

 English
English文献解读|Cell(64.5):使用单细胞转录组学和蛋白质组学成像绘制人类骨髓生态位的细胞生物地理学
✦ +
+
论文ID
原名:Mapping the cellular biogeography of human bone marrow niches using single-cell transcriptomics and proteomic imaging
译名:使用单细胞转录组学和蛋白质组学成像绘制人类骨髓生态位的细胞生物地理学
期刊:Cell
影响因子:64.5
发表时间:2024.05.02
DOI号:10.1016/j.cell.2024.04.013
背 景
骨髓是一个复杂的器官,含有造血细胞、间充质细胞、内皮细胞、血管平滑肌细胞和神经谱系的多种细胞。已知相对罕见的非造血细胞对造血作出重要贡献。许多非造血细胞类型,包括内皮细胞(EC)、间充质基质细胞(MSC)和成骨细胞,是骨髓微环境的关键组成部分。然而,人类骨髓中这些细胞的异质性和空间组织在很大程度上仍然未知。
实验设计
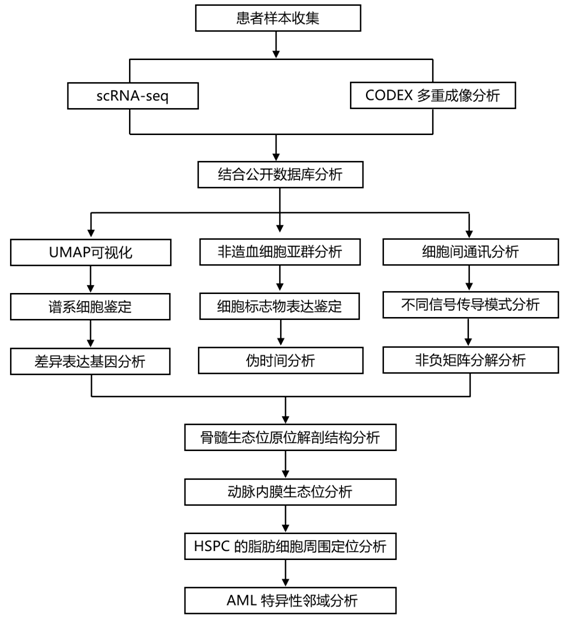
结 果
01

人类骨髓的综合 scRNA-seq 图谱
为了揭示人类骨髓的细胞组成,研究者团队开发了一个实验分选模式,用于从股骨头组织中酶促释放细胞(图S1A)。他们通过微计算机断层扫描分析确认感兴趣区域具有正常的小梁结构(图S1B)。接下来,他们设计了一种富集策略,可以捕获造血细胞、稀有造血干细胞和祖细胞 (HSPC) 以及非造血微环境细胞(图 1 A)。将这三个群体按照有利于稀有 HSPC 和非造血细胞代表性的比例合并,然后进行单细胞转录组分析(scRNA-seq)(图 1 A)。他们总共对 53417 个造血细胞和 29325 个非造血细胞进行了测序(图 1B-C,图S1C-D)。
他们首先分析了造血细胞群体,正如预期的那样,确定了三系造血和 HSPC 亚群(图 1D)。通过他们的样本处理方法,改进了其他以造血为重点的骨髓scRNA-seq参考数据,捕捉到了完整的粒细胞分化轨迹。这是因为本项研究的样本处理方法没有包括基于密度的单核细胞分离——一种标准的临床组织保存过程,由于粒细胞具有多叶核,通常会导致它们的丢失(CSF3R)(图1B)。捕获到了大量浆细胞,年龄并没有影响捕获细胞类型的频率(图1B,图S1E)。
接下来,他们分析了非造血细胞区室,确定了三种主要细胞谱系——血管平滑肌细胞 (VSMC)(ACTA2、RGS5和TAGLN)、EC(CDH5、PECAM1和VWF)和间充质细胞(CXCL12和PDGFRA),包括 MSC 和成骨细胞(图1B-D)。与现有的 scRNA-seq 公开数据相比,骨髓中这些细胞的细胞多样性要高得多(图S1F)。值得注意的是,与早期的研究结果相反,早期的研究结果将 MSC 描述为具有脂肪细胞转录谱的相对同质性,他们发现这些酶释放的细胞具有高度异质性,并鉴定出许多聚类(图 1 B 和图 S1F)。
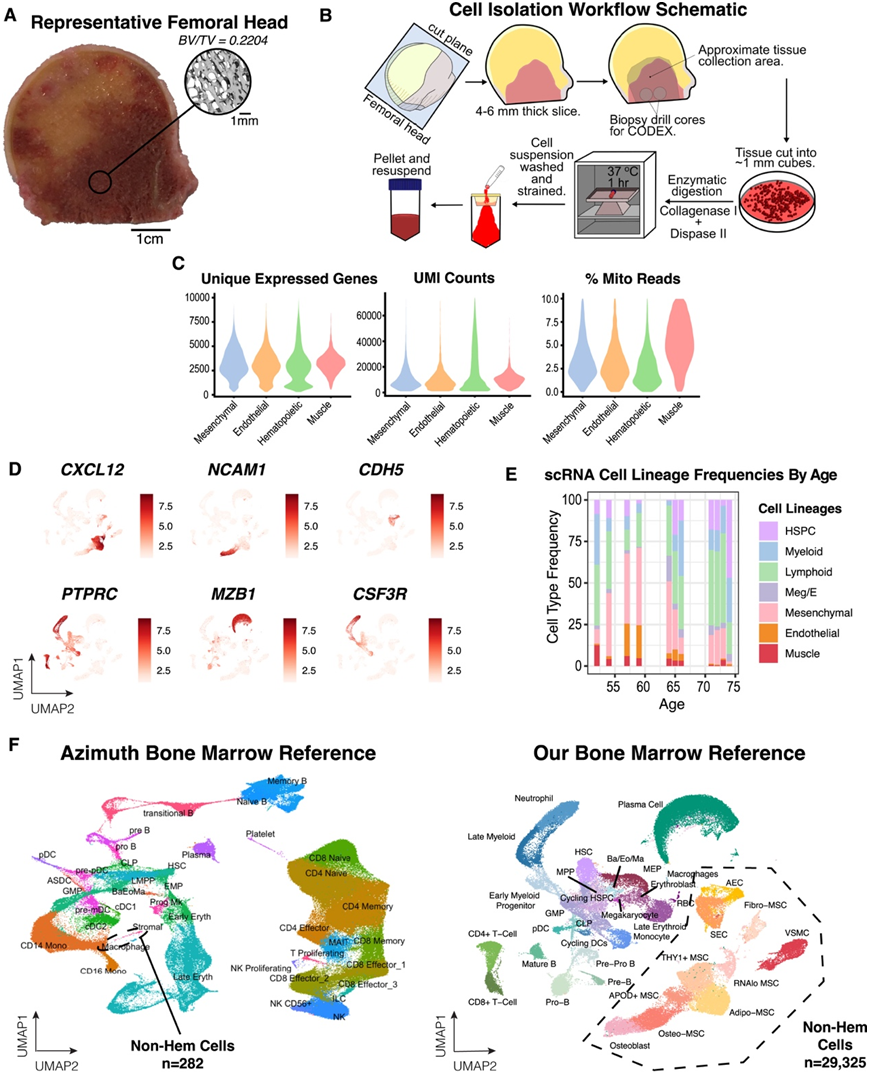
图S1. 人骨髓图谱的特征。
(A)在酶消化之前作为全髋关节置换手术的手术废物接收的股骨头样本的代表性图像,显示许多样本中存在充足的可见红骨髓。(B) 显示股骨头酶消化和细胞分离方案的图表。(C)小提琴图显示独特表达基因的最终图谱分布,测序深度以及过滤掉低质量细胞后映射到线粒体基因组的读数百分比。(D)UMAP可视化。(E) 按捐赠者年龄分层的细胞类型频率。(F) 与本项研究相比,公开骨髓图谱中基质细胞含量的比较。

图1. 人骨髓造血和非造血细胞单细胞转录组图谱。
(A) scRNA-seq 工作流程示意图。(B) UMAP可视化。(C) 条形图显示捕获的每个谱系的细胞计数(左)和每个样本的细胞谱系比例(右)。(D) 热图显示差异表达和关键细胞谱系标记基因表达。
02
非造血细胞亚群分析证明 MSC 和 EC 转录多样性
他们在间充质细胞中发现了显著的异质性,鉴定了骨谱系(NCAM1、SPP1和BGLAP)、脂肪谱系(APOE、LPL、PPARG和CEBPA)和成纤维细胞(PDPN、CSPG4、DCN和DPT)(图2A-B)。成骨细胞分为IBSP/ bglap-高的成骨细胞和IBSP/ bglap-低的成骨间充质干细胞(图2B)。他们发现了具有最高CXCL12水平和高表达脂肪谱系基因(如CEBPA, PPARG, APOE和LPL)的细胞群,将其标记为Adipo-MSC(图2B)。这些细胞类似于报道的小鼠骨髓脂肪生成谱系前体(MALP)和脂肪-Cxcl12丰度网状(Adipo-CAR)细胞。他们还发现了一个强烈表达脂肪细胞基因的THY1+ MSC群体,但也有一个独特的表达谱,包括THY1和LBP,但与Adipo-MSC相比,LPL表达较低,他们称之为THY1+ MSC(图2B)。他们还观察到表达CXCL12、高表达DPT和NT5E以及非常特异性表达PDPN和HAS1的成纤维细胞样细胞,他们将其称为纤维间充质干细胞(图2B)。他们还发现了一个APOD+ GSNhi细胞群,无法将其与先前已知的人类或小鼠细胞类型联系起来,因此他们将其命名为APOD+ MSC,因为该基因高度特异性表达(图2A-B)。Fibro-MSC群体与先前在小鼠中描述的早期间充质祖细胞群体(由CPSG4、CD34和DPT表达定义)以及人类骨骼干细胞群体(由PDPN、CD164和NT5E(CD73)表达定义且缺乏谱系标志物表达)高度一致。纤维间充质干细胞还表达国际细胞治疗学会(ISCT)定义的间充质干细胞标志物,即NT5E (CD73)、THY1 (CD90)和ENG (CD105)(图2C)。需要注意的是,这三个基因在不同的MSC亚群中表达高度可变,在使用它们分离MSC时应考虑这种偏差。特别是,将CD90(THY1)作为分选标志物可能会使分离的MSC群体偏向Fibro-和THY1+ MSC。NGFR(CD271)在各亚群中的表达更为一致,因此可以继续作为一个无偏的MSC标志物(图2C)。
使用 CytoTRACE 伪时间分析,他们确定 Fibro-MSC 是 MSC 亚群中最原始的,这与它们与报道的间充质祖细胞的转录组相似性和 ISCT 定义一致(图2C-D)。为了验证Fibro-和THY1+ MSC更具干细胞/祖细胞特性,他们根据从scRNA-seq数据中获得的标志物,分选了四个MSC亚群(Adipo-MSC、THY1+ MSC、Osteo-MSC/成骨细胞和Fibro-MSC),并进行了成纤维细胞克隆形成单位(CFU-F)实验。分选出的Podoplanin+ Fibro-MSC展示了最高的CFU-F形成能力,其次是THY1+ MSC,而Osteo-MSC和Adipo-MSC表现最差(图2E)。培养的Fibro-和THY1+ MSC在高代次时仍保持增殖,而Adipo-MSC和Osteo-MSC则逐渐减慢(图2F)。Fibro-MSC在适当的分化培养基中培养后,可以分化为成骨细胞、脂肪细胞和软骨细胞。他们的数据还捕捉到了两大类骨髓EC——AEC和SEC(图2G)。AEC表现出较高的CXCL12、ICAM2和PODXL表达,但静脉基因如EPHB4和NR2F2的表达水平比SEC低。相比之下,SEC表达了更多与内皮-造血细胞相互作用相关的基因(例如ACKR1和SELE)(图2H)。
为了进一步强调捕捉非造血细胞异质性的重要性,他们将多个已发表的健康人MSC的scRNA-seq数据集映射到本项研究的图谱上,Adipo-MSC占主导地位(图2I)。本项研究图谱捕捉到的非Adipo-MSC亚群数量比所有映射的成年数据集的总和多24.4倍(图2I)。在其他成年数据集中,基本上没有捕捉到Fibro-MSC。在所有其他数据集中,仅捕捉到76个THY1+ MSC,而在本项研究的数据集中则有4178个(图2I)。
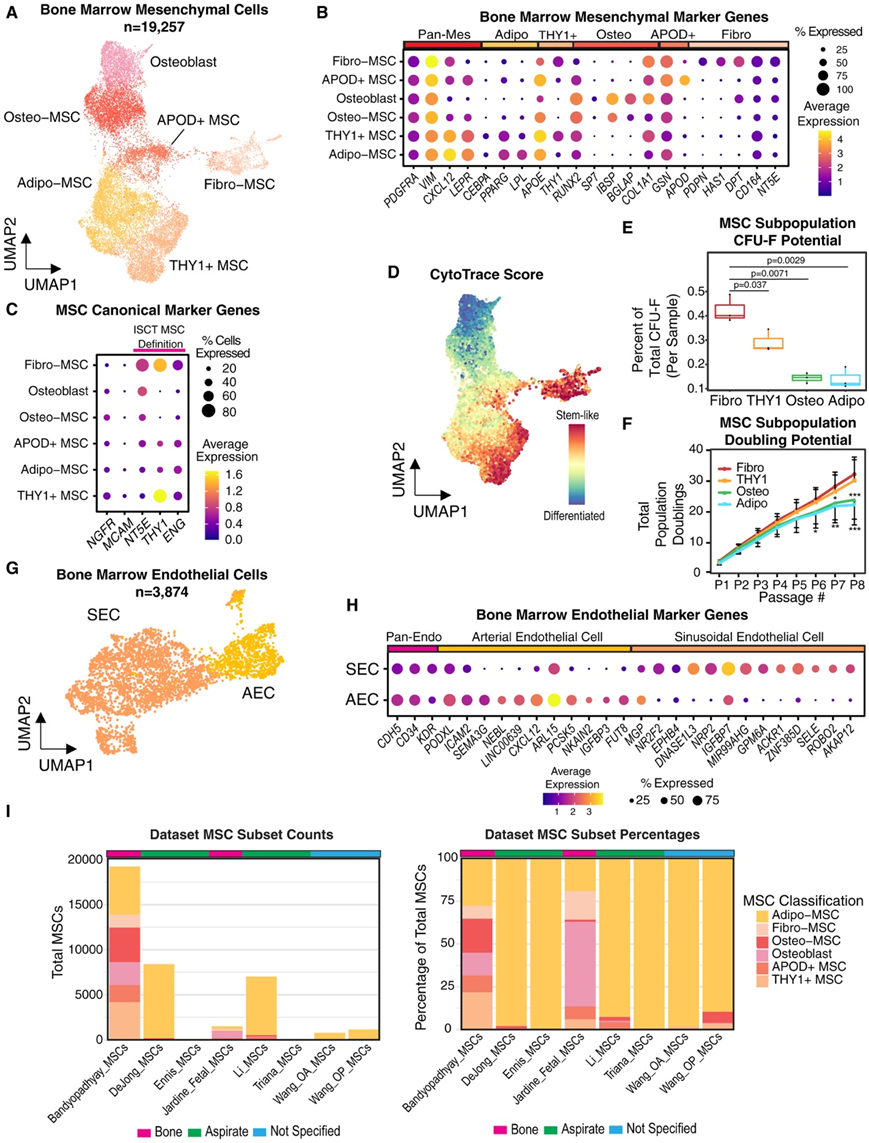
图2. 定义人骨髓的非造血细胞组成。
(A)UMAP可视化。(B) 点图显示 MSC 子集中关键间充质标记基因的标准化表达。(C) 点图显示人类 MSC 的文献来源标记基因的标准化表达。(D) CytoTRACE 分析投射到 MSC UMAP 上,显示预测的分化评分。(E) 箱线图显示分选的 MSC 亚型的相对成纤维细胞克隆形成潜力。(F) 线图显示细胞培养八次传代过程中的群体倍增。(G) UMAP 显示来自 12 个个体的 3874 个内皮细胞。(H) 点图显示了通过调整内皮子集之间的p值选定的泛内皮基因和差异表达基因的标准化表达。(I) 本研究与已发表的研究之间人骨髓 MSC 子集的参考图谱。
03
MSC、EC 和骨系细胞合作产生多种造血支持因子
接下来他们探索 MSC 异质性是否会转化为造血支持因子的转录水平差异。从造血因子开始,造血因子已在非造血细胞亚群中得到广泛研究(图3A),他们发现不同的MSC类型专门产生特定的支持因子;例如,CXCL12在 Adipo- 和 THY1+ MSC 中表达最高,而IGF1主要由 Fibro-MSC 表达。
接下来,他们使用CellChat系统地研究了骨髓中基于同源配体和受体共表达的细胞通讯,如CXCL12和SELE在MSC和SEC与不同HSPC亚群之间的相互作用(图3B)。为了验证HSPC和MSC亚群之间预测的相互作用,他们进行了新鲜分选的MSC亚群和新鲜分离的CD34+细胞的共培养实验。发现分选出的Adipo-、Osteo-和THY1+ MSC显著保护了原始的Lin− CD34+ CD38−细胞,Fibro-MSC也表现出强烈的保护趋势,相比之下,在单独培养中这些细胞几乎无法保存(图3C)。他们还发现,总体而言,间充质谱系细胞具有最多的输出信号贡献(即它们表达配体的显著相互作用)(图3D)。
接下来,他们研究了非造血细胞与造血细胞的支持因子表达和预测的通讯,重点关注HSPC的维持、髓系造血和淋巴系造血。根据CellChat数据库中描述的共有路径注释对支持因子进行了分组(例如,ANGPT家族包含多个血管生成素基因)。据报道,包括成骨细胞和血管周围的MSC在内的几种细胞类型对HSPC的维持非常重要。成骨细胞和MSC可能都支持人类HSPC,但大多数经典的支持因子是由MSC亚群,特别是Adipo-和THY1+ MSC产生的。例如,Adipo-和THY1+ MSC产生了最高水平的经典HSPC微环境因子,如CXCL12(CXCL家族)和KITLG(KIT家族),这些因子由造血细胞广泛接收,但对HSPC没有强特异性(图3A、3E-F)。成骨谱系细胞比Adipo-或THY1+ MSC表达更多的TGFB1和CDH2(图3A),这些因子在小鼠中已证明支持造血干细胞(HSC)的静息状态。HSC还从SEC提供的E-选择素中预测出显著的输入信号(图3A-F)。
他们接下来关注已知的对骨髓形成重要的因素。CSF1在多个MSC亚群中表达,在Adipo-MSC中表达量最高(图3A)。虽然大多数造血支持因子是由间充质细胞提供的,但他们也注意到,通常不是关键生态位细胞的CD4+ T细胞是骨髓中FLT3LG的主要产生者(图3E)。预测造血干细胞、粒细胞单核细胞祖细胞(gmp)和普通淋巴样祖细胞(CLP)接受FLT3LG,已知FLT3LG对骨髓生成和淋巴生成都至关重要(图3F)。
然后,他们通过研究IL-7和Notch配体等因素,分析了微环境对淋巴生成的贡献。IL-7主要由Adipo-和THY1+ MSC产生(图3A)。总的来说, EC和血管平滑肌细胞(VSMC)确实是人类骨髓中DLL1、DLL4、JAG1和JAG2的主要贡献者,其中动脉(AEC和VSMC)表达最多的JAG1(图3A)。他们还注意到,虽然来自间充质细胞的Notch信号(图3E)较弱,但也预测到这种信号的输出,主要基于JAG1表达。值得注意的是,VSMC还通过NOTCH3接收了显著的Notch信号,可能是从AEC接收的(图3)。有趣的是,所有这些非造血细胞类型以及HSPC在一定程度上都预测接收Notch信号,提出了微环境成分之间可能存在的交互和协调调节的可能性。值得注意的是,成骨细胞并未表现出高水平的淋巴相关因子的表达(图3A)。
使用非负矩阵分解(NMF),他们在骨髓中识别出更高级别的通讯模块。不同的非造血亚群贡献于不同的模块,并根据所涉及的细胞类型对这些模块进行了注释(图3G)。内骨膜模块以骨特异性基因家族如NGF、ncWNT和骨桥蛋白为特征,与表达KIT、FGF、THY1等家族的MSC模块显著不同。CXCL12hi的MSC亚群如Adipo-和THY1+ MSC主要向MSC模块贡献独特的信号,而Osteo-或Fibro-MSC则参与了内骨膜和MSC两个模块(图3G)。此外,与通路级CellChat分析一致,VSMC和较少程度的AEC对内骨膜和血管信号模块都有贡献。
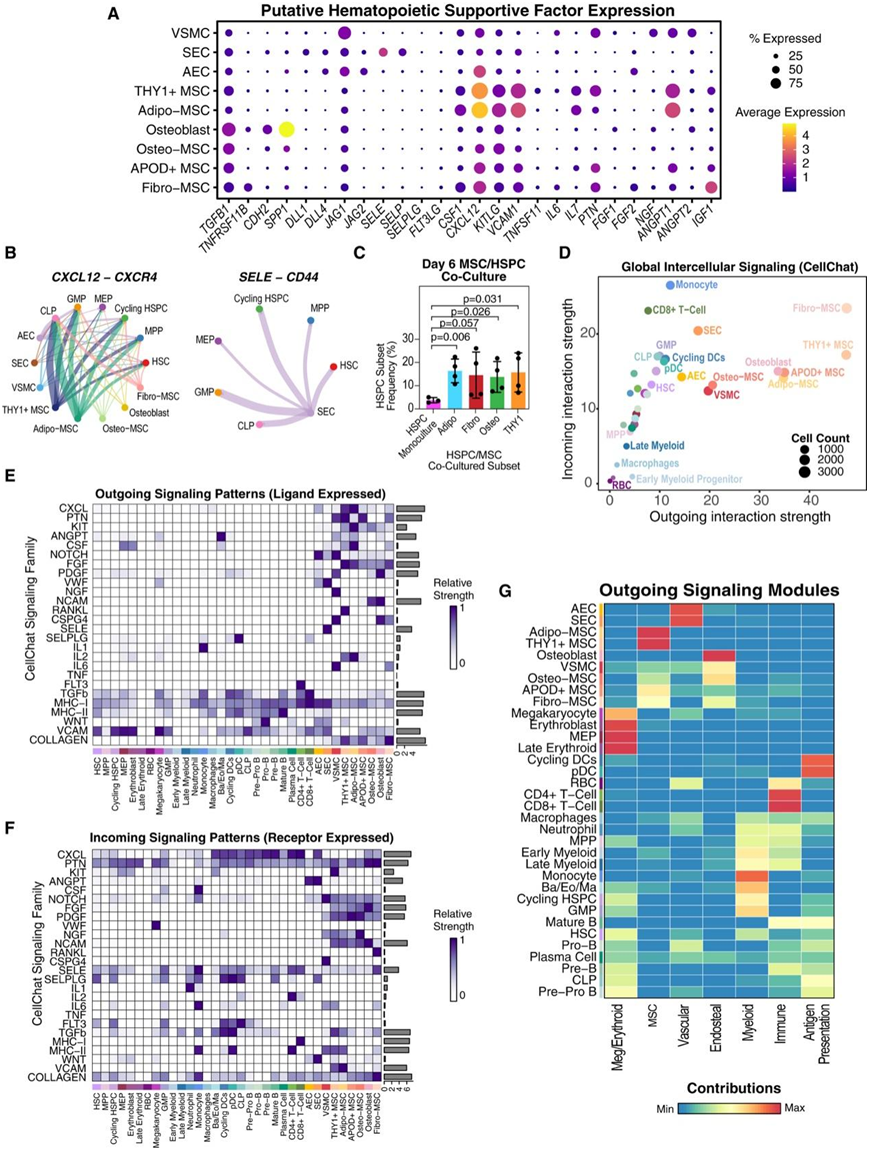
图3. 细胞间通讯分析揭示了造血细胞和非造血细胞类型之间不同的信号传导模式。
(A) 点图显示间充质和内皮细胞群中手动策划的造血支持因子的标准化表达。(B) 和弦图显示 CellChat 预测的源间充质/内皮细胞类型与靶标 HSPC 细胞类型之间的显著相互作用。(C) 新鲜分离的 CD34+ HSPC 与分选和培养的 MSC 子集的共培养。(D) CellChat 网络分析。(E-F)对每个带注释的信号家族的传出(配体富集)和传入(受体富集)显著信号传导预测进行评分。(G) 非负矩阵分解 (NMF),用于识别传出信号模块以及每种细胞类型对模式的作用。
04
CODEX 多重成像揭示了人体骨髓生态位的原位解剖结构
接下来,他们试图定义在 scRNA-seq 图集中定义的骨髓细胞类型的单细胞解剖结构。他们对 12 个样本采用了全玻片 CODEX 多重成像,其中 8 个包含在本项研究的转录组图谱中(图 4 A)。在scRNA-seq数据的指导下,他们设计并验证了一个包含54个标记的面板(53个抗体+DAPI)(图4A-B)。例如,除了经典的HSPC标记物如CD34和CD38外,还包括了从scRNA-seq数据中衍生的标记物如SPINK2,该标记物在原始HSC中表达最高。这提高了识别原始HSPC的分辨率,因为CODEX检测的动态范围低于流式细胞分析,其中CD34和CD38是金标准。他们获取了全片CODEX图像,并随后使用Mesmer对其进行分割以识别单个细胞。然后通过无监督聚类对细胞进行注释,并在少数情况下通过手动分选进行细化。通过计算机注释了803132个细胞(占分割对象的91.6%)(图4C),涵盖了12个样本中的32种细胞类型,包括罕见的细胞,如免疫表型的HSC(Lin− CD34+ CD38− CD45RA− CD90+)和雪旺细胞(PLP1+ CD271+)(图4C)。
他们将鉴定的非造血细胞分为AEC、SEC、VSMC、内皮细胞、Adipo-MSC和THY1+ MSC。值得注意的是,尽管存在一些组织加工产物,如骨组织的频繁丢失,仍然能够识别出具有非造血表达谱的小梁旁细胞,将其称为“内皮细胞”。骨间充质干细胞(Osteo-MSC)和纤维间充质干细胞(Fiber-msc)最初都没有通过计算检测到,这表明这些细胞与骨的联系更紧密,因此在组织加工过程中丢失。经过更仔细的分析,确实检测到这些细胞的罕见实例(图4D),在造血骨界面上检测到Osteo-MSC和成骨细胞,并且在骨区域中部检测到大量Podoplanin+ Fibro- msc(图4D-E)。此外,CODEX细胞类型的蛋白质表达谱与scRNA-seq数据很好地匹配,匹配的细胞类型得分非常高。最后的细胞类型注释用于在“细胞表型图(cpm)”中可视化细胞拓扑结构(图4F)。总的来说,这个带注释的参考文献作为造血细胞微环境背景下的细胞拓扑的综合地图,并证明了这两个地图集的互补性,以了解人类骨髓的细胞解剖。
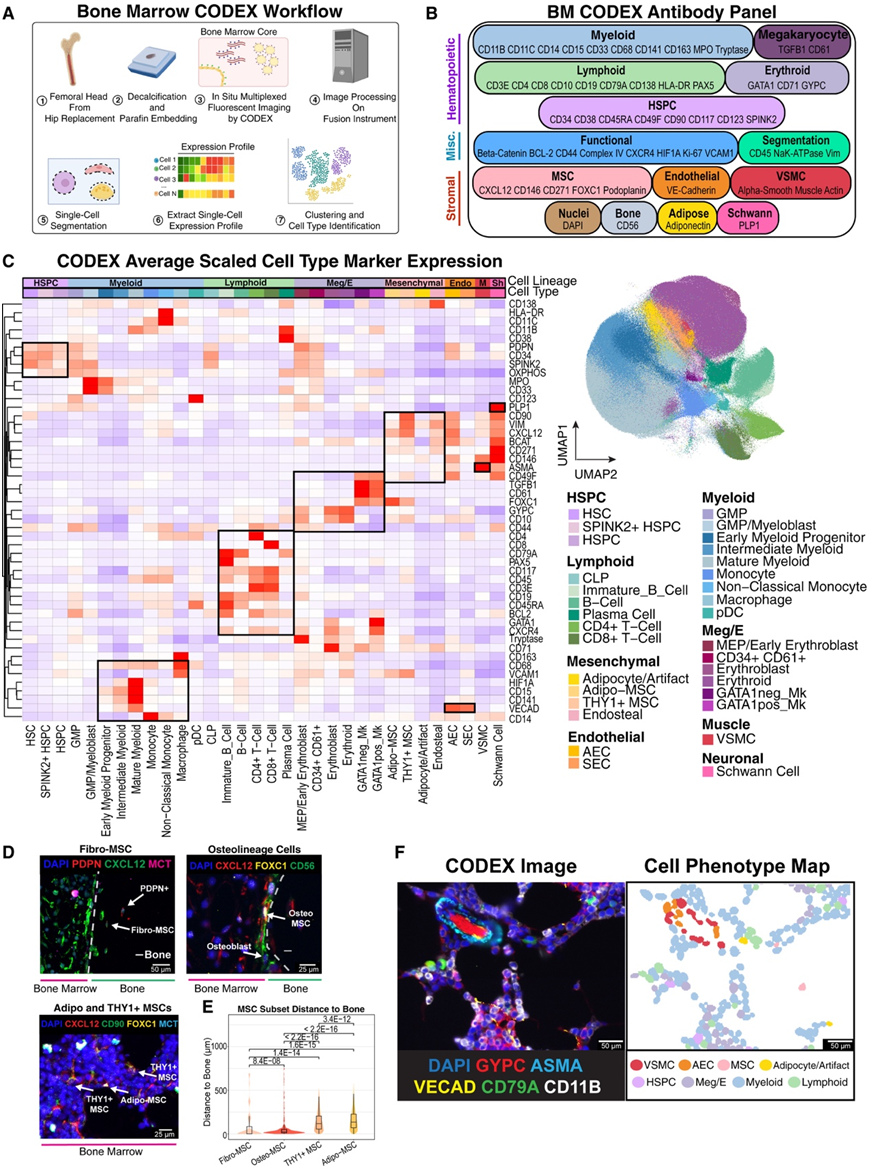
图4. 54 重 CODEX 成像揭示人骨髓的空间细胞拓扑。
(A) 示意图描述了导致细胞类型识别的 CODEX 实验和计算工作流程。(B) 5-抗体CODEX按靶细胞群划分。(C) 热图显示按蛋白质标记缩放并按蛋白质表达层次聚类的每种细胞类型的平均中心对数比 (CLR) 标准化表达(左)。方框突出显示协调的标记表达。 UMAP 显示了 CODEX 图集中来自 12 个个体的 803131 个单细胞,按细胞类型着色(右)。(D) CODEX 图像分析。(E)小提琴图显示不同 MSC 亚型与手动标记细胞的最近距离带注释的骨骼轮廓。(F) CODEX 图像(左)与细胞表型图(右)配对。
05
EMP 和 GMP 定位于相对高氧的动脉内膜生态位
虽然已经提出了多个生态位来组织造血,但试图利用一种公正的、统计严谨的方法来回答造血细胞和间充质细胞如何组织成细胞邻域(CN)的问题。事实上,即使使用CODEX图像的交互式可视化,他们也注意到细胞组织的离散模式,例如MPOhi早期髓系祖细胞(EMP)的内皮周围定位,以及由巨噬细胞和红细胞前体组成的红细胞岛。因此,他们进行了如前所述的无监督邻域分析并验证了根据细胞类型富集手动注释的 15 个 CN(图 5 A)。发现了预期的邻域,例如与成红细胞岛一致的“红系”邻域(CN13和CN15),以及新的邻域,例如小动脉周围淋巴细胞的一个邻域(图5A)。他们还发现了具有所有三种造血谱系(红系/髓系/淋巴系)的多个混合谱系群体(CN7和CN8)。HSPC包括更原始的SPINK2+ HSPC,在混合谱系社区(CN7和CN8)中发现最多,而SPINK2+ HSPC在髓系/淋巴(CN5)和早期髓系/小动脉(CN4)社区中发现较少。淋巴样祖细胞如CLP和未成熟B细胞主要存在于混合谱系(CN7和CN8)、浆细胞/小动脉(CN3)和小动脉周围淋巴样细胞(CN2)中。两个富集GMP/成髓细胞和早期髓系祖细胞(EMP)的邻域(CN4和CN6)强烈定位于髓内表面和小动脉周围(图5A-C)。这与CellChat识别的内皮信号模块一致,该模块由aec和VSMC贡献(图3G)。因此,根据aec和内皮细胞的富集程度,分别将它们命名为早期髓系/小动脉(CN4)和早期髓系/内皮(CN6)。值得注意的是,与早期髓系、血管/内皮/间充质干细胞或浆细胞/小动脉相邻区不同,动脉周围淋巴邻近区内皮细胞不富集(图5A)。最后,富含血管/骨内膜/间充质干细胞的区域高度富集雪旺细胞,表明该区域受到神经支配。所有小动脉和混合谱系邻域(CN1-4 和 CN7-8)均富含 Adipo- 和 THY1+ MSC(图 5 A)。
他们试图利用CODEX和scRNA-seq图谱进一步研究该内皮生态位内的相互作用。首先,发现小动脉细胞出现在骨小梁附近的频率高于随机预期(图5B),这表明内皮生态位实际上可能与小动脉生态位结合。其次,他们发现HIF1A水平在emp和gmp中较低,表明这些细胞没有经历缺氧(图5C),这与它们在供氧血管附近的定位有关。相比之下,更成熟的髓细胞具有高水平的HIF1A。此外,非小动脉区域的emp具有更高水平的HIF1A,这表明缺氧的空间模式不是由于固有的细胞类型差异(图5D)。同样,他们发现emp在scRNA-seq图谱中具有最低的缺氧特征评分(图5E)。
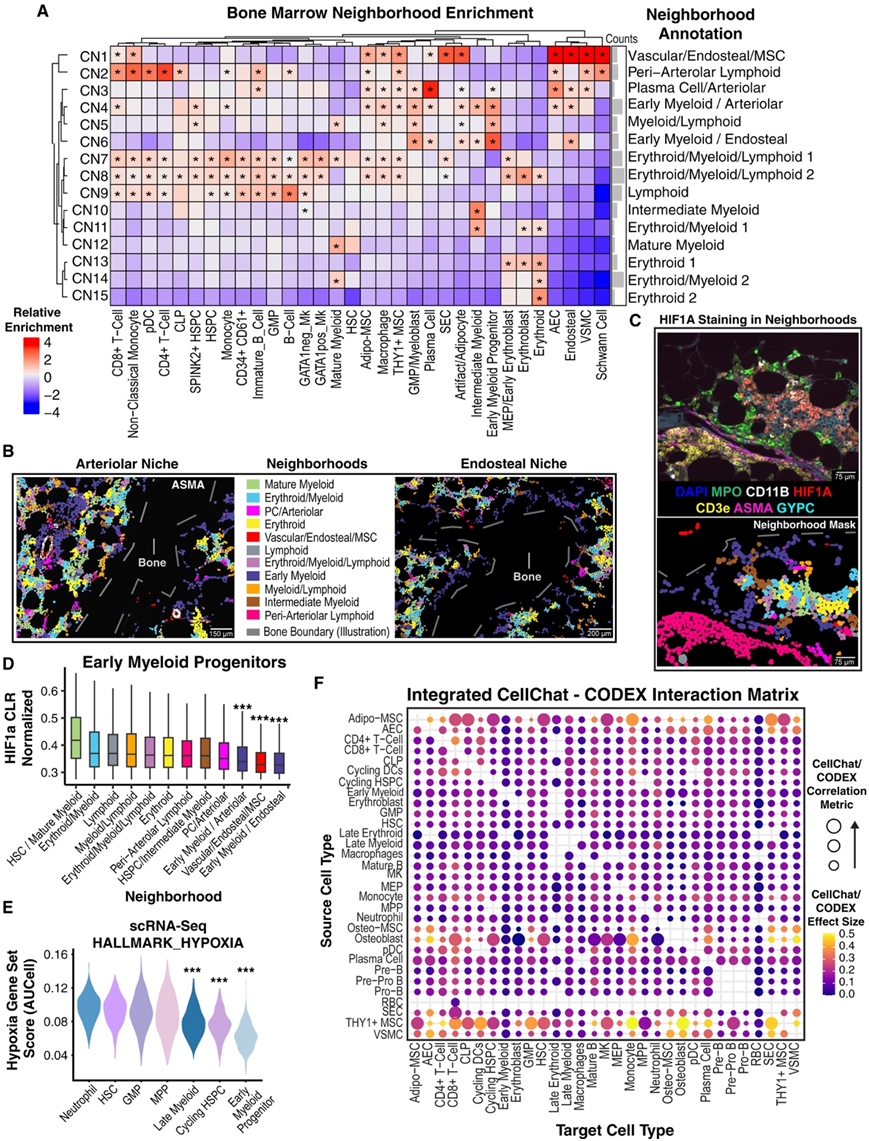
图5. 邻域分析揭示了 GMP/早期骨髓祖细胞的富氧小动脉/骨内膜生态位。
(A) 热图显示邻域中存在的细胞类型的富集。 (B)在ASMA荧光信号旁绘制由邻区成员着色的细胞表型掩模。(C) 跨邻域的 HIF1A 染色模式。(D) 箱线图显示早期骨髓祖细胞中按邻域成员划分的标准化 HIF1A 表达水平。(E) 小提琴图显示使用 AUCell 和scRNA-seq 数据计算的缺氧特征分数。(F) 气泡图显示两种细胞类型的 CellChat 交互预测强度和最大 CODEX 空间邻域共定位强度之间的相关性和 CODEX-CellChat 效应大小 (CCES)。
06
基于成像的显微解剖结构分析揭示了 HSPC 的脂肪细胞周围定位
接下来,他们研究了六种关键的微环境结构,这些结构由于其复杂的形状和/或大小,通常无法通过传统的细胞分割方法很好地捕捉到,包括脂肪细胞、小动脉、骨、巨噬细胞及其突起、窦状隙和CXCL12+基质。在所有测量的结构中,小动脉最靠近骨(图6A-B),这支持了邻域分析的发现,即内骨膜微环境富含小动脉(图6B)。总体上,他们发现定义为与骨无显著统计邻近关系的中央微环境由窦状隙、脂肪细胞、基质和巨噬细胞组成,尽管脂肪细胞和窦状隙比基质或巨噬细胞更远离骨(图6A-B)。此外,结构分析证实了先前提到的红细胞微环境对应于红细胞岛,因为这种微环境非常靠近巨噬细胞,而由于其不规则的形状和大小,巨噬细胞在邻域分析中未捕捉到。早期髓系细胞邻域靠近小动脉和骨,进一步证明了小动脉-内骨膜EMP微环境的存在(图6C)。这些结果也验证了细胞类型注释,因为细胞类型与预期的结构共定位,如脂肪-间充质干细胞-基质、巨噬细胞-巨噬细胞、骨内皮和sec -窦状体(图6D)。有趣的是,所有的MSC都靠近动脉内皮壁位,但Adipo-MSC靠近窦状窦,而THY1+ MSC则没有(图6D)。
他们还发现了在基于细胞分割的分析中不明显的空间模式。发现Lin - SPINK2+和SPINK2 - CD34+ HSPC比预期的更接近脂肪细胞(图6D-E)。CODEX图像显示,两种类型的HSPC都与脂肪细胞频繁接触(图6F)。HSPC,而不是更原始的SPINK2+ HSPC,也与巨噬细胞接近(图6E)。有趣的是,GMP/成髓细胞和EMP也接近脂肪细胞,尽管这些细胞明显更接近骨,而HSPC则不是。相比之下,成熟的髓细胞最靠近窦状隙,而不是小动脉或骨(图6D)。总的来说,这些数据表明骨髓发育的空间限制,其中最早的骨髓祖细胞在HSPC的脂肪细胞生态位中,迁移到相对高氧的动脉-内皮早期骨髓生态位,最后迁移到窦状动脉附近并成熟,它们可以利用窦状动脉从骨髓进入血液(图6G)。
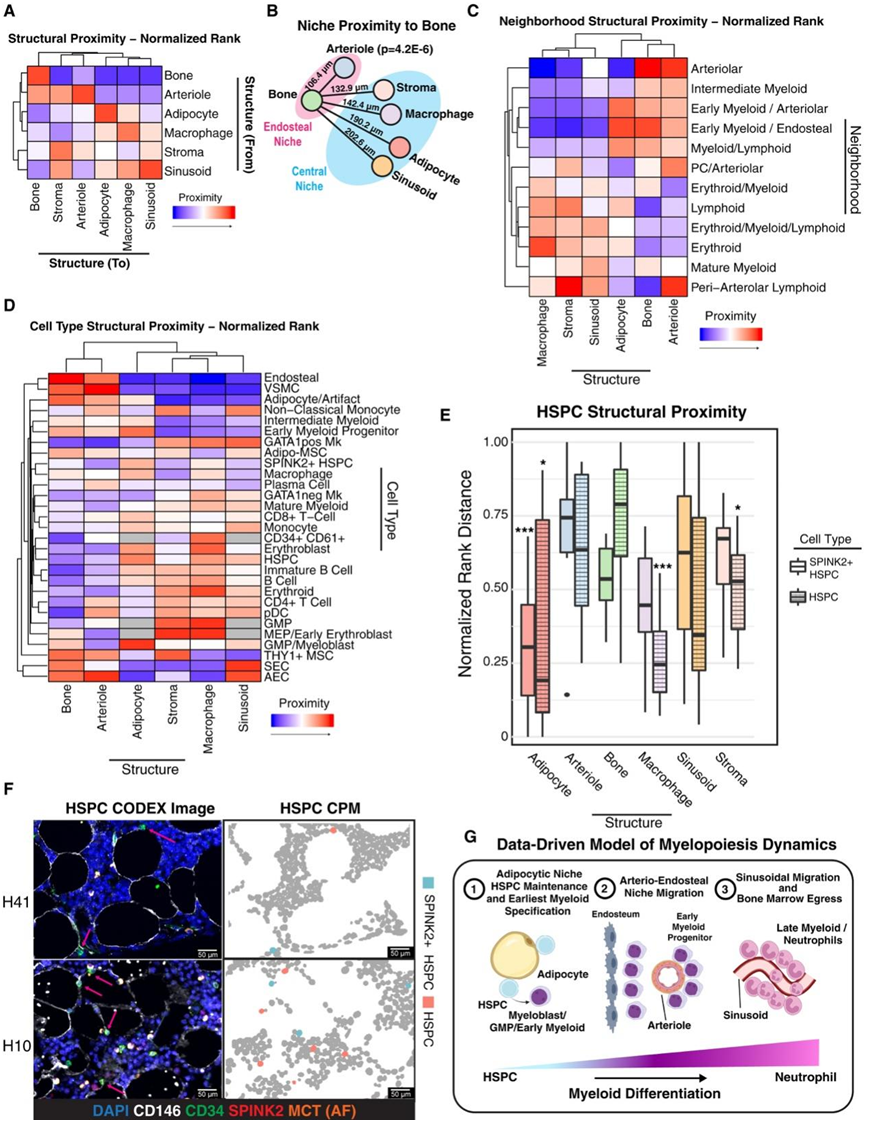
图6. 骨髓微环境的综合结构分析揭示了谱系非特异性HSPC的脂肪细胞定位。
(A)热图显示了每个微环境结构与每个结构的标准化等级接近度。(B) 正常骨髓中结构邻近性的数据驱动图示。(C-D) 热图显示每个邻域和细胞类型与每种类型的微环境结构的标准化排序接近度。(E) 箱线图显示 SPINK2+ HSPC(实线框)和 HSPC(虚线框)与每个微环境结构的标准化排名接近度。(F) 两个样本的代表性 CODEX 图像和相关细胞表型图。(G) 显示了人类骨髓生成空间动力学的模型。
07
使用健康骨髓 CODEX 图谱进行无监督参考绘图揭示了 AML 基质扩张和富含 AML-MSC 的新区域
了解骨髓微环境的空间组织对健康造血以及疾病状态有重大影响。例如,急性髓性白血病(AML)已描述为与内骨膜微环境相互作用以获得化疗耐药性。他们应用骨髓CODEX面板,以健康图谱为参考,探索肿瘤演变和微环境变化。对来自髂嵴活检的三例诊断(Dx)和两例接受Ven/HMA(维奈托克斯加去甲基化剂)治疗的配对治疗后(PostTx)AML患者样本进行了分析。作为对照,他们还纳入了3例38- 62岁的髂骨淋巴瘤分期阴性骨髓活检(NSM)患者。他们选择了检测到NPM1突变(NPM1c W288 * fs)的AML患者,因为可以获得突变特异性抗体,因此他们可以识别假定的白血病原细胞。他们采用互反主成分分析(RPCA)参考图谱,将AML和NSM样本中的细胞与健康图谱中最接近的对应细胞进行分类。正如预期的那样,他们发现与NSM相比,Dx AML中除成熟髓细胞外的髓细胞比例显著增加(图7B)。
将这些细胞标签投影到分割掩模上显示,与 NSM 样本相比,Dx 和 PostTx AML 样本中的结构发生了明显的变化,例如白血病环境中脂肪细胞几乎完全丧失,以及在存在残留原始细胞的情况下,PostTx 后 30 天造血恢复不完全(图7C)。对 AML 细胞景观的更详细观察还揭示了显著的基质扩张,与 NSM 样本相比,AML 中 Adipo- 和 THY1+ MSC 的相对频率高出 2 至 3 倍(图 7 D)。他们在 AML3 的 PostTx 样本和单个NPM1突变 PostTx 细胞中鉴定了白血病群体,包括 GATA1+ NPM1c+ 双阳性群体(图 7 E)。在两个 PostTx 样本中观察到NPM1突变母细胞中 GATA1 表达增加了约 2 倍,表明这些残留细胞表现出谱系可塑性(图 7 F)。
接下来,他们分别对 AML Dx、PostTx和 NSM样本进行邻域分析,并在每个样本中识别出 15 个 CN(图 7 G)。在Dx AML标本中,发现了四个急变富集的邻域Dx-CN3/CN5/CN9/CN12(图7G),其中两个是MSC群体富集的。值得注意的是,THY1+ MSC仅在这些急变富集的邻域中显著富集(图7G)。他们还观察到,恢复后的 Tx GMP/EMP 邻域不包含 AEC/内膜细胞 (PostTx-CN8),这可能反映了稳态和紧急骨髓生成之间的差异(图7G)。
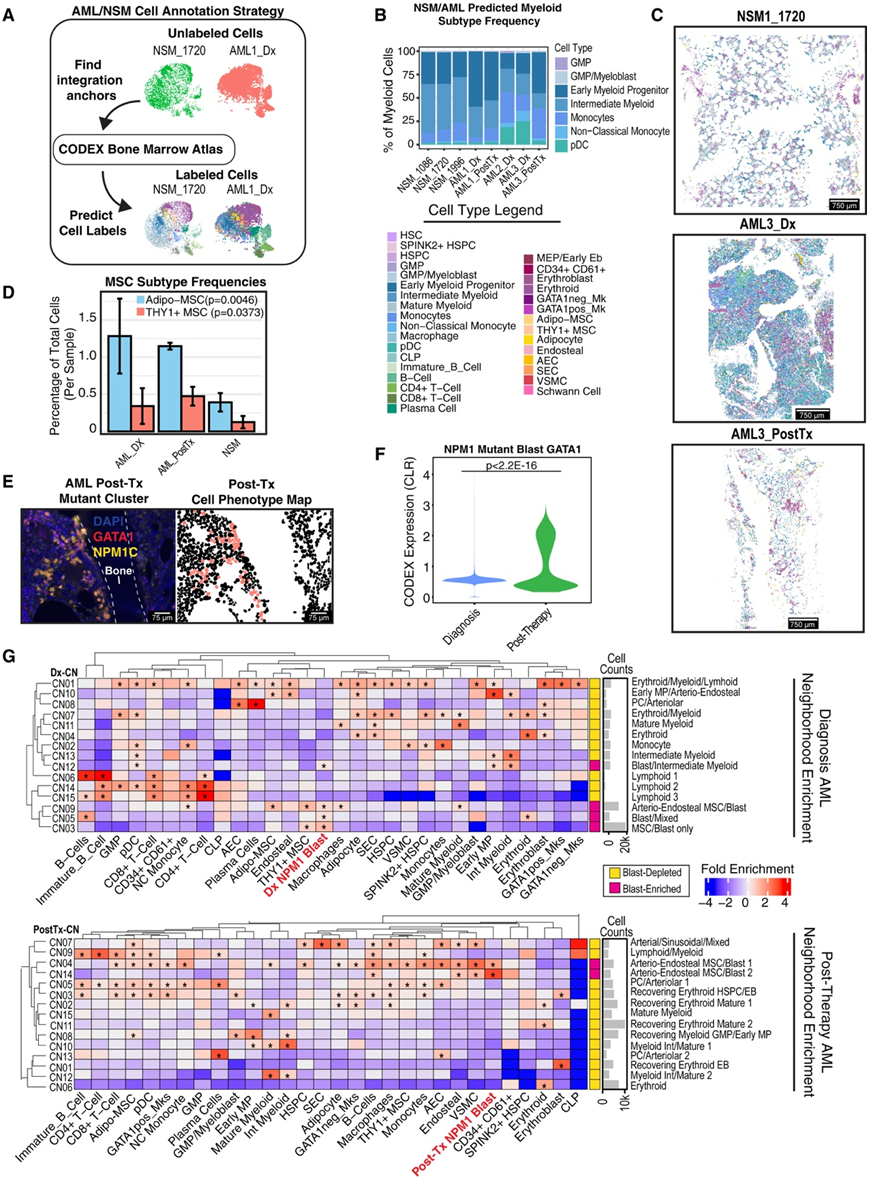
图7. 无监督的 AML 单细胞图谱揭示了基质扩张和富含 MSC 的 AML 特异性邻域。
(A) 示意图显示了无监督标签转移计算方法。(B)条形图显示每个样本中的骨髓细胞类型频率。(C) 显示代表性样本图像的细胞表型图 (CPM)。(D) 条形图显示 AML 与 NSM 中 MSC 的每个样本频率。(E) CODEX 图像显示罕见残留NPM1突变母细胞的聚集。(F) 小提琴图,显示来自诊断和治疗后样本的 NPM1 突变母细胞中的 GATA1 蛋白水平。(G) 显示邻域细胞富集的热图。
+ + + + + + + + + + +
结 论
本项研究使用scRNA-seq对 29325 个非造血细胞进行了分析,发现了九种转录上不同的亚型。同时分析了 53417 个造血细胞并预测了它们与非造血细胞亚群的相互作用,采用CODEX对超过 120 万个细胞进行空间分析。整合了 scRNA-seq 和 CODEX 数据,将预测的细胞信号传导与空间邻近性联系起来,揭示了早期骨髓生成的高氧动脉内膜邻域,以及早期HSPC的脂肪细胞定位。使用 CODEX 图集注释新图像,并发现AML患者样本中的MSC 扩张和白血病母细胞和 MSC 共同富集的空间邻域。这个空间分辨的人类骨髓多组学图谱为研究驱动造血的细胞相互作用提供了参考。
+ + + + +



