

 English
English文献解读|Cell(64.5):中性粒细胞分析阐明了抗肿瘤抗原呈递效力
✦ +
+
论文ID
原名:Neutrophil profiling illuminates anti-tumor antigen-presenting potency
译名:中性粒细胞分析阐明了抗肿瘤抗原呈递效力
期刊:Cell
影响因子:64.5
发表时间:2024.03.05
DOI号:10.1016/j.cell.2024.02.005
背 景
中性粒细胞是最丰富、最有效的病原体防御者,在各种癌症类型中发挥相反的功能。然而,鉴于中性粒细胞的半衰期较短,探索中性粒细胞如何在癌症中采取特定的命运仍然具有挑战性。
实验设计
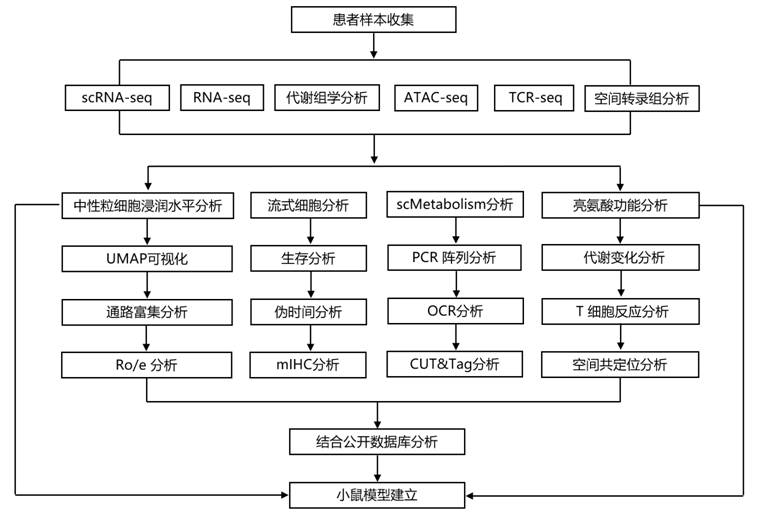
结 果
01

中性粒细胞优先浸润某些癌症类型
为了探索浸润模式并选择合适的癌症类型,研究者团队测试了 8 种常见的免疫定量算法,并分析了涵盖 31 种实体癌的 8766 个样本的癌症基因组图谱 (TCGA)。基于三种算法建立了一致的中性粒细胞浸润评分,该评分揭示了中性粒细胞浸润的强烈组织选择性模式,可分为 3 个亚型(高 29.6%、异质性 30.0% 和低 40.4%)(图 1A)。通过比较免疫亚型之间的中性粒细胞水平,他们观察到炎症或纤维化抑制性肿瘤微环境(TME)中有优先的中性粒细胞浸润(图1B),这与先前报道的中性粒细胞浸润模式和功能一致。
之后,通过单细胞转录组分析(scRNA-seq),他们生成了来自17种癌症类型143例患者的225个样本的中性粒细胞图谱(图1C)。为了分析转录特征,他们聚焦中性粒细胞,并注意到癌症和组织类型的高度异质性(图1D)。他们观察到由S100A12+、HLA-DR+CD74+、VEGFA+SPP1+、TXNIP+、CXCL8+IL1B+、CXCR2+、IFIT1+ISG15+、MMP9+、NFKBIZ+HIF1A+和ARG1+组成的10种不同的中性粒细胞状态(图1E-F)。例如,HLA-DR+CD74+亚群表现出主要组织相容性复合体(MHC)-II分子的高表达和在癌症中的普遍浸润。与中性粒细胞生物学一致,他们还确定了可能代表炎症反应(CXCL8+IL1B+)和特定趋化性(CXCR2+)特征的聚类(图1F)。
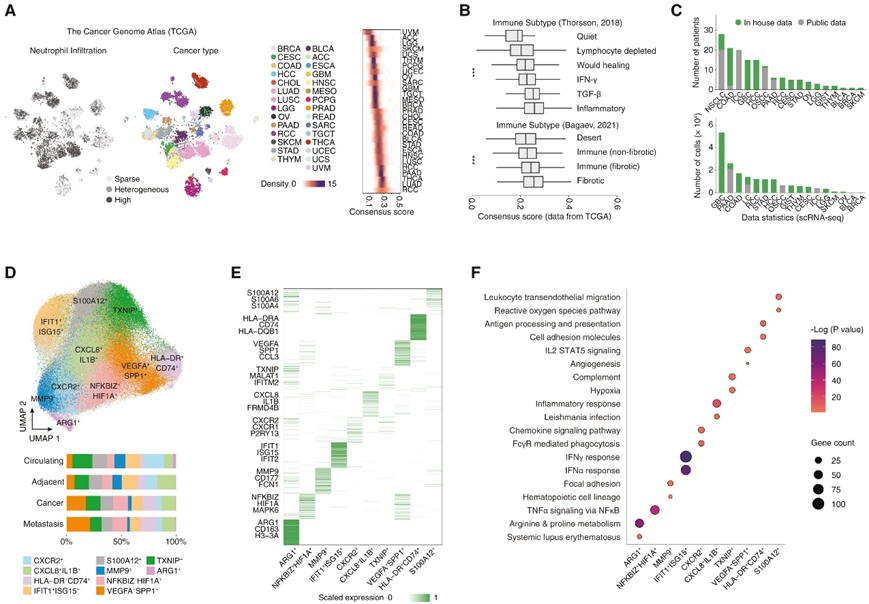
图1. 泛癌单一中性粒细胞图谱的生成。(A) 泛癌样本的中性粒细胞一致浸润水平。(B) 根据免疫亚型的中性粒细胞一致浸润水平。(C) 纳入的患者和细胞数量。(D) UMAP 图(上)和中性粒细胞比例(下)。(E) 中性粒细胞子集中的基因表达热图(前 50 个表达)。(F) 每个中性粒细胞亚群的富集通路。
02
分子分化与生存相关性
他们计算了树形结构,并观察了不同子集的分散树叶(图2A)。根据Ro/e分析(观察到的细胞数量与预期的细胞数量之比)(图2B), HLA-DR+CD74+和VEGFA+SPP1+中性粒细胞是总体上最丰富癌症的亚群(图2B),但显示出癌症类型偏好。HLA-DR+CD74+中性粒细胞在非小细胞肺癌(NSCLC)、膀胱癌(BLCA)和卵巢癌(OV)中富集,在肾细胞癌(RCC)和口腔鳞状细胞癌(OSCC)中浸润减少。相比之下,VEGFA+SPP1+中性粒细胞在NSCLC、BLCA和OV中呈稀疏浸润,但在RCC和胃腺癌(STAD)中富集。这些数据通过流式细胞分析和多重免疫组织化学(mIHC)以及使用独立的多癌tma队列[包括8种癌症类型,包括乳腺癌(BRCA)、结肠癌(COAD)、肝内胆管癌(ICC)、肝细胞癌(HCC)、STAD、NSCLC、RCC和胰腺腺癌(PAAD)]进行验证(图2C-E)。
为了探索中性粒细胞亚群与患者生存的关系,他们首先分析了基于TCGA泛癌症数据集的中性粒细胞亚群特征。在这些亚群中,VEGFA+SPP1+亚群与最差的患者预后相关,而HLA-DR+亚群与最好的患者预后相关(图2F)。在一个独立的8-癌tma队列中,他们通过mIHC进一步证实了HLA-DR+中性粒细胞与预后的相关性(图2G)。
接下来,他们比较了不同中性粒细胞亚群的细胞因子谱(图2H)。HLA-DR+CD74+中性粒细胞显示高丰度的CCL5,它可募集(recruit) T细胞。IFIT1+ISG15+中性粒细胞与PD-L1 (CD274)的高表达相关,表明其免疫抑制作用。只有HLA-DR+CD74+中性粒细胞特异性富集MHC II类分子,如HLA-DRA和HLA-DRB1(图2I)。几乎所有的中性粒细胞亚群都表达高水平的MHC I类分子,中性粒细胞亚群的免疫表型特征表现出显著的多样性(图2J),而几乎所有亚群都表现出高度的衰老特征,这证明肿瘤相关中性粒细胞大多处于成熟状态。
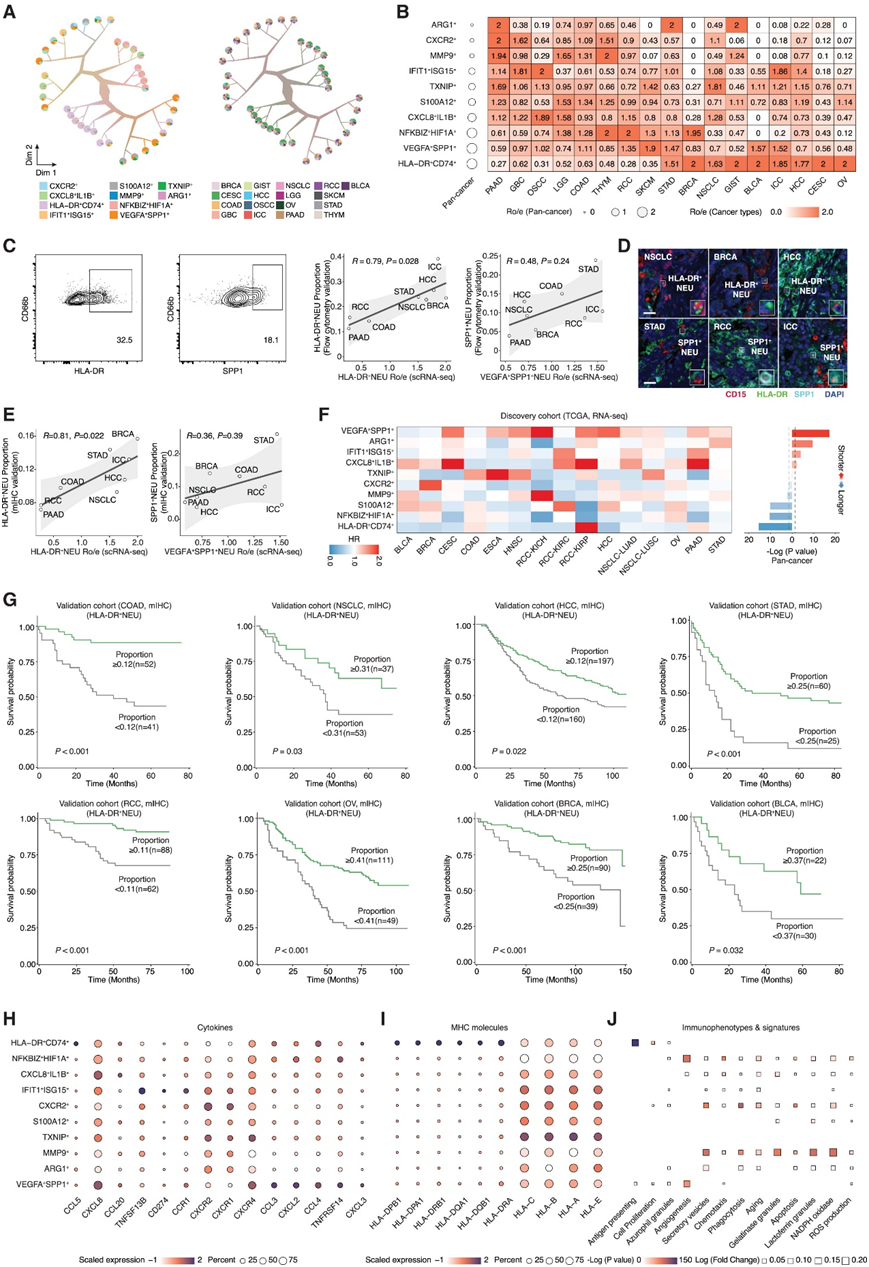
图2. 中性粒细胞的分子特征和生存相关性。
(A) 使用 TooManyCells 根据细胞亚群(左)和癌症类型(右)划分的中性粒细胞树结构。(B) 不同癌症类型中的中性粒细胞 Ro/e(观察到的细胞数与预期细胞数的比率)。(C)基于scRNA-seq和流式细胞分析以及中性粒细胞浸润的相关性结果图。(D)使用mIHC成像NSCLC、BRCA和HCC (HLA-DR+中性粒细胞富集癌型)中HLA-DR+中性粒细胞和STAD、RCC和ICC(SPP1+中性粒细胞富集癌型)中SPP1+中性粒细胞。(E)基于 scRNA-seq 和 mIHC 的HLA-DR +(左)或 SPP1 +中性粒细胞(右)浸润之间的相关性。(F) 中性粒细胞特征的预后价值。(G)生存分析。(H–J) 差异表达细胞因子、MHC 分子、免疫表型和特征的表达谱。
03
成熟和代谢状态分析
他们观察到沿中性粒细胞状态持续分化,HLA-DR+CD74+中性粒细胞的末端伪时间值最多(图3A)。根据肿瘤特异性和伪时间对中性粒细胞进行排序,发现HLA-DR+CD74+中性粒细胞可能留在末端(图3B)。为了验证这一发现,他们首先使用流式细胞分析评估了来自24例8种癌症类型患者的肿瘤内中性粒细胞中的成熟标志物CD11b和CD1636(图3C)。结果,HLA-DR+中性粒细胞亚群的CD11b和CD16表达显著升高。随后,他们利用mIHC证实了在不同癌症中HLA-DR+中性粒细胞中成熟标志物(CD11b和MPO)表达的增强(图3D-E)。
他们分析了通路方差,并观察到代谢通路的显著多样性,然后,他们量化了中性粒细胞亚群的代谢通路活性(图3F)。值得注意的是,HLA-DR+中性粒细胞显示出显著的氨基酸代谢富集(即缬氨酸、亮氨酸和异亮氨酸)(图3G)。在具有免疫抑制作用的VEGFA+SPP1+中性粒细胞中,维生素代谢和聚糖代谢的激活占主导地位。
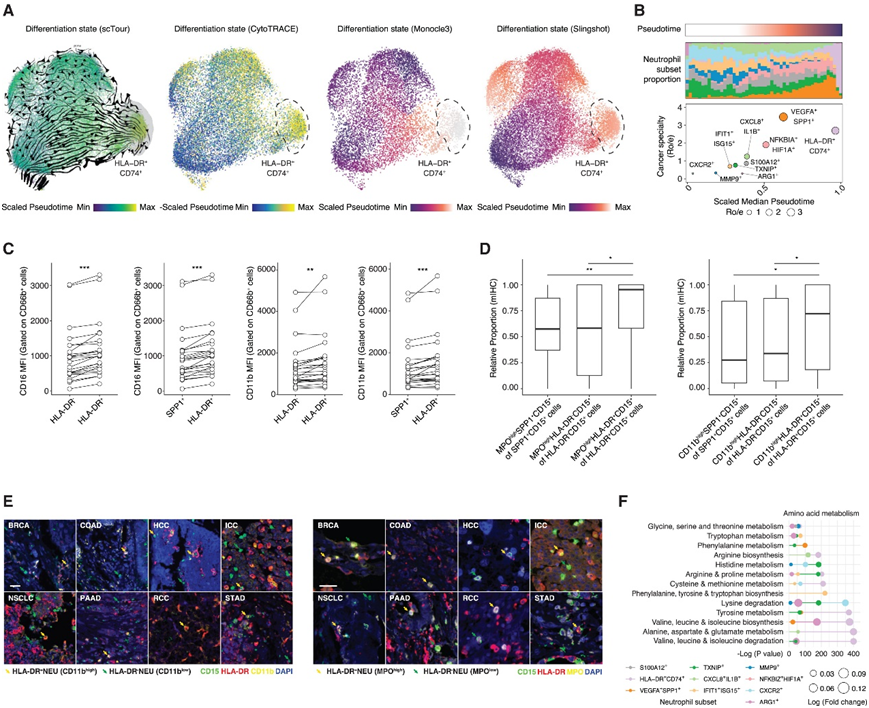
图3. HLA-DR +中性粒细胞终末分化并代谢重编程。
(A) 通过 scTour、CytoTRACE、monocle3 和 Slingshot估计的分化状态。(B) 按癌症特征 (Ro/e) 和分化状态(scTour 的伪时间)排序的中性粒细胞亚群(上)以及癌症特征 (Ro/e) 和伪时间(下)之间的相关性。(C)平均荧光强度 (MFI)分析。(D)在多癌tma队列中,使用mIHC的SPP1+CD15+、HLA-DR-CD15+和HLA-DR+CD15+中性粒细胞中CD11bhigh和MPOhigh中性粒细胞比例。(E)在多癌tma队列中使用mIHC的HLA-DR+和HLA-DR-中性粒细胞中的CD11b和MPO强度。(F) scMetabolism检测的中性粒细胞亚群氨基酸代谢通路活性。
04
亮氨酸代谢支配着抗原呈递机制的表观遗传学
为了系统地研究氨基酸对中性粒细胞的影响,他们设计了一种体外筛选策略,包括所有20种氨基酸,并研究了它们对来自健康供体的循环中性粒细胞抗原呈递的影响(图4A)。特别是亮氨酸上调HLA-DR和CD80等共刺激分子(图4B-C)。他们进一步使用PCR阵列分析。值得注意的是,亮氨酸显著促进MHC-II复合物组装的基因表达(图4D),增强抗原加工蛋白酶,促进抗原装载过程(图4E-F)。RNA-seq分析也证实了亮氨酸对MHC-II的影响,但对MHC-I没有影响(图4G)。
为了探索亮氨酸如何调控代谢组并驱动抗原呈递程序,他们进行了非靶向代谢组学研究,并观察到ATP和脂肪酸产生的强烈差异(图4H)。考虑到ATP的产生主要发生在线粒体中,他们假设亮氨酸可能通过线粒体触发功能或表型重塑。事实上,亮氨酸引起线粒体聚集和表型改变(即线粒体质量、Ca2+和ROS产生)(图4I)。为了测试亮氨酸对线粒体的动态影响,他们检测了氧气消耗率(OCR)的实时变化,并观察到亮氨酸刺激显著增强了线粒体OCR(图4J)。透射电镜(TEM)分析显示,亮氨酸处理可以诱导特定的形态学特征,如线粒体长度变长,膜上伪足增多(图4K),表明细胞间接触电位增加。
鉴于线粒体呼吸和电子传递链机制的复杂性,他们探究了亮氨酸对特定线粒体亚组分的因果影响。通过量化单个HLA-DR+中性粒细胞的线粒体呼吸特征(图4L),发现从还原性烟酰胺腺嘌呤二核苷酸(NADH)转移电子并产生烟酰胺腺嘌呤二核苷酸(NAD)的复合物I在亮氨酸处理后表现出更高的活性。他们抑制复合物I的活性,观察到线粒体膜状态和HLA-DR+中性粒细胞比例下降(图4M)。
之后,他们用13c标记的亮氨酸喂养来自健康供体血液的中性粒细胞,发现亮氨酸转化为乙酰-CoA,进入三羧酸(TCA)循环,并通过分解代谢通路产生增加的谷氨酸和谷氨酰胺(图4N)。与此一致的是,在亮氨酸处理期间,辅酶a的生物合成特征发生激活,乙酰-CoA激活因子显著上调HLA-DR(图4O)。乙酰-CoA抑制可显著降低HLA-DR水平,而其恢复可挽救HLA-DR,表明乙酰-CoA对HLA-DR的调控依赖。鉴于乙酰-CoA与组蛋白H3赖氨酸27乙酰化(H3K27ac)之间已知的联系,他们检测了总H3K27ac水平,并观察到亮氨酸处理后H3K27ac的增强(图4P),但组蛋白H3本身没有变化。此外,CUT&Tag分析显示MHC-II基因上的H3K27ac显著上调(图4Q),但H3K27me3和H3K4me3没有上调(图4R-S)。这些数据证明亮氨酸可以影响H3K27ac,从而激活MHC-II基因(即HLA-DRA和HLA-DQB1)、其转录因子和其调控元件(即MHC-II超增强子)(图4T)。

图4. 亮氨酸通过代谢表观遗传调控启动HLA-DR +中性粒细胞的生成。
(A) 20种氨基酸的体外筛选策略,探讨其与中性粒细胞免疫表型的相关性。(B)每种氨基酸刺激下的HLA-DR+中性粒细胞比例。(C) 亮氨酸组和对照组中性粒细胞的 CD80、CD86 和 CCR7 表达的比较。(D–F) 相对 RNA 表达分析。(G) 亮氨酸处理组和对照组的 RNA-seq 的 MHC II 类特征。(H) 亮氨酸处理的中性粒细胞与对照组之间的代谢比较。(I) 使用流式细胞分析检测亮氨酸处理的中性粒细胞和对照组的线粒体聚集水平。(J) 亮氨酸处理的中性粒细胞与对照之间的实时耗氧率 (OCR)。(K) 通过亮氨酸和对照组的透射电子显微镜进行线粒体成像。(L) 基于 HLA-DR+中性粒细胞的 scRNA-seq 数据的线粒体呼吸复合体特征。(M) 线粒体呼吸复合物 I 抑制降低了 HLA-DR+比例。(N)亮氨酸的13C 标记显示其分解代谢为乙酰辅酶A、TCA 循环和谷氨酰胺。(O) HLA-DR+中性粒细胞的亮氨酸乙酰辅酶A依赖性调节。(P) 对照组、亮氨酸处理组和 AcCoAa(AcCoA 激活剂)组之间的组蛋白 H3K27ac 水平。(Q) 使用 CUT&Tag 绘制亮氨酸和对照组 H3K27ac 峰的热图。(R-S)MHC-II 基因转录起始位点(TSS)上的 H3K27ac、H3K27me3 和 H3K4me3 覆盖率和得分比较。(T) 亮氨酸和对照组的 HLA-DRA 和 HLA-DQB1 位点上的 H3K27ac 修饰。
05
抗原呈递中性粒细胞在空间上与T细胞反应相关
他们接下来对匹配的肿瘤样本进行了bulk转录组分析(bulk RNA-seq),然后对免疫细胞图谱进行了反卷积,再对免疫细胞进行了聚类,并观察了模式化的中性粒细胞浸润图谱(图 5 A)。HLA-DR+中性粒细胞与广谱的抗肿瘤T细胞亚群(即CD4+效应记忆T细胞,CD8+效应记忆T细胞和CD8+中枢记忆T细胞)共定位。他们获取了来自9种癌症类型(BRCA、SKCM、CESC、COAD、OV、PRAD、LGG、HCC和RCC)的50个空间转录组学数据集,涵盖178330个位点,并计算了HLA-DR+中性粒细胞与主要免疫谱系之间的相关性。值得注意的是,在RCC样本(GSM5924040)中,CD8+ T细胞与HLA-DR+中性粒细胞表现出强烈的共定位,在OV (10x)和结直肠癌(OEP001756)样本中也观察到类似的结果(图5B)。这些结果表明抗原呈递中性粒细胞与T细胞在空间上有联系相关。
为了分析HLA-DR+中性粒细胞对T细胞的影响,他们将中性粒细胞亚群(从肿瘤中分类)与自体CD3+ T细胞(从PBMC中分类)共培养3天,表明HLA-DR+中性粒细胞可以促进T细胞表达TNFα(图5C)。他们随后在单细胞水平上量化了等位基因特异性HLA基因的表达(图5D)。HLA-DRB1、HLA-DPA1和HLA-DQB1显示出高频率,其中这些位点具有T细胞表位限制性反应(HLA-DRB1∗09:01)或细胞因子产生(HLA-DPB1∗05:01)。他们用人MHC-II抗原(gp100, 44-59;CMV, 65-71),并与自体T细胞共培养(图5E)。
值得注意的是,分选的HLA-DR+中性粒细胞引起了自体T细胞的抗原特异性反应,但比阳性对照(dc)弱。他们将HLA-DR+中性粒细胞与突变衍生的新抗原(TP53、KRAS、IDH2和BAP1)一起培养24小时,并进一步与自体CD3+ T细胞共培养(图5F)。他们观察到大多数新抗原的刺激T细胞反应,但这些反应在不同的供体中是不同的。例如,KRASG12V新抗原(mteyklvvvgavgggksaltiqli)在供体2和3中与中性粒细胞触发的T细胞激活密切相关。他们发现KRASG12D新抗原(LVVVGADGV)也导致反应性T细胞反应。针对这两种多肽,他们在HLA - DRB1∗07:01和HLA - a∗02:01型的供体中进行了共培养,其中控制中性粒细胞与T细胞的比例和多肽的浓度(图5G)。当HLA-DR+中性粒细胞与T细胞的比例为10:1时,KRASG12V或KRASG12D新抗原可激活T细胞的强效新抗原反应。通过对KRASG12V新抗原激活的T细胞进行TCR测序(TCR-seq),他们观察到HLA-DR+中性粒细胞刺激产生强反应性 TCR 基因重排(图5H-I)。
他们最终验证了在多癌tma队列中反应性T细胞(CD39+CXCL13+CD4+ T细胞和CD39+CXCL13+CD8+ T细胞)和抗原呈递中性粒细胞(HLA-DR+CD15+中性粒细胞)的共定位(图5J)。总之,这些数据表明抗原呈递中性粒细胞可以有效地产生反应性T细胞反应。
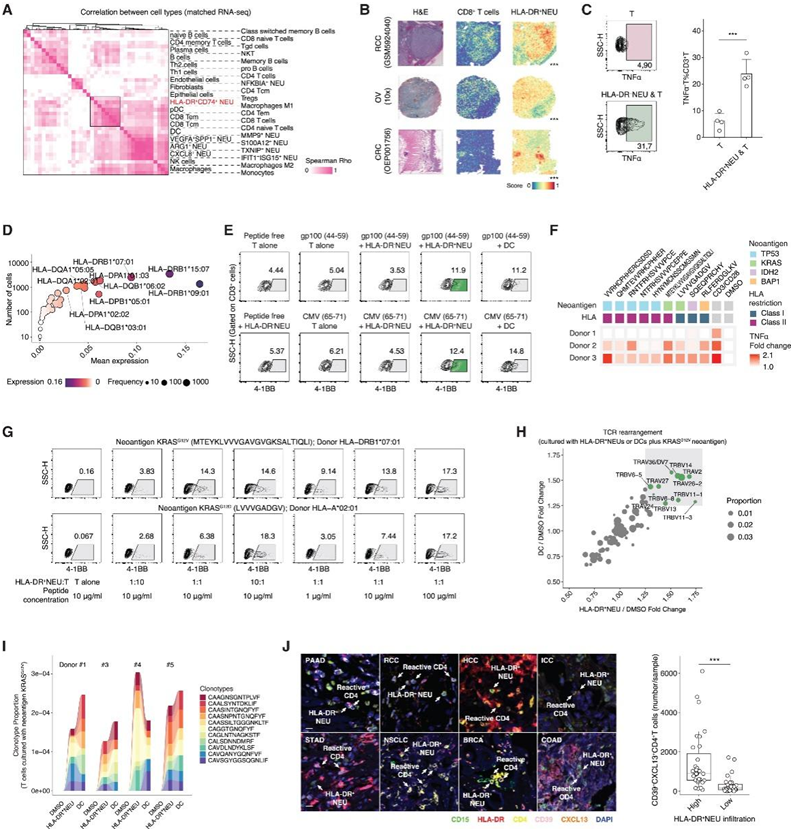
图5. HLA-DR+中性粒细胞和相关 T 细胞反应。
(A) 免疫细胞类型和中性粒细胞亚群之间的相关性。(B) RCC、OV 和 CRC 样本中HLA-DR +中性粒细胞特征与 CD8 + T 细胞之间的空间共定位。(C) 与肿瘤浸润的 HLA-DR +中性粒细胞共培养或不与肿瘤浸润的 HLA-DR + 中性粒细胞共培养的 T 细胞 TNFα 强度。(D) 基于 scRNA-seq 数据对肿瘤浸润中性粒细胞的 MHC-II 等位基因定量。(E)与亮氨酸处理的中性粒细胞(HLA-DR+中性粒细胞)、未处理的中性粒细胞(HLA-DR-中性粒细胞)、自体dc或阴性对照共培养时的T细胞反应性(4-1BB强度)。(F)与新抗原处理的HLA-DR+中性粒细胞共培养时的T细胞毒性(TNFα强度)。(G)不同新抗原KRASG12V (mteyklvvvgavgggksaltiqli)或KRASG12D (LVVVGADGV)在不同NEU:T比和肽浓度下与HLA-DR+中性粒细胞共培养时的T细胞反应活性(4-1BB强度)。(H-I) 由 HLA-DR+嗜中性粒细胞或用KRAS G12V (MTEYKLVVVGAVGVGKSALTIQLI)新抗原处理的 DC刺激的 T 细胞的 TCR 重排和 TCR 克隆型比例。(J)mIHC分析。
06
探索以中性粒细胞为基础的免疫疗法来激活TME
另一个关键问题是HLA-DR+中性粒细胞是否具有增强体内免疫治疗的潜力。为此,他们首先通过收集来自7种小鼠癌症类型的小鼠scRNA-seq数据来评估小鼠TME中中性粒细胞亚群的保护。他们在小鼠中观察到类似的亚群,如Cd74+、Spp1+Vegfa+和Isg15+中性粒细胞(图6A)。他们进一步观察了Cd74+亚群中H2-Aa(HLA-DQA同源物)、H2-Ab1 (HLA-DQB1同源物)、Cd74 (Cd74同源物)的表达和抗原呈递特征(图6B),并使用mIHC证实了这一亚群(图6C)。
鉴于亮氨酸的作用,他们在体外用亮氨酸刺激小鼠循环中性粒细胞,观察到Cd74、Cd80和Cd86的上调(图6D)。然而,来自缺乏亮氨酸转运体或分解代谢酶(Lat1KO、Bcat2KO和DbtKO)的敲除小鼠的循环中性粒细胞对亮氨酸治疗没有反应(图6D),再次证明亮氨酸在MHC-II过程中的作用。
他们随后研究了抗原呈递中性粒细胞和肿瘤表型之间的体内关联,生成MHC-IIflox/flox;Ly6GCre-tdTomato小鼠特异性缺失抗原呈递中性粒细胞(图6E),皮下注射小鼠癌细胞(LLC、MC38和Hepa 1-6),观察到肿瘤生长增加。有趣的是,肿瘤内Cd4和Cd8 T细胞在MHC-IIflox/flox;Ly6GCre-tdTomato组中的浸润均显著减少(图6F)。接下来,他们试图通过给予小鼠富含亮氨酸的饮食(水中亮氨酸含量为1.5%)来增加体内抗原呈递中性粒细胞,并观察到Cd74+中性粒细胞的增加(图6G),但这种短期饮食对肿瘤体积或体重没有影响。
在癌细胞内部,亮氨酸代谢信号也发生上调,上皮-间质转化(EMT)和tnf - α的活性发生发生上调。鉴于CD74+中性粒细胞可以激活T细胞,他们检测了T细胞的浸润,观察到其浸润增加(图6H-I)。总之,这些数据表明,短期亮氨酸饮食有利于抗tme,并诱导癌细胞表型的轻微变化,提高了其潜在的治疗用途。
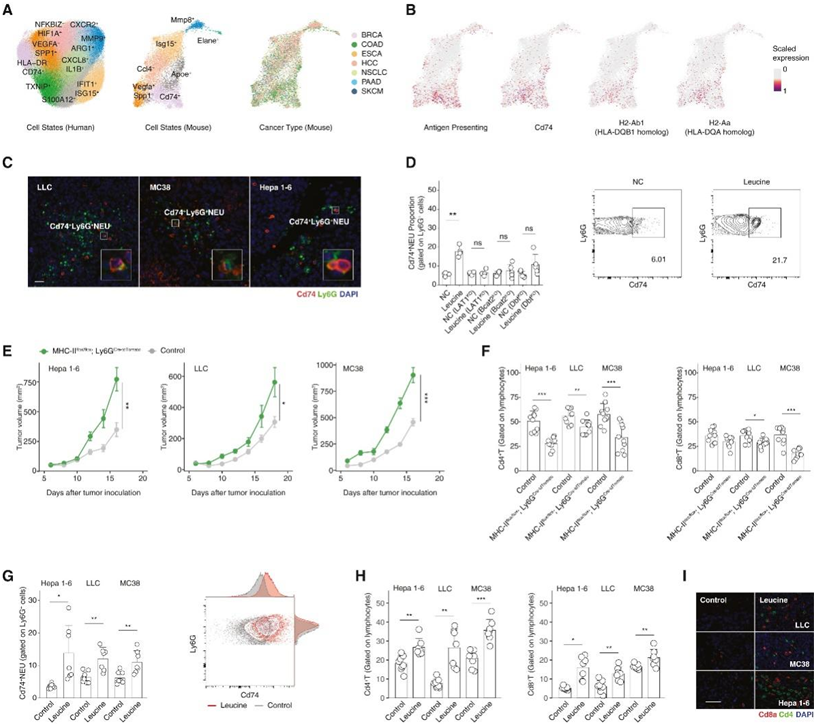
图6. 抗原呈递中性粒细胞与相关体内免疫表型之间的相关性。
(A) 根据其标记基因和癌症类型,整合小鼠肿瘤浸润中性粒细胞的 UMAP,并与人类中性粒细胞亚群进行比较。(B) 小鼠中性粒细胞的抗原呈递基因表达和特征。(C) Cd74 +中性粒细胞的代表性 mIHC 图像。(D)来自 LAT1 KO、Bcat2 KO、Dbt KO和野生型小鼠血液的亮氨酸处理和对照中性粒细胞之间的Cd74 +中性粒细胞比例。(E)肿瘤体积的比较。(F)Cd8a + T 和 Cd4 + T 细胞比例。(G) 在 LLC、MC38 和 Hepa 1-6 皮下模型中,亮氨酸饮食诱导较高的 Cd74 +瘤内中性粒细胞比例。(H) 瘤内 Cd8a + T和 Cd4 + T 细胞比例。(I)代表性 mIHC 图像。
然后,他们探讨了Cd74+中性粒细胞在小鼠癌症中的治疗作用。值得注意的是,亮氨酸饮食加抗pd -1治疗显著减少肿瘤体积,使疾病稳定(图7A)。他们还研究了抗原呈递中性粒细胞进入TME作为另一种治疗选择,分离小鼠循环中性粒细胞,用亮氨酸刺激它们,并证实它们的Cd74上调。将这些Cd74+中性粒细胞输送到肿瘤中,即使中性粒细胞数量增加到1 × 107,也能显著减小肿瘤大小,但仍不能产生稳定的疾病结果(图7B)。当转移cd74缺陷的中性粒细胞时,他们发现抗肿瘤作用明显减弱(图7B)。结合抗pd -1抗体和Cd74+中性粒细胞递送(图7C),他们在所有肿瘤模型中观察到强大的抗肿瘤效果。值得注意的是,在MC38和Hepa 1-6模型中,有相当比例的肿瘤表现出完全缓解。相比之下,cd74敲除中性粒细胞与抗pd -1联合使用的效果较弱。
在8个接受免疫治疗的队列中,包括SKCM、STAD、HCC、BLCA和NSCLC患者,他们观察到HLA-DR+中性粒细胞特征与更好的生存或反应之间存在显著的正相关(图7D-E)。接下来,他们根据患者源性肿瘤片段(PDTF)策略,在抗pd -1免疫治疗耐药的HCC样本中检测了过继性体外递送。共培养3天后,CD4和CD8 T细胞均表现出细胞毒性分子(IFNγ和TNFα)以及活性分子(4-1BB和CD39)的上调(图7F)。将抗原呈递中性粒细胞与PD-1抗体结合也会产生更大的T细胞反应性和细胞毒性(图7G)。
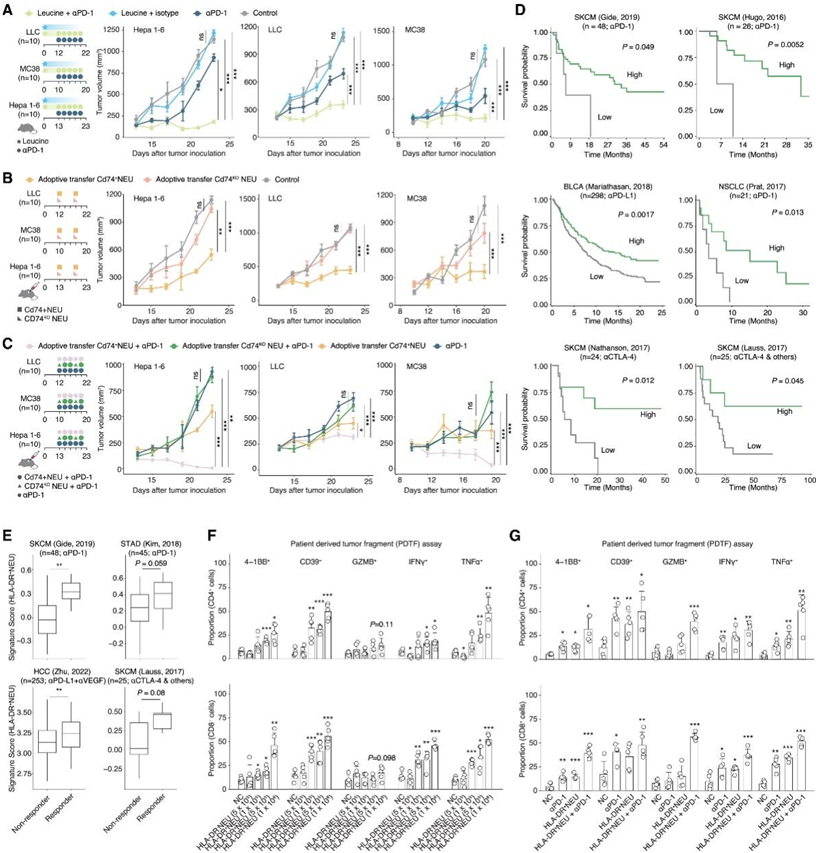
图7. 抗原呈递中性粒细胞治疗价值的分析。
(A) 亮氨酸饮食加αPD-1治疗、亮氨酸饮食加同种型治疗、单独αPD-1治疗以及对照组的肿瘤体积。(B) Cd74 +中性粒细胞过继递送、Cd74-KO 中性粒细胞过继递送和对照组中的肿瘤体积。C) Cd74 +中性粒细胞过继递送加 αPD-1 治疗、Cd74-KO 中性粒细胞过继递送加 αPD-1 治疗以及单独 αPD-1 治疗组中的肿瘤体积。(D-E) HLA-DR + CD74 +中性粒细胞特征与免疫疗法治疗的患者生存和反应模式相关。(F-G) CD4+细胞(上)和CD8+细胞(下)中4-1BB+、CD39+、GZMB+、IFNγ+和TNFα+亚群的比例。
+ + + + + + + + + + +
结 论
本项研究生成并整合了 17 种癌症类型(来自 143 名患者的 225 个样本)的单细胞中性粒细胞转录组。中性粒细胞表现出非凡的复杂性,具有 10 种不同的状态,包括炎症、血管生成和抗原呈递。值得注意的是,抗原呈递程序与大多数癌症的良好生存相关,并且可以由亮氨酸代谢和随后的组蛋白 H3K27ac 修饰引起。这些中性粒细胞可以进一步激发抗原特异性和抗原非依赖性 T 细胞反应。中性粒细胞输送或亮氨酸饮食可微调免疫平衡,从而增强各种小鼠癌症模型中的抗 PD-1 治疗。总之,这些数据不仅表明中性粒细胞在癌症之间的差异,而且还表明了治疗机会,例如抗原呈递中性粒细胞递送。
+ + + + +



