

 English
English文献解读|Cell Rep(8.8):衰老细胞模型的转录组与多组织患者样本的整合揭示了 COL6A3 作为衰老诱导剂的减少
✦ +
+
论文ID
原名:Integration of transcriptomes of senescent cell models with multi-tissue patient samples reveals reduced COL6A3 as an inducer of senescence
译名:衰老细胞模型的转录组与多组织患者样本的整合揭示了 COL6A3 作为衰老诱导剂的减少
期刊:Cell Reports
影响因子:8.8
发表时间:2023.11.07
DOI号:10.1016/j.celrep.2023.113371
背 景
细胞衰老表现为稳定的生长停滞、染色质重塑、自噬增加以及衰老相关分泌表型(SASP)的出现。衰老细胞是年龄依赖性心血管组织功能障碍的主要贡献者,但缺乏对其体内细胞标记和组织背景的了解。
实验设计
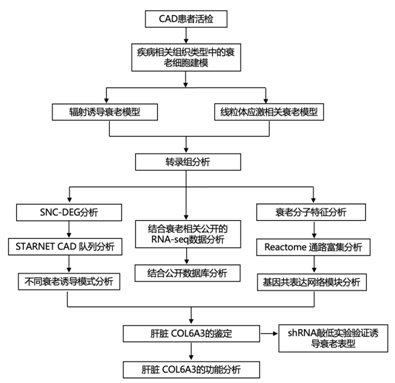
结 果
01

衰老应激引起的转录变化很大程度上是细胞类型特异性的
研究者团队建立了多种衰老的体外模型,并进行转录组分析(RNA-seq),随后又分析了 10 种与衰老诱导相关的不同差异基因表达特征(图 1 A)。IMR90细胞是一种正常的人胎肺成纤维细胞系,他们使用各种应激源诱导细胞衰老,包括X射线照射、致癌RasV12转化或用博来霉素、鱼藤酮、抗霉素或寡霉素(线粒体应激)处理。辐射用于诱导其他四种细胞系的衰老,包括RPTEC(人肾近端小管上皮细胞)、HUVEC(人脐静脉内皮细胞)、HMVEC(人类肺微血管内皮细胞)和ADSC(人脂肪组织衍生干细胞)(图1A)。他们使用对衰老细胞进行转录组分析(RNA-seq),然后分析差异表达基因(DEG)。他们将DEG定义为与静止或DMSO处理的对照细胞相比,根据调整后的p值排序的最显著上调和下调的蛋白质编码基因,最多有1250个上调和1250个下调基因(图 1 A)。
10 个衰老细胞 DEG 的交叉特征比较揭示了 8 个共同基因且显著重叠(图 1 B)。共有DEG 数量最多 (472) 的是使用寡霉素、抗霉素或鱼藤酮处理(线粒体应激)诱导衰老的 IMR90 细胞(图1C)。比较单一细胞类型 (IMR90) 内所有衰老诱导模式,他们发现累 147 个共有基因(图 1 C)。在具有相同衰老诱导模式的五种细胞类型中发现了最少数量的共有基因(X 射线照射)(图1 C)。Jaccard相似性测量和DEG重叠评估显示,同一细胞类型(IMR90)内不同应激源诱导的衰老分子谱比相同应激源(x射线照射)诱导的不同细胞类型的衰老分子谱更相似(图1D)。通过与公开的数据进行比较,他们确认了衰老转录组(图 1 E)。
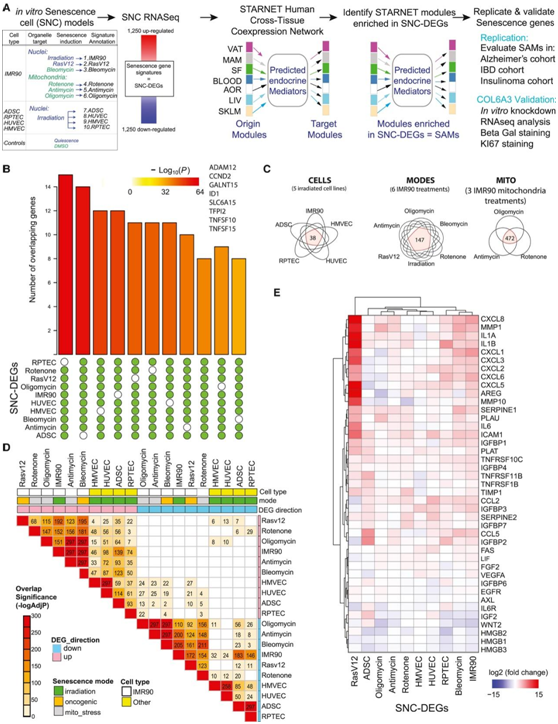
图1. 衰老的体外分子特征分析。
(A) 实验流程的示意图。(B) 在至少 9/10 或 10/10 衰老 DEG 中的 UpSet 图。(C) 维恩图总结了 X 射线照射细胞 (“CELLS”)、线粒体应激相关衰老 (“MITO”) 和衰老 IMR90 细胞在多种衰老诱导模式 (“MODES”) 中常见的 SNC-DEG 基因数量”)。 (D) 热图显示了在不同衰老诱导模式下差异表达的基因重叠的显著性。(E) 热图总结了从文献中筛选出的与衰老相关的基因的 10 个衰老 DEG 的倍数差异。
他们进一步将本项研究的衰老 DEG 与公开发表的RNA-seq数据进行了比较。在三种类型的刺激(辐照、复制抑制或Ras癌基因诱导)中,成纤维细胞基因的表达随衰老而改变(图2A)。这组上调的基因在所有上调的IMR90成纤维细胞衰老模型DEG中显著富集,与下调的DEG观察结果相似。有趣的是,本项研究鉴定衰老诱导模式中IMR90成纤维细胞中通常改变的基因,也在先前报道的成纤维细胞S1E特征中富集,这进一步证明了细胞类型内的衰老程序具有许多共性,无论在衰老诱导模式下(例如致癌与线粒体应激源)。
在本项研究的衰老HMVEC、HUVEC、ADSC和RPTEC的转录组中,富集的数量最少,包括称为“通用特征”(universal signature)(图2A中标记为universal S2F)的基因集(三个细胞系中的三个应激源)。一个例外是“复制性衰老特征”(在图2A中标记为ReplSen S1C),该特征在来自ADSC、RPTEC和IMR90细胞的衰老DEG中显著富集,提示复制性衰老和辐射诱导的分子表型之间有共性。相比之下,最近发表的基因集“SenMayo”特征(在图2A中标记为SenMayo)在对衰老细胞清除有反应的组织和物种中随着衰老而增加,并且在本研究上调的SNC-DEG特征系列中的所有基因中都有富集,证明了一个更普遍的特征。
接下来,他们评估了作为上调和下调基因集分离的10个衰老DEG中富集的Reactome通路(图2B)。细胞外基质组织和胶原形成通路在衰老DEG特征中普遍富集,相比之下,这些通路通常只富集与炎症相关的上调基因,如干扰素信号,而富集的下调衰老 DEG 的通路主要包括细胞周期控制。
一些通路是衰老诱导的特有通路,例如 IMR90 衰老 DEG(X 射线照射)中的胆固醇生物合成以及衰老 HMVEC 和 HUVEC 中的“rRNA 加工”通路。总的来说,这些结构这些数据代表了体外衰老最全面的分子检测,证明了衰老的细胞类型共性和特定分子特征。
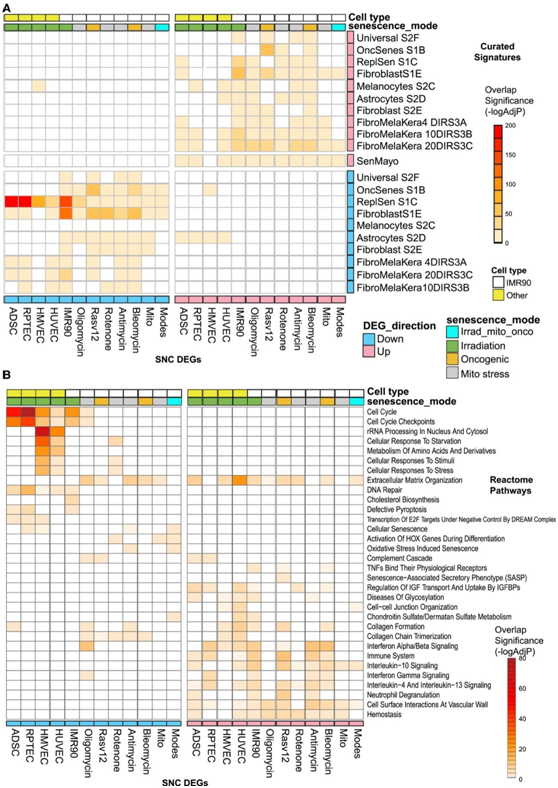
图2. 衰老分子特征对比分析和通路富集分析。
(A)公开的衰老相关差异表达基因集中 10 个衰老 DEG 显著富集的热图。(B) Reactome 通路中 10 个 DEG 的显著富集的热图。
02
将衰老相关程序映射到人类 CAD 多组织基因共表达网络
基于衰老 DEG 与衰老相关的疾病基因的联系,他们接下来的目标是通过检测从 STARNET 人类多组织共表达网络中识别的基因模块中每个衰老 DEG 的富集来确定体内相关的衰老程序。这些网络是从接受 CABG 的活体患者的基因表达遗传学 STARNET 研究中推断出来的。在患者手术期间收集了代表血液、代谢和动脉壁的七个组织样本用于基因组分析(图1A)。为了识别多组织基因共表达,将 WGCNA 算法联合应用于所有七个组织的转录组数据,总共产生了 131 个组织特异性(TS)和93 个跨组织(CT) 基因共表达网络模块。总共有 56 个 STARNET 共表达模块在衰老模型 DEG 中显著富集,并且研究者团队将他们注释为衰老相关模块(SAM)(图3 A)。
24 个 SAM 为 CT,32 个为 TS。一些 SAM 富含 10 个衰老模型 DEG 特征中的 10 或 9 个(例如CT 模块 28_LIV 和 15_VAF,图3 A),而其他 SAM 仅富含少数特征(例如,IMR90 和 HUVEC 中的 TS 肝脏模块 98)衰老DEG)。因此,实验性衰老细胞系统的分子程序组成部分通常在各种人体组织类型中共表达,并反映模式和细胞特异性的衰老。
他们重点关注 SAM_CT.28_LIV 模块中既是预测关键驱动因素又是内分泌因子(即 SAM-SASP 因子)的基因。这是最突出的 SAM,在所有衰老模型 DEG 中富集,表明它具有衰老程序的核心特征,与应激源或细胞类型无关,并通过全基因组关联分析研究(GWAS)富集与冠心病(CAD) 因果相关。从与 SAM_CT.28 相关的内分泌介质列表(即 16 个 SAM-SASP 基因)开始,他们优先考虑那些关键驱动因素。其中,他们优先考虑胶原蛋白 6A3 (COL6A3),因为它在 7个衰老 DEG中下调(图3 B),并且作为内分泌介质,其预测靶模块数量最多(图 3 C)。预测COL6A3靶向的模块/组织包括219.Blood、37.SF和 61.Blood模块也在衰老模型DEG中富集(图3D)。COL6A3是细胞外基质(ECM)的三种主要胶原VI多肽之一,它是一个SAM-SASP因子,并且具有一种内啡肽的可溶性裂解产物。
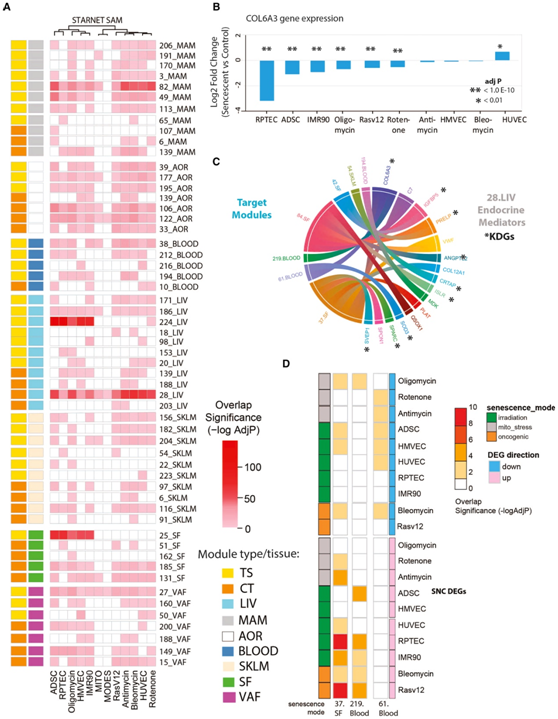
图3. STARNET CAD队列共表达网络中衰老模型DEG的富集,识别体内衰老相关基因。
(A) 热图总结了衰老 DEG 中显著富集的跨组织 (CT) 和组织特异性 (TS) 模块。鉴定出 56 个衰老相关模块。(B) 衰老诱导对 28.CT_LIV SAM 和 COL6A3 的预测关键驱动基因表达的影响。(C) 显示 SAM、28.CT_LIV 及其目标模块的预测内分泌介质的和弦图。(D) 热图总结了三个 STARNET 模块(37_SF、219_Blood 和 61_Blood)中 10 个衰老 DEG 的富集重要性。
03
证实COL6A3敲低在成纤维细胞中诱导衰老样表型
与 SAM_CT.28_LIV 基因相关的生物过程包括细胞外基质组织、IGF 结合蛋白的调节和糖胺聚糖代谢(图 4A)。有趣的是,他们发现了大量与心脏健康和功能相关的通路,例如扩张型心肌病、弹性纤维形成和肌肉收缩。鉴于 COL6A3 在 CAD 患者与健康对照中的基因表达差异,他们推测这些通路可以反映肝脏中的纤维化过程。事实上,SAM_CT.28_LIV在与肝硬化相关的细胞类型标记物中显著富集。
接下来,他们检测了 COL6A3 作为潜在 SASP 因子的 CT 分子结果,并注意到肝脏 COL6A3 的表达与皮下脂肪模块 37.TS_SF 的表达呈负相关,而与两个血液模块 61.TS_BLOOD和219.TS_BLOOD呈正相关。尽管存在相关性,但白细胞介素-4 (IL4)和IL13信号传导在所有三个目标模块中均显著富集(图4A)。由于这些Th2细胞因子是纤维增殖性疾病发病机制中的关键介质,这表明肝脏COL6A3也可能参与CT对伤口愈合和脂肪和血液中的纤维化过程的调节。
为了验证COL6A3在衰老中的作用,他们建立了COL6A3表达降低的稳定转导的IMR90成纤维细胞(COL6A3down)(图4B),并测试了COL6A3down细胞的转录谱是否模拟了各种实验性衰老模型的转录谱。事实上,COL6A3down的DEG(1102个上调,930个下调)在大多数衰老DEG中显著富集。一般来说,随着COL6A3表达缺失而上调或下调的基因与衰老DEG中上调或下调的基因相似,这验证了COL6A3减少与促衰老细胞表型相关。利用COL6A3down和对照shRNA的转录组生成的衰老DEG的基因集变异分数显示了类似的趋势,在衰老模型中,COL6A3down的上调基因的分数普遍高于对照shRNA。此外,COL6A3down细胞还表现出衰老相关β半乳糖苷酶染色的增加和Ki-67染色的减少,表明减少COL6A3表达诱导了一般的衰老样分子和生化表型。
接下来,他们检测了所有 STARNET 模块中 IMR90 或肌成纤维细胞 COL6A3downDEG 的富集情况,并重点关注 STARNET 模块的子集,其中 COL6A3 是预测的KDG (CT.28_LIV) 或预测作为内分泌介质(61_Blood、219_ Blood和37_SF)的靶标。他们还在219_Blood模块中观察到与COL6A3敲低相关的上调DEG重叠,以及在61_Blood和37_SF模块中下调的DEG重叠,证明COL6A3敲低可能影响这些基因的表达(图4D)。总的来说,IMR90细胞中辐照下调和上调的基因也在SAM_28.CT_LIV,证明COL6A3敲低“模拟”辐照诱导的衰老。随后,他们发现COL6A3down DEG和SAM_CT.28_LIV之间存在富集通路的重叠(图4A)。COL6A3下调导致胶原生物合成和修饰酶通路中的相关基因富集。
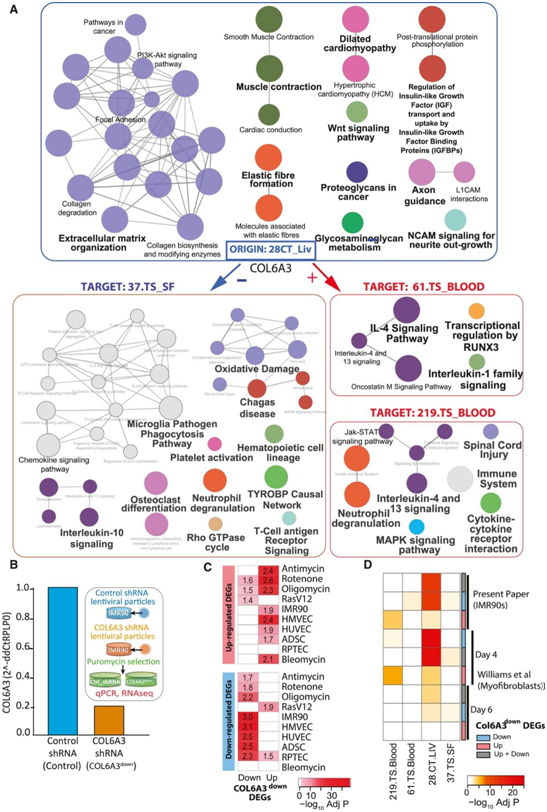
图4. COL6A3是预测的28.CT_LIV SAM的关键驱动基因和衰老相关基因。
(A) ClueGO 对 28.CT_LIV 模块基因和来自三个肝脏 COL6A3 靶模块 37.TS_SF、219.TS_BLOOD 和 61.TS_Blood 的基因进行的通路富集分析。(B) 通过 qRT-PCR 将 COL6A3 表达标准化为 RPLP0,显示在 IMR90 细胞中短发夹 RNA (shRNA) 靶向 COL6A3 敲低后,COL6A3 表达减少约 5 倍。(C-D) 热图显示了 IMR90 细胞中 COL6A3 敲低后 DEG 富集相对于衰老 DEG 或从 (A) 中选择 SAM 的重要性。
04
COL6A3敲低后IMR90成纤维细胞的基因表达变化揭示DREAM复合物在调节衰老细胞周期中的作用
COL6A3down的上调和下调的DEG在“癌症中的衰老和自噬”通路中均有富集,他们观察到SASP因子IGFBP5和IGFBP7在COL6A3down中下调,而IGFBP3上调(图5A)。鉴于此,他们评估了在IMR90实验中COL6A3的抑制影响已知的SASP基因的表达。通过整理的SASP基因列表,他们发现在COL6A3down下调和上调的DEG中,SASP因子基因显著富集,这与在衰老DEG中观察到的SASP因子普遍上调形成对比。因此,COL6A3的下调部分促进了衰老过程中典型的SASP因子诱导,另外还有一些分子触发器对SASP因子诱导具有重要作用(图1A)。
令人惊讶的是,尽管COL6A3down DEG显著富集,但细胞周期过程与SAM_CT.28_LIV基因无关(图5A)。细胞周期控制是衰老细胞的一个重要表型,这使他们假设COL6A3可能与SASP之外的其他衰老表型相关。
他们推测COL6A3可能通过肝脏内的自分泌效应介导细胞周期控制,检测了其他肝脏模块,以确定COL6A3downDEG的富集情况,并确定了SAM_224.TS_LIV富集了在COL6A3down下调的基因(图6A-B)。224.TS_LIV也是一个SAM (SAM_224.TS_LIV),在大多数衰老模型DEG中显著富集下调的基因(图6C)。SAM_224.TS_LIV 和衰老DEG的Hallmark富集分析证实了细胞周期调控过程,如E2F和myc靶标以及G2M检查点控制。有趣的是,在IMR90细胞中,两个细胞周期进程的调节因子,即MYBL2和FOXM1 (G2/M),随着COL6A3的下调而下调(图5A)。这些因子与MuvB形成复合体,MuvB是一个核心复合体,也存在于由RB样蛋白p130, E2F4和DP1(DP、RB、E2F和MuvB)组成的第三个转录复合体中,称为DREAM复合体,它的功能是限制细胞周期的再进入。他们观察到DREAM靶基因在SAM_224.TS_LIV中富集。在IMR90细胞和肌成纤维细胞中,DREAM靶基因随着COL6A3敲低,基因表达下调(图5B)。
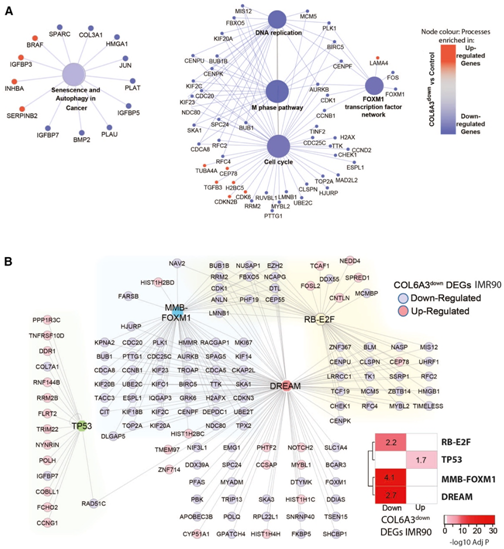
图5. IMR90 成纤维细胞中 COL6A3 敲低后的基因表达变化揭示了 DREAM 复合物在调节衰老细胞周期中的作用。
(A) “衰老、自噬和癌症”和“细胞周期”通路分析。(B) DREAM 复合物靶向 G1/S 和 G2/M 细胞周期基因,这些基因包含受 RB-E2F 和 MMB-FOXM1 以及 TP53 调节的不同亚组。
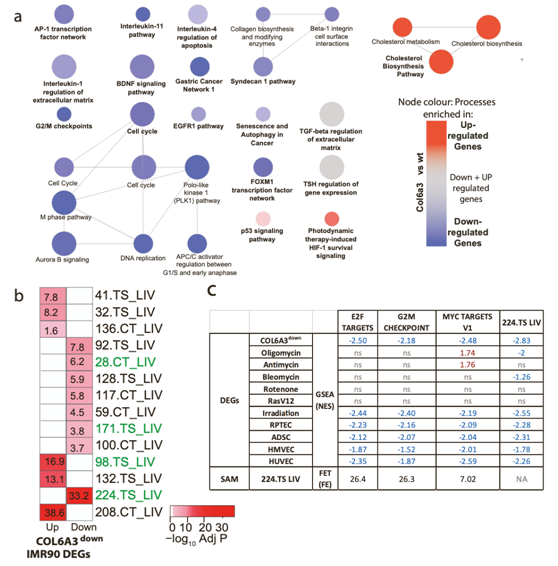
图6. COL6A3down DEG生物富集分析。
(A) 根据ClueGO的KEGG、BioPlanet和Wiki Pathway数据库进行分析。 (B) 热图总结了IMR90-COL6A3 DEG中肝脏模块的富集。(C) 根据Hallmark通路和肝脏SAM,与细胞周期相关的各种基因集的基因集富集分析(GSEA)结果。
+ + + + + + + + + + +
结 论
本项研究将 10 个实验性衰老细胞模型的转录组与基于约 600 名冠状动脉疾病 (CAD) 患者的 7 个组织活检的 RNA-seq 数据的 224 个多组织基因共表达网络进行了整合,鉴定了 56 个与衰老相关的模块,其中许多富含 CAD GWAS 基因并与心脏代谢特征相关,这证明了衰老基因程序在跨组织和 CAD 中的普遍性。跨组织网络分析揭示了 86 个候选衰老相关分泌表型 (SASP) 因子,包括 COL6A3。COL6A3 的实验性敲除会诱导与大多数实验性衰老模型重叠的转录变化,其中细胞周期停滞与 DREAM 复合物靶向基因的调节有关。本项研究提供细胞衰老的转录组数据,并鉴定了候选生物标志物、SASP 因子和人体组织衰老的潜在驱动因素。
+ + + + +



