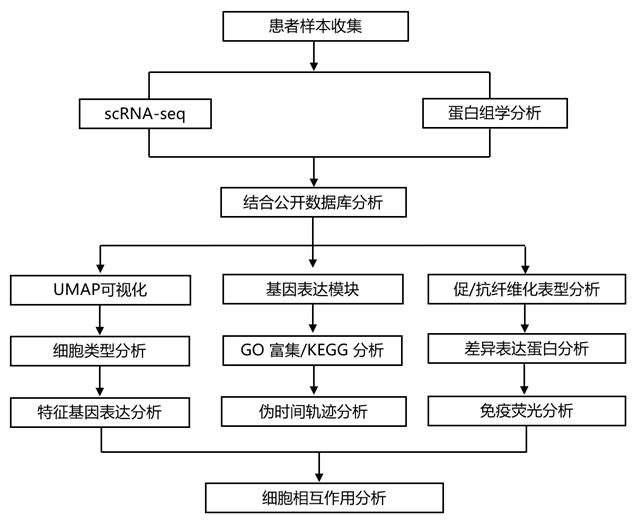
结 果
01

人类肌腱粘连组织的单细胞转录组图谱
为了确定肌腱粘连组织的细胞组成,研究者团队从 12 名患有屈肌腱肌腱溶解症的患者中手术获取组织样本,包括 3 个正常肌腱周围组织样本和 3 个来自损伤后 3、10 天(dpi:损伤后天数) 和 12 周(wpi:损伤后周数)的病变病理样本,立即对样本进行单细胞转录组分析(scRNA-seq)(图1a)。共保留了12名患者的74350个细胞以供后续分析,其中10081个细胞来自正常腱周组织,18866个细胞来自3 dpi组织,14953个细胞来自10 dpi组织,30 450个细胞来自12 wpi组织(图1b)。
首先将细胞核以粗略方式聚类,然后用细胞类型特异性标记物对每个聚类进行注释,建立了正常腱鞘和受伤组织中的细胞类型图谱(图1c)。
他们鉴定了8个主要的细胞群,包括由CD14分子(CD14)和CD68分子(CD68)标记的单核吞噬细胞(MP),由血小板和内皮细胞粘附分子1 (PECAM1)和血管性血友病因子(VWF)标记的内皮细胞(EC),分别由配对同源盒1 (PRRX1)和Thy-1细胞表面抗原(THY1)鉴定的间充质细胞(MC),由克隆刺激因子3受体(CSF3R)和Fc γ受体IIIb (FCGR3B)标记的中性粒细胞,B细胞表达跨膜4结构域A1 (MS4A1)和CD79a分子(CD79a),T细胞表达T细胞受体复合物CD3 δ亚基(CD3D)和T细胞受体复合物CD3 ε亚基(CD3E),肥大细胞标记跨膜4结构域A2 (MS4A2)和KIT原癌基因、受体酪氨酸激酶(KIT)以及钙粘蛋白19 (CDH19)和S100钙结合蛋白B (S100B)鉴定的雪旺细胞。
炎症细胞、内皮细胞和MC的比例根据肌腱粘附阶段的不同而不同(图1d-e)。在正常腹膜组织中,MCs是最丰富的细胞类型,占总数的52.9%(图1d)。在第3 dpi(炎症期),MC和EC的比例较低(分别占总细胞的0.3%和0.5%),而包括单核吞噬细胞、T细胞、B细胞和中性粒细胞在内的炎症细胞的比例增加;此阶段最丰富的细胞类型是MP(55.3%),其次是T细胞(25.5%)和中性粒细胞(14.7%)(图1d)。在10 dpi(增殖期)时,炎症细胞数量下降,巨噬细胞仅占所有细胞的18.8%。同时,EC和MC广泛增殖,分别占细胞总数的22.4%和41.8%。在12 wpi(重塑期)时,成纤维细胞已经迁移到病变处,并出现在肉芽组织中,从而诱导肌腱粘附。MC (71%)和EC(19.6%)是这一阶段的主要细胞类型,炎症细胞较少(图1d)。在12 wpi时,主要细胞类型的分布与正常肌腱周组织相似,表明在肌腱粘连的晚期,组织状态趋于稳定。基于这些细胞群的动态,他们推测MP、EC和MC在肌腱粘附的发展中起重要作用。
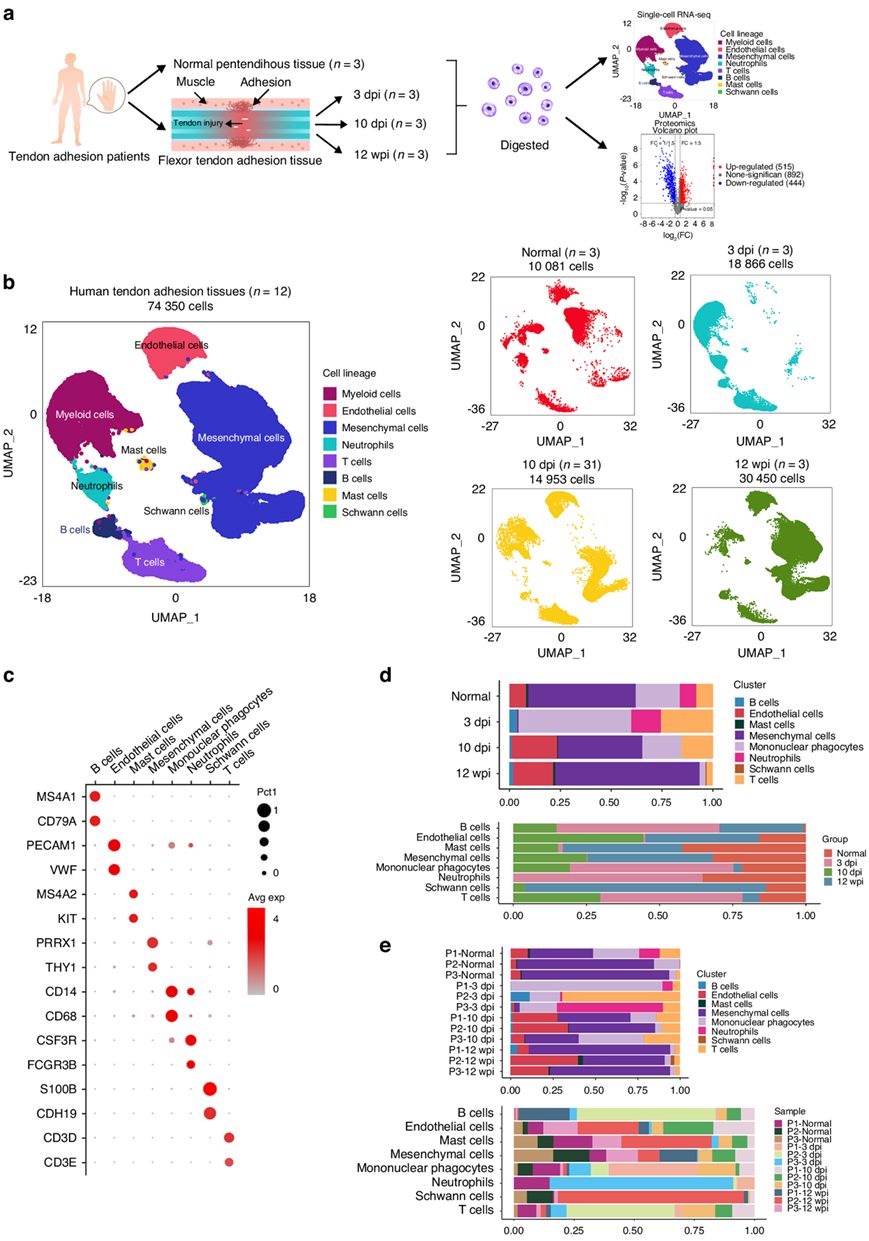
图1. 人类正常肌腱周围组织和肌腱粘连组织的单细胞图谱。
(a) 本研究设计概述。(b)UMAP可视化。(c) 点图:显示已知标记的人类肌腱粘连组织的细胞聚类。(d) 各时间点人体肌腱粘连组织中八种主要细胞类型比例的条形图。(e) 每位患者的人体肌腱粘连组织中八种主要细胞类型比例的条形图。
02
内皮细胞的独特亚群存在于纤维化微环境中
通过PECAM1和VWF的高表达水平鉴定了肌腱粘附组织中的EC(图1c)。然后根据不同的标记进一步将EC亚群分为四个亚群,包括间隙连接蛋白α 4 (GJA4)、细胞周期调节剂(RGCC)、非典型趋化因子受体1 (ACKR1)和C-C基序趋化因子配体21 (CCL21)(图2a-d)。在3 dpi时,人肌腱粘连组织中几乎不存在内皮细胞,有两个样本缺乏EC(图2e)。GJA4和信号素3G (SEMA3G)在人动脉中特异表达,提示ENDO0为动脉EC(图2b-c)。RGCC作为人毛细血管的标记物,提示ENDO1为毛细血管EC(图2b-c)。ENDO2高水平表达ACKR1, ACKR1在人静脉中特异表达,提示ENDO2为静脉EC(图2b-c)。以CCL21和podoplanin (PDPN)标记的ENDO3鉴定为淋巴EC(图2b-c)。最近的研究报道RGCC和ACKR1在ECM组织中起作用。EC主要在10 dpi和12 wpi时增殖(图2f), ENDO1在10 dpi时扩张(图2e)。肌腱损伤后,与正常肌腱周组织相比,共三个阶段,肌腱粘附组织中EC信号通路上调,包括ecm受体相互作用和局灶性粘附(图2g)。
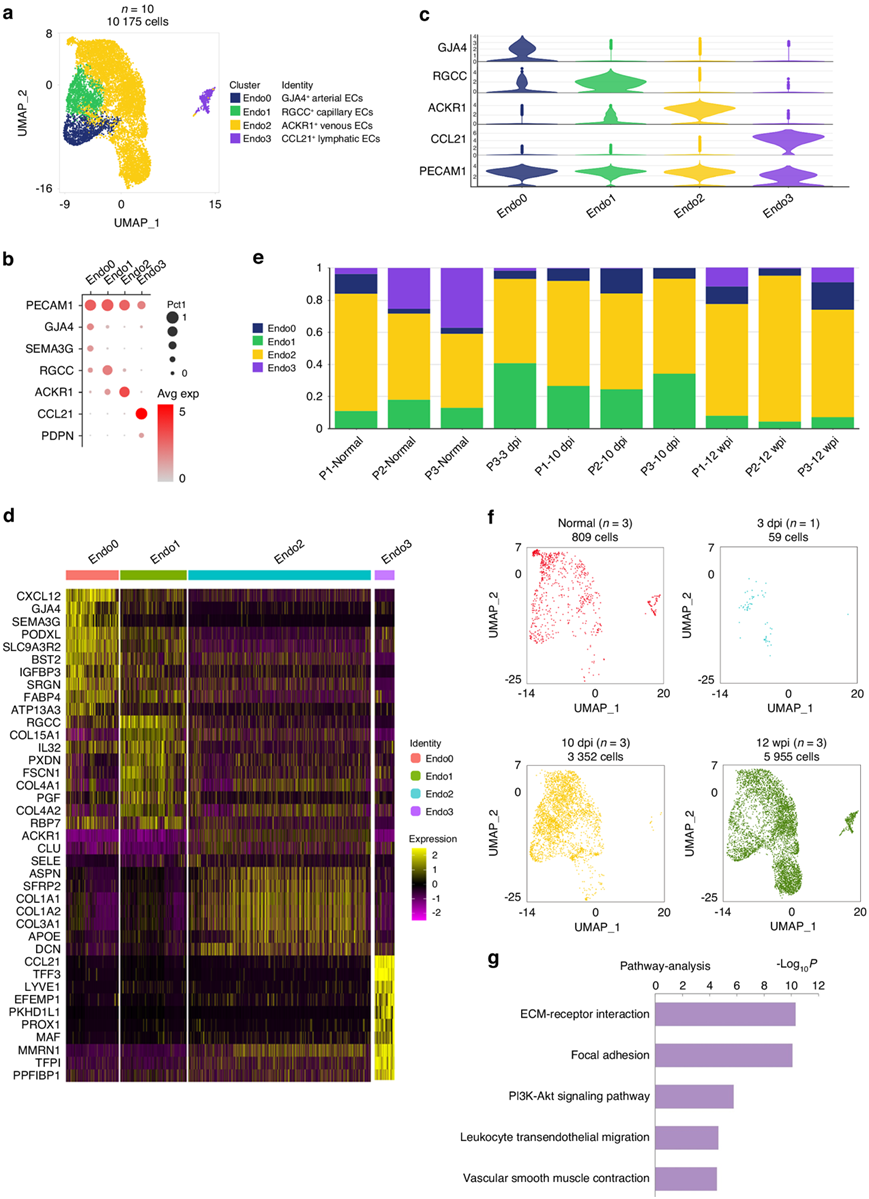
图2. 人类肌腱粘连组织中存在不同的 EC 聚类。
(a)从总共 10 名患者中聚类出 10 175 个内皮细胞。EC,内皮细胞。(b)点图:按已知标记显示内皮细胞的细胞聚类。(c) EC中每个聚类的选定基因表达的小提琴图。(d)每个 EC 聚类中标记基因的热图。(f)每个时间点的 EC 的 UMAP 图:正常、3 dpi、10 dpi 和 12 wpi。(g)肌腱损伤后所有三个阶段上调信号通路的通路分析。
03
伪时间分析
进一步对伪时间轨迹的分析表明,损伤后,ENDO1 分化为 ENDO0 再分化为 ENDO2,并且 ENDO1 的起源是内皮祖细胞,构成了 3 个不同的基因表达模块(图3a)。ENDO1 到 ENDO2 伪时间轨迹的差异基因模块热图表明,包括 SPP1、FN1、COL1A1 和 MMP9 在内的纤维化基因在此过程中上调(图3b)。模块1 的 GO 富集分析显示,在此过程中参与 ECM 组织的基因发生上调(图3c)。GO富集分析揭示了 ENDO1 中的血管生成和 ECM 组织,以及 ENDO2 中的 ECM 组织和胶原原纤维组织(图3d)。标记通路分析表明在ENDO1 和ENDO2 中都存在ECM-受体相互作用。EC 富集通路的 Qusage 分析表明,包括NOTCH信号通路在内的纤维化相关通路在ENDO1中上调(图3e)。KEEG 分析揭示了 ENDO1 和ENDO2中 ECM-受体相互作用的富集通路(图3f)。转录因子(TF)热图显示 ENDO1 富集了促纤维化 TF,包括Jun 原癌基因(JUN),而 ENDO2 富集了促纤维化 TF,包括Fos 原癌基因(FOS)。细胞实验表明 ACKR1 + EC促进纤维化,从人类 10 dpi 组织中分离并培养了 ACKR1 + EC,体外实验表明这些细胞增加了人成纤维细胞中COL1A1的 mRNA 水平(图3g)。ACKR1+ EC的存在毗邻COL1A1,在10 dpi时累及人肌腱粘连组织的纤维化生态位(图3h)。此外,CD31、RGCC、ACKR1和GJA4在10 dpi处的共定位表明,GJA4+ EC和RGCC+ EC可能在肌腱粘连进展过程中转化为ACKR1+ EC(图3h)。总的来说,他们确定了人肌腱粘附组织中的EC亚群,表明ACKR1+ EC有助于人肌腱粘附的纤维化。
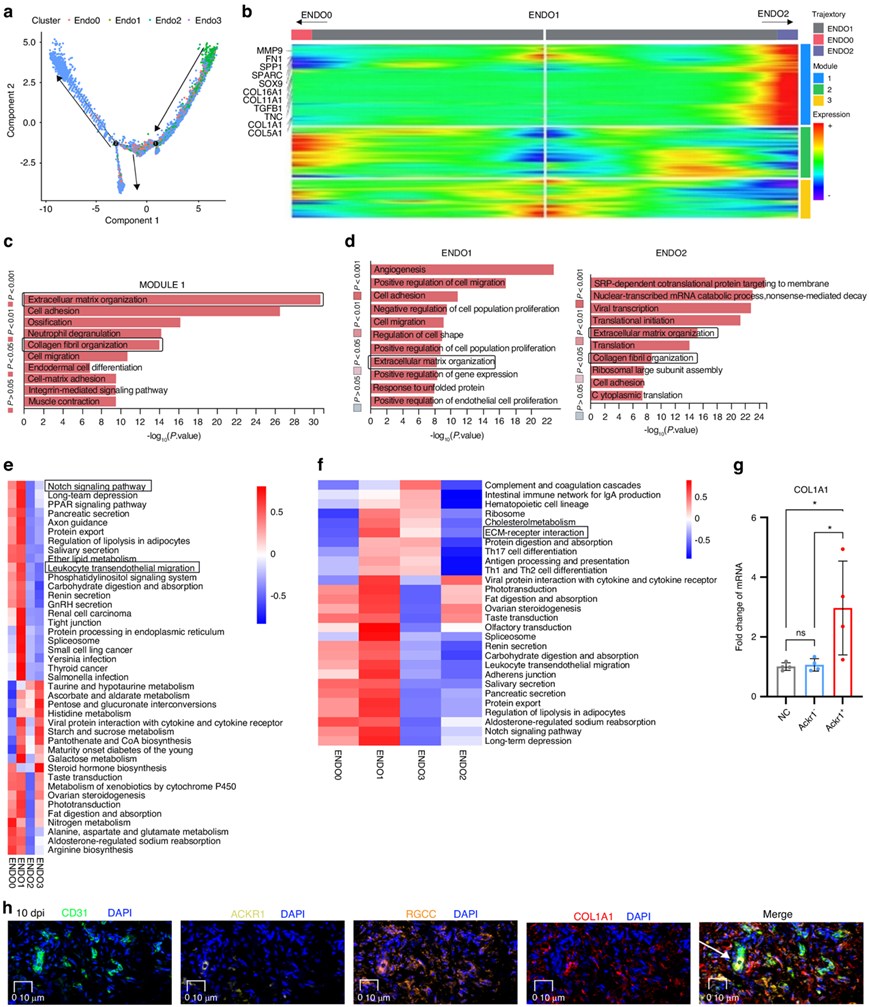
图3. 识别促纤维化的 EC。
(a) ENDO0、ENDO1 和 ENDO2 的伪时间轨迹分析。(b) ENDO1 到 ENDO2(右箭头)和 ENDO1 到 ENDO0(左箭头)伪时间轨迹上的差异基因模块热图。 (c)沿着ENDO0 到 ENDO2 伪时间轨迹,模块 1 中所有基因的 TOP 15 基因本体富集。(d) ENDO1(左)和 ENDO2(右)的GO富集分析。(e)每个EC 聚类的富集通路的 Qusage 分析。(f)对每个 EC 聚类的富集通路进行 KEGG 分析。(g) qPCR分析。(h) CD31(EC 标记物)、ACKR1、RGCC和COL1A1的多色免疫荧光分析。
04
肌腱粘连组织中存在明显的 MP 聚类
他们表征了五个 MP 亚群,包括树突状细胞 (DC)、单核细胞和巨噬细胞(图4a)。MP4定义为dc,由CD1c分子(CD1c)和Fc epsilon受体Ia (FCER1A) 标记(图4b-e)。MP0定义为富含S100钙结合蛋白A8 (S100A8)和S100钙结合蛋白A12 (S100A12)的单核细胞(图4b-e)。将MP1、MP2和MP3定义为巨噬细胞,由CD68、载脂蛋白E (APOE)和补体C1q A链(C1QA) 标记。MP1 富含白细胞介素 1β (IL1B)和表皮调节素 (EREG),GO分析表明 MP1 参与炎症反应(图4b)。因此,MP1定义为促炎巨噬细胞。MP2以促纤维化巨噬细胞为特征,通过分泌磷酸化蛋白1 (SPP1)和基质金属肽酶9 (MMP9)进行标记(图4b-c)。正常腹膜周围组织主要由MP3组成,占总MP的70.9%(图4d)。肌腱损伤后,MP1和MP2在3 dpi时显著升高,分别占48.9%和15.9%,而FOLR2的比例下降至30.2%(图4d)。肌腱损伤导致粘连组织内MP3丰度降低,在粘连进展过程中,这一阶段巨噬细胞计数最高(图4f)。MPs在四个不同阶段的促炎性和抗炎性细胞因子表达水平显示,细胞因子在3 dpi时水平最高(图4g),表明炎症性跟腱损伤在3 dpi时最活跃。随着粘连的进展,10 dpi时MP1和MP2比例降低,而MP3增加(图4d)。巨噬细胞数量逐渐减少,到12周时,肉芽组织形成,巨噬细胞数量减少。巨噬细胞中的 MP3 比例接近 10 dpi,但仍低于正常的腱周组织(图4d)。肌腱损伤后,共三个阶段与正常的肌腱周围组织相比,GO分析显示炎症相关反应上调(图4h),肌腱粘连组织中MP上调的信号通路包括在肌腱损伤中报道的HIF-1信号通路、TNF信号通路和IL-17信号通路(图4i)。与正常的肌腱周围组织相比,肌腱损伤后MP下调的信号通路包括MAPK信号通路(图4j)。
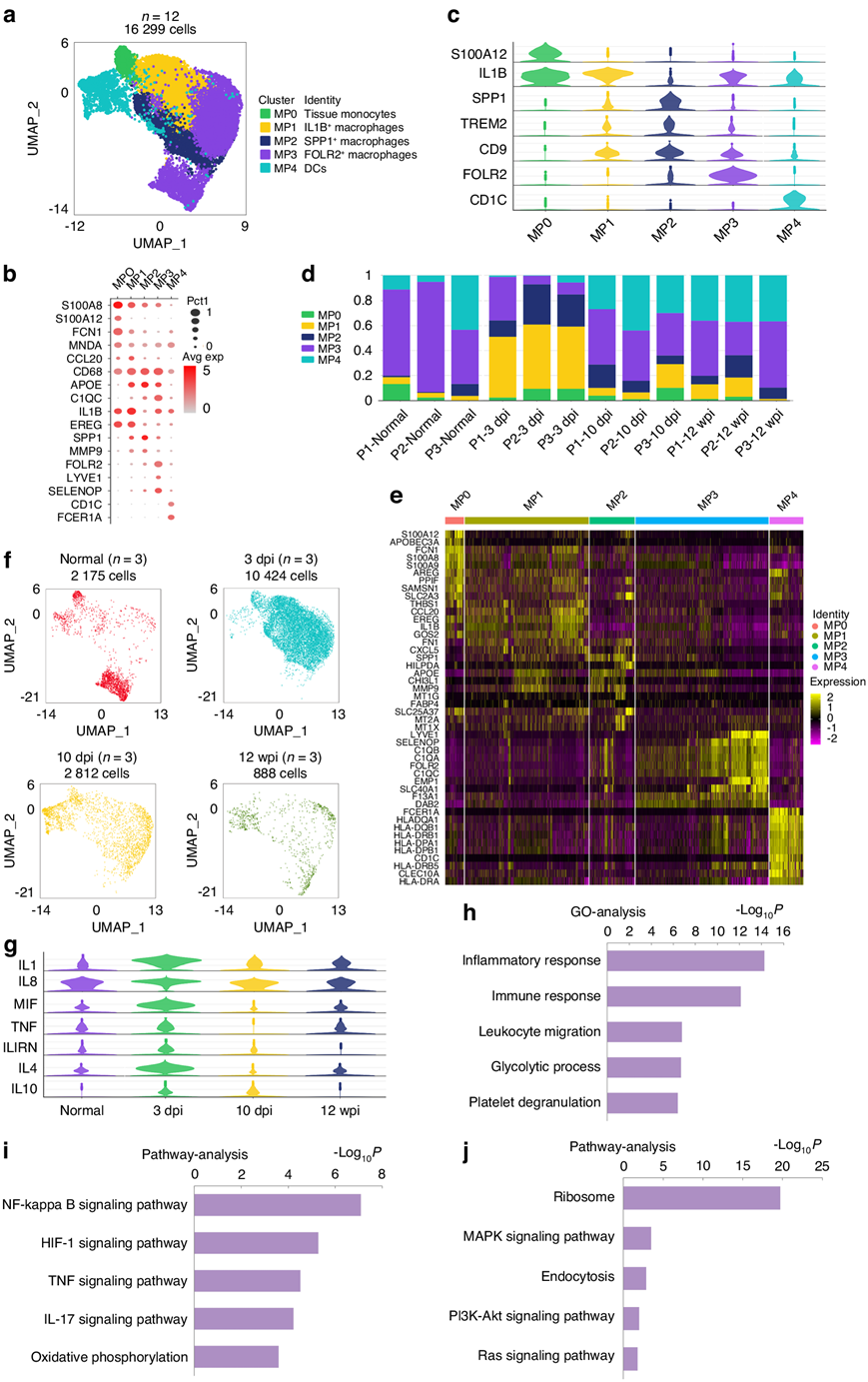
图4. 人类肌腱粘连组织中存在不同的 MP 聚类。
(a)从总共 12 名患者中聚类出 16 299 个单核吞噬细胞 (MP)。(b)点图:按已知标记显示 MP 的细胞聚类。(c) MP中每个聚类的选定基因表达的小提琴图。(d) 12名患者的 MP 中每个聚类的比例条形图。(e)每个MP 聚类中标记基因的热图。(f)每个时间点的 MP 的 UMAP 图:正常、3 dpi、10 dpi 和 12 wpi。(g) MP中促炎和抗炎细胞因子表达水平的小提琴图,包括促炎细胞因子如IL1、IL8、MIF、TNF,以及抗炎细胞因子如IL1RN、IL4、IL10。(h)肌腱损伤后三个阶段上调基因的GO分析。(i) 肌腱损伤后三个阶段上调信号通路的通路分析。(j)肌腱损伤后三个阶段下调信号通路的通路分析。
05
SPP1 +巨噬细胞的促纤维化表型和FOLR2 +巨噬细胞的抗纤维化表型
在最近的研究中,人类纤维化肝组织中的 TREM2 + SPP1 +巨噬细胞鉴定为可促进肝纤维化的瘢痕相关巨噬细胞。本研究中,他们已经在人类肌腱粘连组织中鉴定了标记有SPP1 的MP2、在髓样细胞上表达的触发受体 2 (TREM2)和MM9(图4b-c)。然而,人类肌腱周围巨噬细胞亚群的个体发育仍不清楚。为了进一步研究 MP2 的起源,他们将伪时间轨迹可视化。这些分析表明,在受伤时,分化轨迹从 MP0 发展到 MP1,然后从 MP1 发展到 MP2(图5a),MP0 的起源是血液中的循环单核细胞,这表明 MP2 是单核细胞衍生的,这与之前的研究一致。为了进一步表征MP2表型,他们分析了MP2表达基因的热图,并观察到促纤维化基因SPP1、TREM2和CD9分子(CD9)的上调(图4c)。GO分析结果表明MP2参与了细胞外基质(ECM)组织(图5b),KEEG分析表明细胞参与了ECM受体相互作用(图5d)。标记通路分析提示MP2与ECM受体相互作用。对每个MP亚群中富集的通路的Qusage分析显示,包括ecm受体相互作用和糖酵解在内的纤维化相关通路在MP2中上调(图5c)。糖酵解与炎性巨噬细胞有关。MP2 ecm受体相互作用标志基因组的基因富集提示SPP1和纤维连接蛋白1 (FN1)等纤维化基因上调。MP2糖酵解标志基因组基因富集提示甘油醛-3-磷酸脱氢酶(GAPDH)、乳酸脱氢酶A (LDHA)和烯醇化酶1 (ENO1)表达上调。
MP2参与ecm受体的相互作用表明其具有潜在的促纤维化功能。随后,他们从人3dpi组织中分离培养SPP1+巨噬细胞,体外实验发现SPP1+巨噬细胞上调人成纤维细胞中I型胶原α 1链(COL1A1) mRNA水平(图5e)。为了比较跨物种,他们对小鼠肌腱粘附组织进行了scRNA-seq,从损伤指长屈肌腱的小鼠肌腱粘连模型中收集肌腱粘连组织。最后,他们从小鼠肌腱粘连组织中聚集并注释了19919只小鼠MP,包括五个不同的阶段:正常腱周组织,损伤后3、7、14和28天。鉴定出5个MP聚类,包括由SPP1标记的MP2,由FOLR2标记的MP3和硒蛋白P (SELENOP)。小鼠MP富集通路的Qusage分析表明MP2参与了ecm受体相互作用和糖酵解。
差异表达蛋白热图显示,在SPP1+巨噬细胞中,包括SPP1、FN1、COL1A1和COL3A1在内的纤维化蛋白以及包括GAPDH、三磷酸异构酶1 (TPI1)和果糖二磷酸酶1 (FBP1)在内的糖酵解蛋白上调(图6a)。差异表达蛋白的生物学过程GO分析提示SPP1+巨噬细胞中ECM组织和胶原原纤维组织上调(图6b)。差异表达蛋白的KEGG富集表明SPP1+巨噬细胞中ecm受体相互作用和糖酵解上调(图6c)。免疫荧光分析显示,在10 dpi处胶原周围存在人SPP1+巨噬细胞(图6e)。在其他阶段显示SPP1+巨噬细胞在正常肌腱周组织中稀疏,并在3 dpi时增殖。
在正常肌腱周组织中,MP3占巨噬细胞的比例最大,由于肌腱损伤后早期炎症,在3dpi时巨噬细胞显著减少(图4d)。GO富集分析表明MP3参与中性粒细胞脱颗粒过程(图5b)。对不同MP亚群中富集通路的Qusage分析显示,MP3中氧化磷酸化上调,这与抗炎巨噬细胞有关,而纤维化相关通路在MP3中很少存在(图5c)。先前的研究表明,氧化磷酸化可以减少肝纤维化。MP3氧化磷酸化标志基因集的基因富集提示细胞色素c氧化酶亚基5B (COX5B)、细胞色素c氧化酶亚基7B (COX7B)和ATP合酶膜亚基e (ATP5ME)等相关基因表达上调。KEEG分析显示MP3的内吞和产热通路富集(图5d)。MP3信号通路分析显示MAPK信号通路上调(图5g)。细胞实验证实,FOLR2+巨噬细胞表现出抗纤维化表型,从人10 dpi组织中分离培养了FOLR2+巨噬细胞,体外实验表明,FOLR2+巨噬细胞可降低人成纤维细胞COL1A1 mRNA水平(图5f)。
差异表达蛋白的热图显示,氧化磷酸化相关蛋白包括ATP合成酶F1亚基α (ATP5F1A)、ATP合成酶F1亚基β (ATP5F1B)、ATP5ME、ATP合成酶外周茎亚基d (ATP5PD)、COX5B、COX7B、细胞色素c1 (CYC1)、在FOLR2+巨噬细胞中,NDUFA4线粒体复合物相关(NDUFA4)和琥珀酸脱氢酶复合物亚基D (sdHd)上调(图6a)。差异表达蛋白KEGG的富集表明,FOLR2+巨噬细胞的氧化磷酸化水平上调(图6d),提示FOLR2+巨噬细胞的抗纤维化特性可能是通过其高度活跃的氧化磷酸化特性实现的。此外,MP3在正常肌腱周组织中占很大比例,MAPK信号通路在肌腱损伤后三个阶段均下调,而在MP3中上调,这些可能支持MP3通过MAPK信号通路抗纤维化。免疫荧光显示,在10 dpi时,以及在肌腱粘附的各个阶段,包括正常、3 dpi、10 dpi和12 wpi,胶原周围存在人FOLR2+巨噬细胞。
综上所述,肌腱粘附组织中的FOLR2+巨噬细胞起源于单核细胞,在肌腱粘附过程和肌腱损伤后修复的三个阶段均表现出抗纤维化表型,且在物种间具有保守性。氧化磷酸化或MAPK信号通路是这种抗纤维化表型的潜在通路。FOLR2+巨噬细胞有助于防止人类肌腱粘附过程中的过度纤维化。
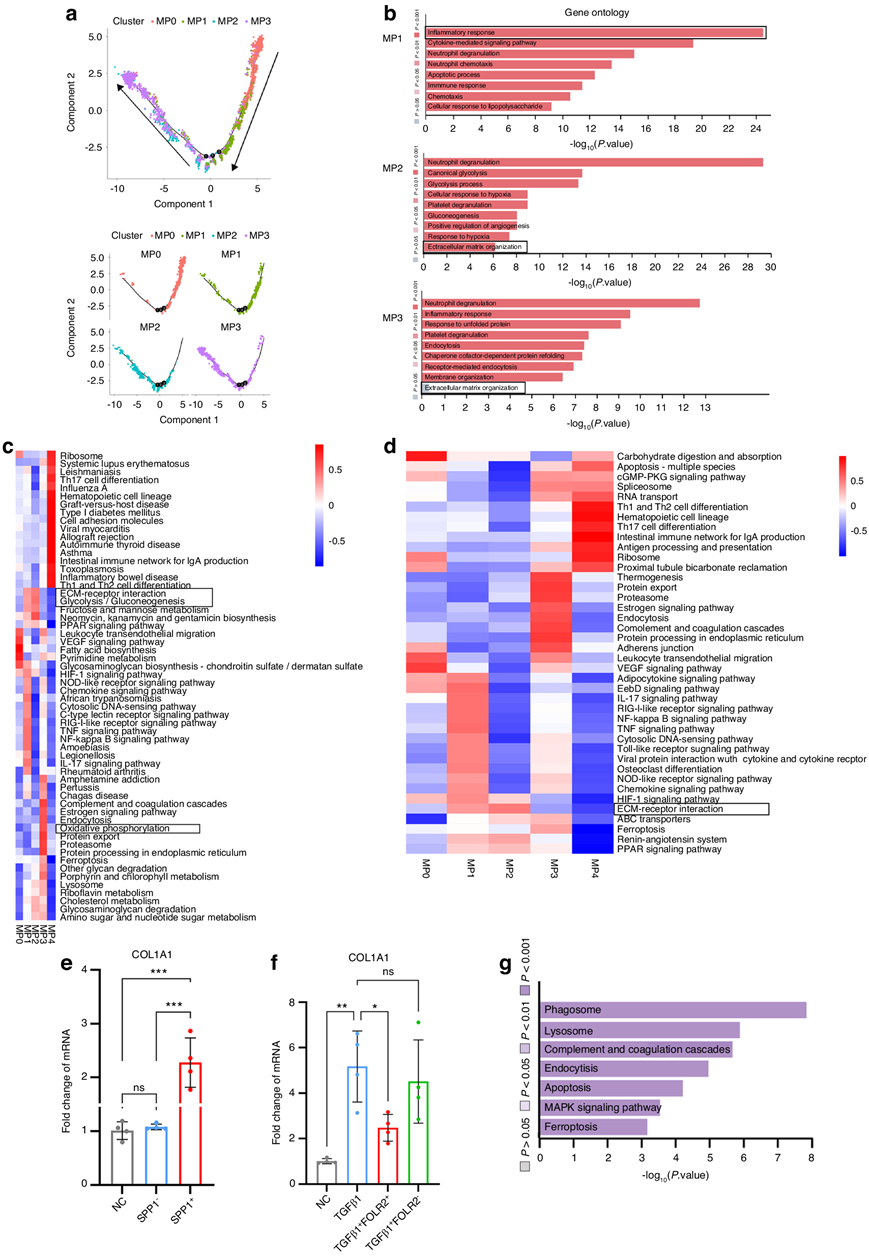
图5. 识别促纤维化巨噬细胞和抗纤维化巨噬细胞。
(a) MP0、MP1、MP2 和 MP3 的伪时间轨迹分析。(b) MP1 、 MP2 和 MP3 的GO富集分析。(c)各MP 聚类的富集通路的 Qusage 分析。(d)各MP 聚类的富集通路的 KEGG 分析。(e-f) qPCR分析。(g) MP3 的通路分析。
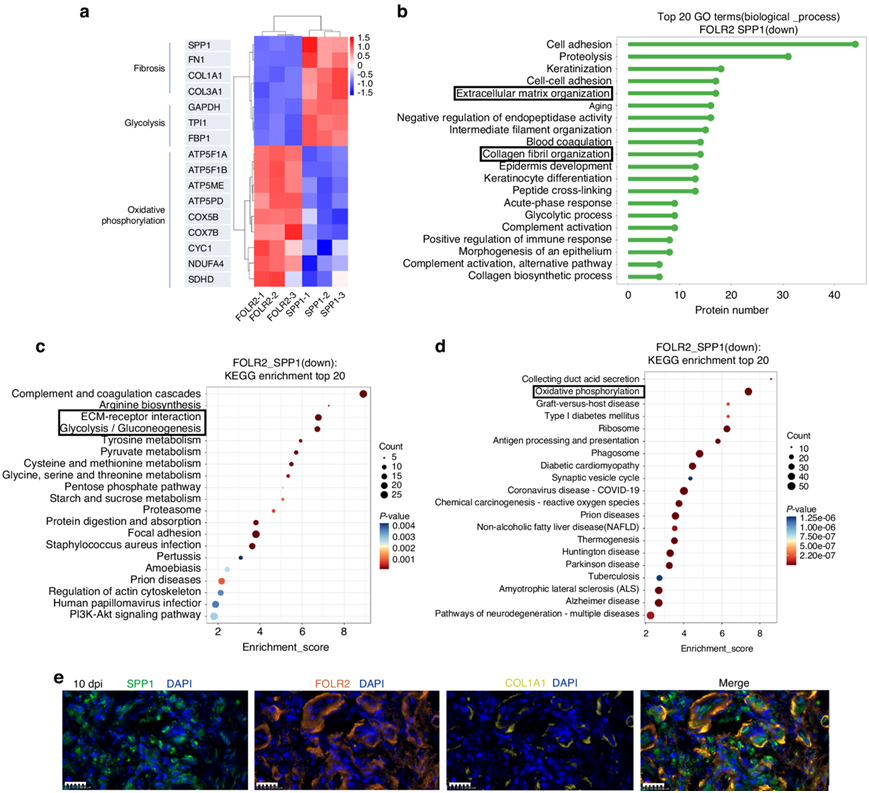
图6. SPP1 +巨噬细胞和FOLR2 +巨噬细胞的蛋白质组学和免疫荧光分析。
(a)人FOLR2 +巨噬细胞和SPP1 +巨噬细胞差异表达蛋白热图。(b)GO分析。(c-d)KEGG分析。(d) 10 dpi 人类肌腱粘连组织的代表性多色免疫荧光图像。
06
ACKR1 参与 FOLR2 +巨噬细胞迁移
最近的研究表明ACKR1可以支持单核细胞迁移。在此基础上,他们将Lysm-Cre;R26RtdTomato小鼠的骨髓移植到致命辐照的Ackr1−/−小鼠(Ackr1−/−chimeras;ACKR1缺陷)和对照小鼠(WT嵌合体)。免疫荧光分析显示,损伤后14天,Ackr1−/−嵌合体中CD68+FOLR2+单核细胞减少在此基础上,将Lysm-Cre;R26RtdTomato小鼠的骨髓移植到致命辐照的Ackr1−/−小鼠(Ackr1−/−chimeras;ACKR1缺陷)和对照小鼠(WT嵌合体)。免疫荧光分析显示,损伤后14天,Ackr1−/−嵌合体中CD68+FOLR2+单核细胞减少(图7a-d)。结果表明ACKR1参与了FOLR2+巨噬细胞在肌腱粘附中的迁移。
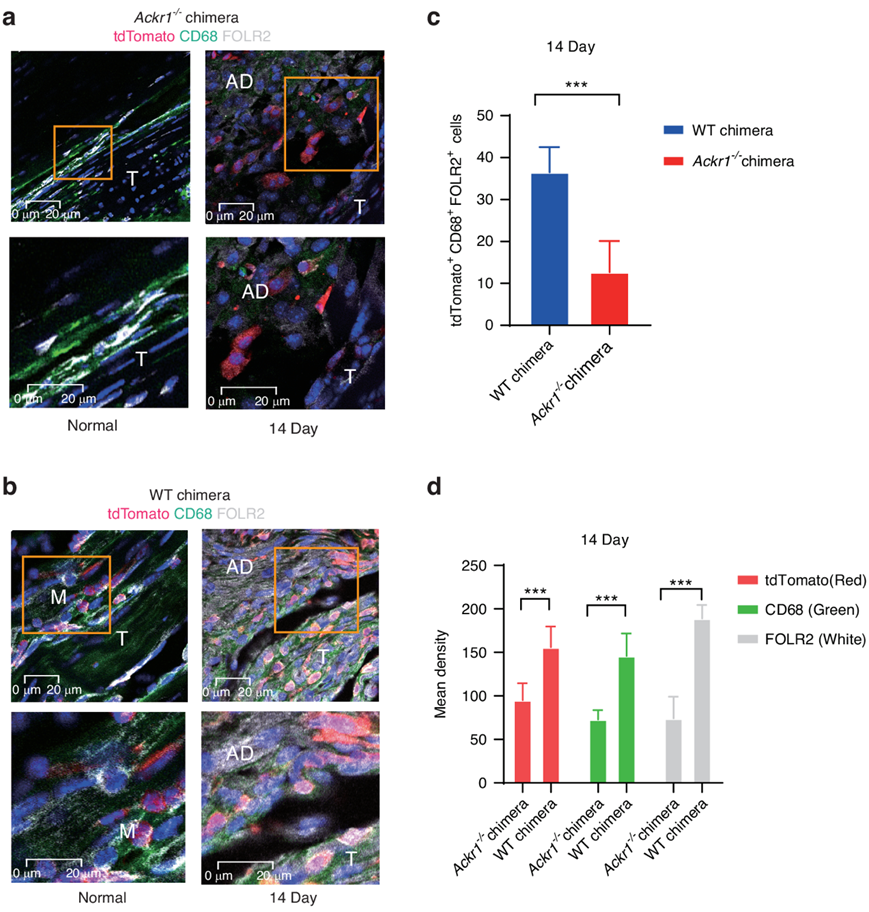
图7. 免疫荧光分析。
(a, c) 多色免疫荧光。(b,d) 平均免疫荧光密度定量。
07
腹膜周围纤维化生态位中多个细胞系之间复杂的相互作用
为了进一步探索人类肌腱粘连组织中的多谱系相互作用,他们使用 cellphoneDB 来揭示各种纤维化聚类之间的相互作用,并发现人类肌腱粘连组织中 ENDO1、ENDO2 和 MCs 之间频繁的细胞相互作用(图8a)。圆形图还显示在肌腱粘连的每个阶段 ENDO1、ENDO2 和 MC0 之间存在强烈的细胞相互作用(图8b)。MP、EC和 MC在 10 dpi 阶段与 MC2 相互作用(图8c)。同时,EC和 MC在 10 dpi 阶段与 MC3 相互作用(图8d)。具体来说,ENDO1 表达高水平的配体 delta 样典型 Notch 配体 1 ( DLL1 )、delta 样典型 Notch 配体 4 ( DLL4 )、锯齿状典型 Notch 配体 1 ( JAG1 )和锯齿状典型 Notch 配体 2 ( JAG2) ,它们与 MC0 上的NOTCH受体相互作用,包括 notch 受体 1 (NOTCH1)、notch 受体 2 (NOTCH2)和 notch 受体 3 (NOTCH3)。此外,ENDO1 表达高水平的配体PDGFB,并与 MC0 上的受体血小板衍生的生长因子受体 α (PDGFRA)和血小板衍生的生长因子受体 β (PDGFRB)相互作用(图8e)。ENDO2 表达高水平的选择素 E ( SELE),最近的研究表明它们是粘附分子,可介导白细胞在 EC 上的粘附和迁移(图8e)。MP2 和 MP3 都表达高水平的配体TGFB1(图8e)。MP2 可能通过配体SPP1、EREG和TGFB1导致人类肌腱粘附中的纤维化(图8e)。MP3 表达高水平的针对MC0 上受体IGFR的配体IGF1 ,据报道IGF1-IGFR在 MAPK 信号通路中(图8e)。MC2、MC3 与 ENDO1、ENDO2、MC0 之间的细胞相互作用显示出相似的结果(图8f-g)。有趣的是,ENDO1表达配体PDGFB, MP2表达配体TGFB1和血管内皮生长因子A (VEGFA), MC0表达配体血小板衍生生长因子D (PDGFD)和血管内皮生长因子B (VEGFB),MC2和MC3上分别有受体(图8f-g)。TGFB、PDGF和VEGF均与PMT有关。这些研究结果支持了PMT可能由RGCC+ EC、SPP1+巨噬细胞和ADAM12+成纤维细胞通过各自的途径驱动的观点。此外,MP3在ENDO2受体ACKR1上特异性表达了配体C-X-C基序趋化因子配体1 (CXCL1)(图8h),表明ACKR1可能通过配体CXCL1参与了FOLR2+巨噬细胞的迁移。
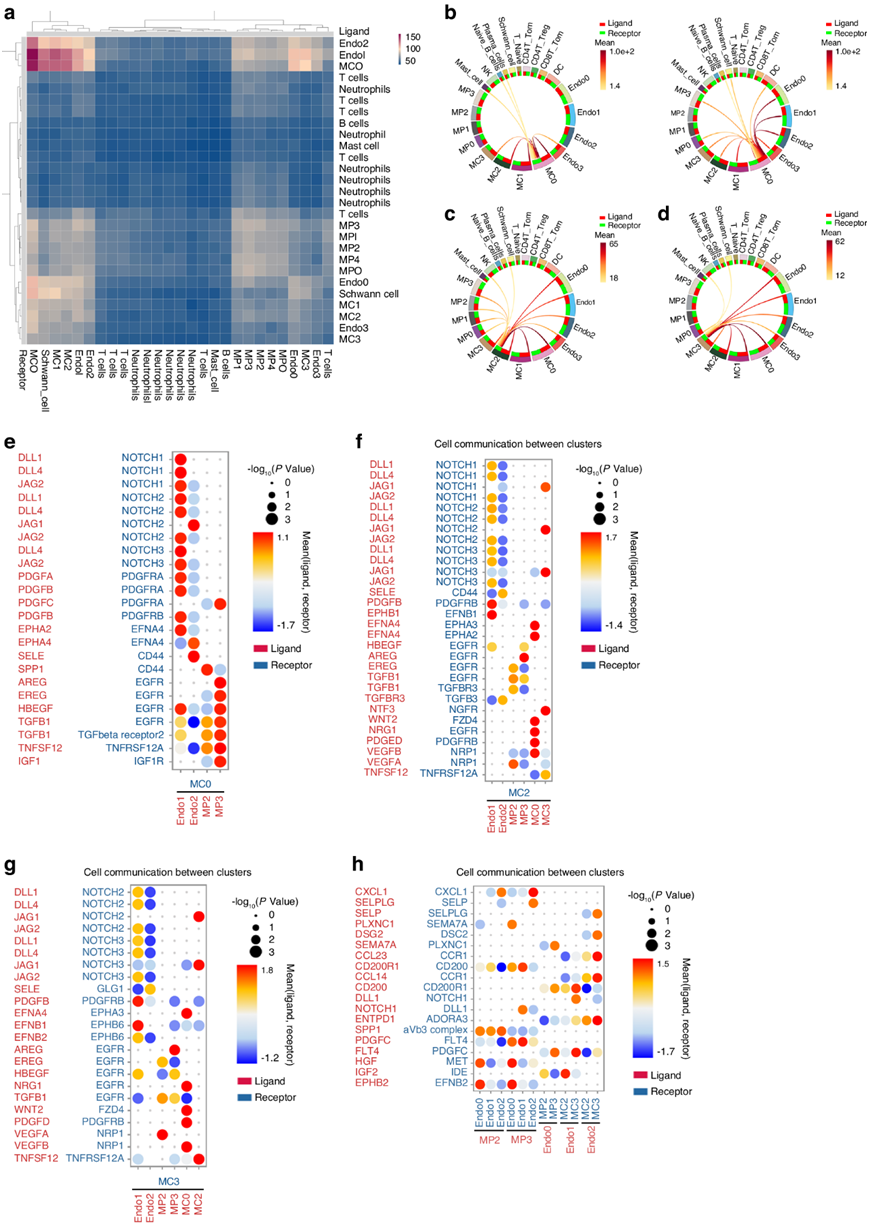
图8. 人类肌腱粘附中细胞相互作用组的特征。
(a)所有聚类之间相互作用的热图。(b) 10 dpi 下 MC0 与其他聚类之间相互作用的圆图。(c) 10 dpi 下 MC2 与其他聚类之间相互作用的圆图。(d) 10 dpi 下 MC3 与其他聚类之间相互作用的圆图。(e) MC0与 ENDO1、ENDO2、MP2、MP3 之间的配体-受体相互作用的点图。(f) MC2 与 ENDO1、ENDO2、MP2、MP3、MC3 之间的配体-受体相互作用的点图。(g) MC3 与 ENDO1、ENDO2、MP2、MP3、MC3 之间的配体-受体相互作用的点图。 (h) MP 与 ENDO 之间的配体-受体相互作用的点图。
+ + + + + + + + + + +
结 论
本项研究对肌腱损伤后不同阶段的人和小鼠肌腱粘连组织进行了多组学分析,包括scRNA-seq和蛋白质组学。对超过74000个人类单细胞的转录组进行了分析。发现 SPP1 +巨噬细胞、RGCC +内皮细胞、ACKR1+内皮细胞和 ADAM12 +成纤维细胞参与了肌腱粘连的形成。有趣的是,尽管肌腱粘连中存在特定的纤维化聚类,但使用人体细胞的体外实验将 FOLR2 +巨噬细胞鉴定为抗纤维化聚类。此外,在小鼠模型中,证实 ACKR1 可调节受伤肌腱周围部位的 FOLR2 +巨噬细胞迁移。与 WT 嵌合体相比,还观察到 FOLR2+ 巨噬细胞的减少,这表明 ACKR1 特异性参与了 FOLR2+巨噬细胞的迁移。总之,这项研究不仅通过多组学分析表征了肌腱粘连的纤维化微环境景观,而且揭示了一种新的抗纤维化巨噬细胞聚类及其起源。
+ + + + +



