

 English
English文献解读|Cell(45.5):空间转录组景观揭示免疫球蛋白相关衰老是衰老的标志
✦ +
+
论文ID
原名:Spatial transcriptomic landscape unveils immunoglobin-associated senescence as a hallmark of aging
译名:空间转录组景观揭示免疫球蛋白相关衰老是衰老的标志
期刊:Cell
影响因子:45.5
发表时间:2024.11.04
DOI号:10.1016/j.cell.2024.10.019
背 景
生物体组织错综复杂,数十亿个细胞作为功能单位跨器官、系统和整个身体进行协调。在衰老组织中,分子和细胞变化会破坏空间组织,导致功能衰退。研究这些时空动态可以揭示新的衰老机制和潜在的治疗靶点。然而,单胞转录组学可以在年轻和老年生物体中生成详细的基因表达谱,将细胞与其原生组织环境分离。空间分辨转录组学将细胞位置与测序数据联系起来,提供了对组织内基因表达的深入了解。因此,要剖析组织水平的衰老机制,必须探索衰老如何影响完整组织细胞微环境内的细胞和分子动力学。
实验设计
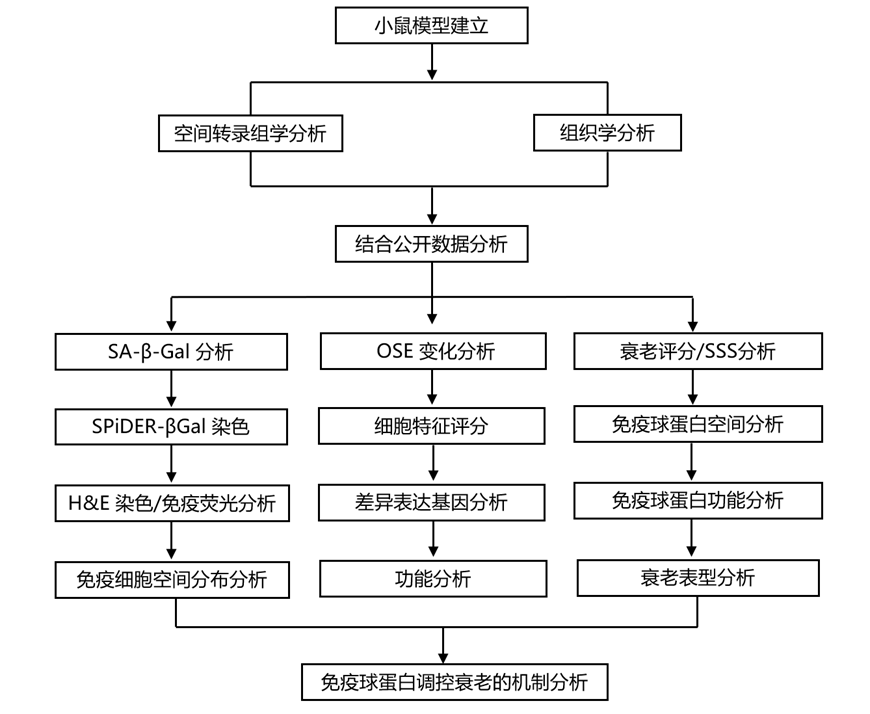
结 果
01

跨多种组织细胞类型的空间分子制图
为了探索各个器官衰老过程中细胞和分子的空间变化,研究团队检查了年轻(2 月龄)和老年(25 月龄)阶段的 C57BL/6J 雄性小鼠的九种关键组织——海马、脊髓、心脏、肺、肝脏、小肠、脾脏、淋巴结和睾丸(图 1 A)。他们首先对年轻和老年组织样本进行了全面的表型比较(图 1 B,图 S1A-J)。在 SA-β-Gal 和 SPiDER-βGal 染色分析中,他们观察到老年雄性小鼠的阳性染色增加,呈现为组织特异性的拓扑模式(图 1 B)。同时,多种组织中 CD45 阳性免疫细胞的数量增加,这是与年龄相关的慢性炎症的特征(图 S1A-J)。因此,他们在老年肝脏中检测到扩大的炎症区域,在海马和脊髓中检测到小胶质细胞(中枢神经系统的主要免疫细胞)数量增加,表明存在神经炎症(图S1A-J)。此外,老年小鼠的心脏和肠道中的纤维化水平均升高(图 S1A-J)。再者,在多种老年组织中观察到内皮细胞(CD31 +)减少,同时在老年海马的齿状回 (DG) 中新生神经元 (DCX+) 减少,这表明微血管密度和神经发生受损(图 S1A-J)。与年轻小鼠相比,他们还发现老年小鼠的器官存在结构变化,例如心脏中心肌细胞的横截面积增加,肺部中肺泡面积扩大(图 S1)。
为了获得更精细的组织分辨率,他们使用立体测序技术对年轻和老年雄性小鼠的 9 种组织进行了空间转录组学分析(stRNA-seq)(图1 C)。他们从 103 个组织样本中获得了 1535191 个高分辨率点,平均每个点有 1450 个基因,并且不同组织类型之间存在差异(图 S2A-C)。用标记基因注释可鉴定出 71 种不同的点类型,与各种组织和器官中已知的细胞成分相对应(图 1 C-D,图S2E)。
他们分析了维持组织稳态和衰老的关键干细胞的空间属性,包括 DG 中的神经祖细胞和小肠隐窝中的肠干细胞(图 1 C,图 S2 E)。这些分析表征了核心功能细胞和基质细胞(如神经胶质细胞、成纤维细胞、平滑肌细胞、内皮细胞和 Leydig 细胞)在组织和器官间质和基质区域内的分布(图 1 C,图S2 E)。此外,他们还绘制了重要免疫细胞的空间分布图,特别是在脾脏和淋巴结等免疫中心,包括巨噬细胞、粒细胞、B 细胞、浆细胞和 T 细胞(图 1 C,图S2 E)。
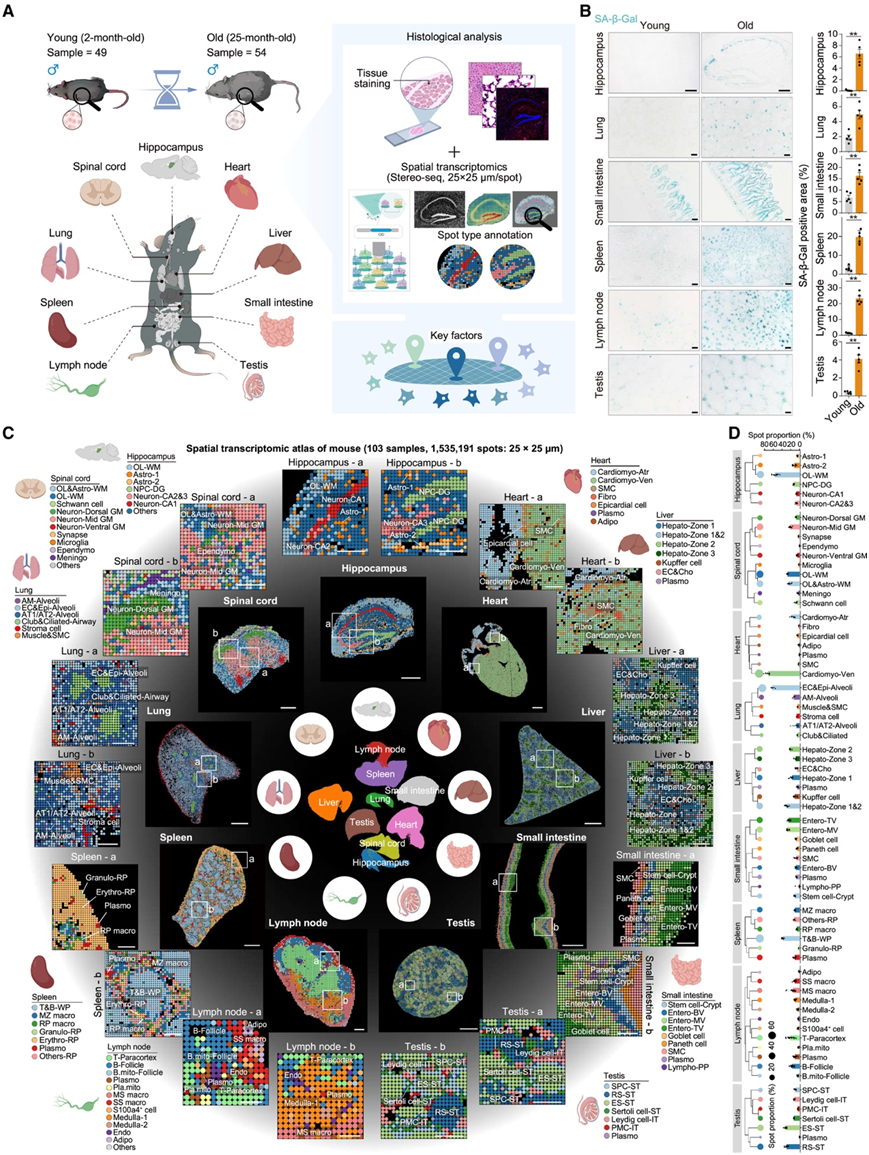
图1. 构建年轻和老年雄性小鼠跨多个组织的空间转录组图谱。
(A) 年轻小鼠和老年小鼠组织采样、组织学分析和空间转录组学测序示意图。(B) 年轻小鼠(2 月龄)和老年小鼠(25 月龄)的多种组织进行 SA-β-Gal 分析。(C) 年轻小鼠组织的空间转录组图谱。(D) 年轻小鼠样本中每种组织中斑点类型的比例。
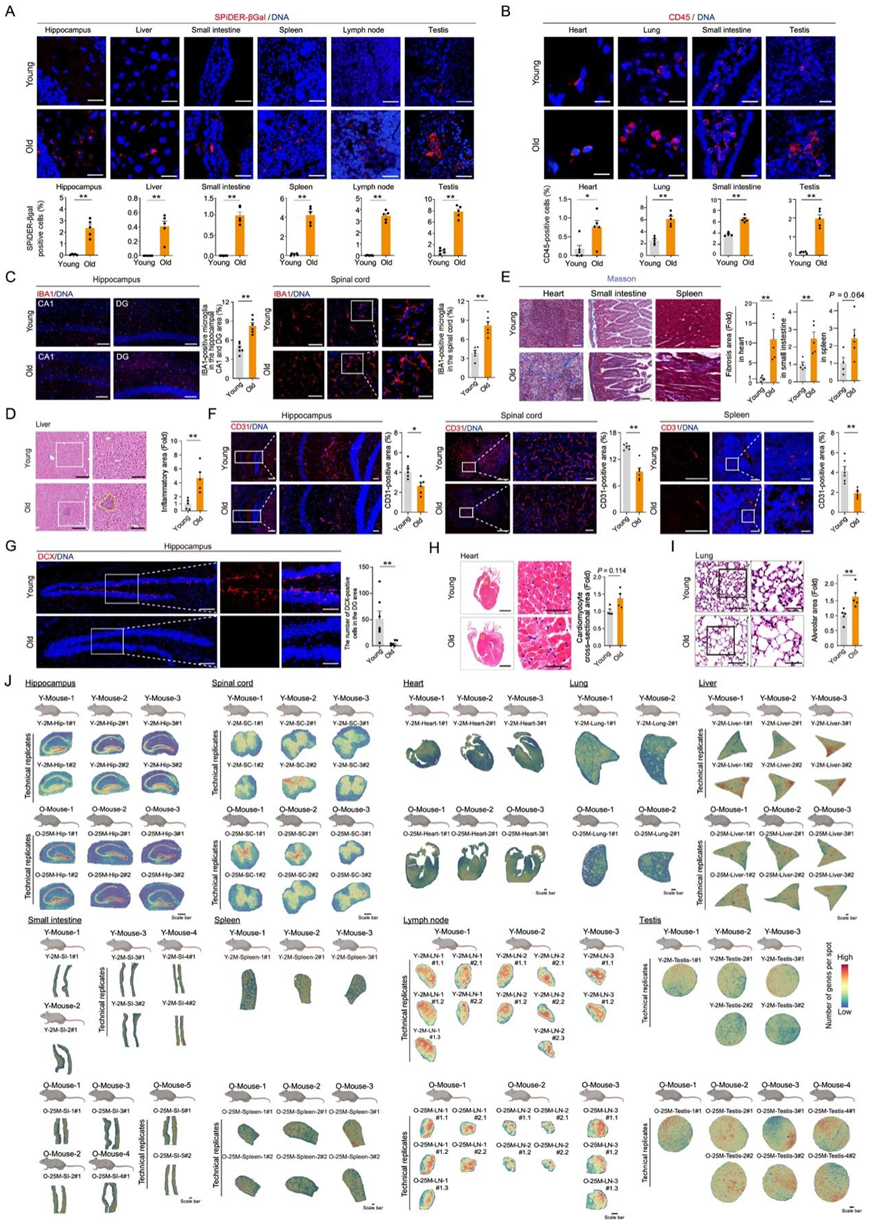
图S1. 对雄性小鼠组织进行形态学和组织学分析,以确定衰老表型。
(A) 多种组织(包括海马、肝脏、小肠、脾脏、淋巴结和睾丸)的 SPiDER-βGal 染色。(B)雄性小鼠的心脏、肺、小肠和睾丸中的CD45的免疫荧光。(C) 年轻和老年雄性小鼠海马和脊髓中小胶质细胞标志物 IBA1 的免疫荧光。(D) 年轻和老年雄性小鼠肝组织的 H&E 染色。(E) 年轻和老年雄性小鼠的脾脏、小肠和心脏的 Masson 三色染色。(F) 年轻和年老雄性小鼠的海马、脊髓和脾脏中的 CD31 免疫荧光。(G) 年轻和老年雄性小鼠海马中 DCX 的免疫荧光。(H) 所示各组雄性小鼠心脏组织的 H&E 染色。(I) 所示各组雄性小鼠肺组织的 H&E 染色。(J) 空间映射显示了年轻(2 月龄)和老年(25 月龄)雄性小鼠的海马、脊髓、心脏、肺、肝脏、小肠、脾脏、淋巴结和睾丸中每个点的基因数量。

图S2. 年轻和老年雄性小鼠的九种组织的空间转录组数据摘要。
(A) 条形图显示小鼠组织空间转录组数据集的质量控制相关信息。(B) 对年轻(2 月龄)和老年(25 月龄)雄性小鼠的九个组织中的所有样本(103 个样本)的空间转录组数据进行t -SNE 可视化。(C) UMAP 可视化年轻(2 月龄)和老年(25 月龄)雄性小鼠的九个组织中的所有点的空间转录组数据。(D) UMAP 图显示批次校正前后不同批次、样品或年龄组的斑点分布。(E) 空间映射显示了 9 种小鼠组织中斑点类型和标记基因得分的代表性分布。
02
雄性小鼠组织中与衰老相关的空间组织障碍
在老化组织中,组织学分析通常会发现有序的组织受到了损害。为了分析这种现象,他们引入了“组织结构熵”(OSE) 是通过空间转录组数据集量化组织无序程度的指标(图 2 A)。在各种老化组织中,他们发现 OSE 增加,海马、脾脏、淋巴结和肝脏中的空间转录扰动水平最高(图 2 B-C)。在表现出高 OSE (R-OSEhigh) 的老化组织区域中,免疫细胞(包括小胶质细胞、巨噬细胞和浆细胞)优先富集。此外,与 R-OSElow相比,细胞身份丧失(衰老过程中的常见现象)在R-OSEhigh中更为普遍(图2D-E)。
在空间转录组学和组织学分析中都注意到老年小鼠脾脏白髓和红髓之间的界限模糊。这种结构性恶化表现为老年脾脏中白髓结构破坏和淋巴细胞分散。在老年淋巴结(另一种关键淋巴组织)中,他们检测到 T 细胞减少,同时浸润副皮质区的巨噬细胞增加,在滤泡区,B 细胞减少,但浆细胞(产生抗体的终末分化 B 细胞)增加。他们始终检测到围绕中央静脉的中央区 3 肝细胞扩散,以及门管区 1 肝细胞减少,这些肝细胞由老年小鼠中积累的脂肪细胞样特征所取代。
为了以更高的时间分辨率增强对衰老引起的结构熵变化的理解,他们从 4、13 和 19 月龄的雄性小鼠身上获取了补充组织样本,对一系列组织进行了高精度空间转录组分析,包括海马、肝脏、脾脏和淋巴结(图2F-G)。来自这些不同生命阶段的数据揭示了随着年龄的增长,OSE 不断增加,细胞身份不断减少(图2H-J)。
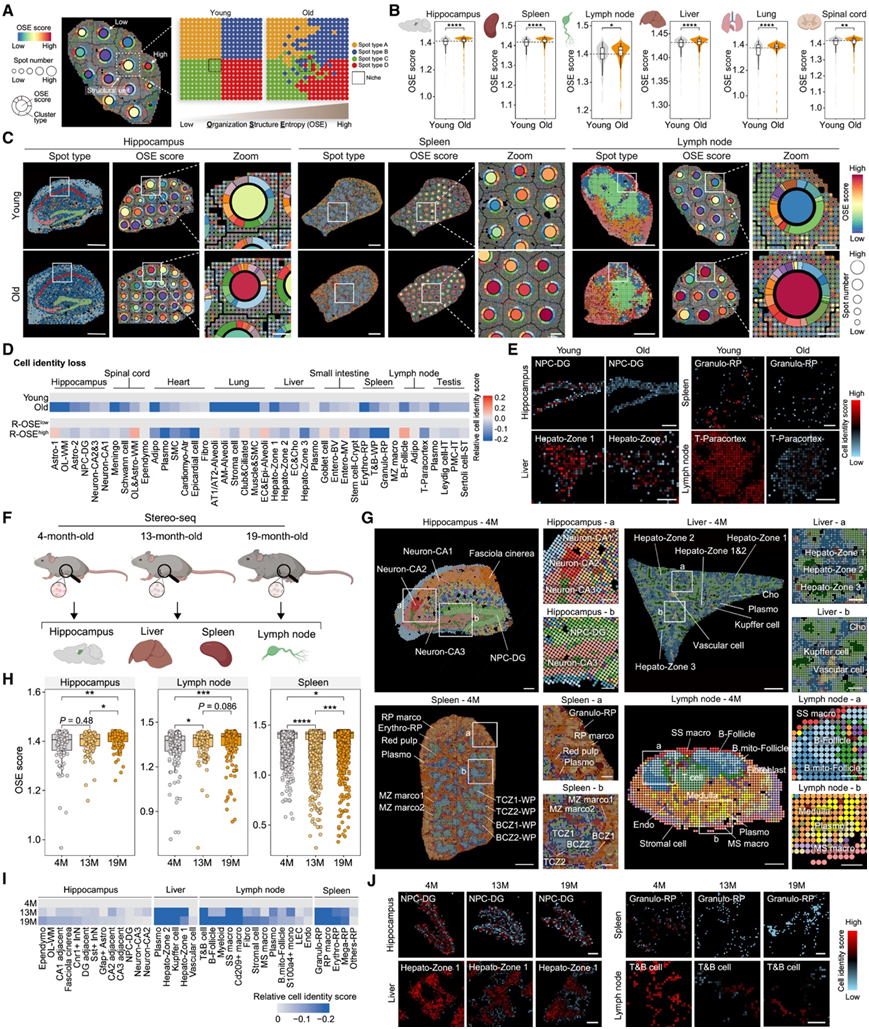
图2. 老年雄性小鼠结构恶化和细胞身份丧失。
(A) 组织结构熵(OSE)分析示意图。(B) 小提琴图显示衰老过程中 OSE 得分增加。(C) 空间映射显示年轻小鼠(2 月龄)和老年小鼠(25 月龄)组织中的 OSE 评分。(D) 热图显示老化组织和 OSE 分数较高的区域中的细胞身份分数。(E) 空间映射显示衰老过程中细胞身份评分下降。(F) 实验示意图。(G) 空间映射显示 4 月龄小鼠的斑点类型。(H) 箱线图显示衰老过程中 OSE 的变化。(I) 热图显示衰老过程中细胞身份评分下降。(J) 空间映射显示衰老过程中细胞身份评分下降。
03
雄性小鼠组织中空间分辨的年龄相关基因表达改变
接下来,为了在空间分辨率下描述细胞类型依赖的衰老相关改变,他们对年轻和老年组织之间以及不同空间位置的差异表达基因 (DEG) 进行了深入分析(图 S4A-D)。这种综合方法突出了一个关键发现:相当一部分与衰老相关的 DEG 表现出空间和细胞类型特异性的表达模式(图 S4 D)。例如,衰老会导致特定空间区域发生显著变化,包括海马 DG 中的神经祖细胞区域、脊髓中部灰质 (GM) 区域、心脏中的心室心肌细胞和肝脏中的 1 区肝细胞。在脾脏和淋巴结等淋巴组织中,白髓内的 T 细胞和 B 细胞以及滤泡中的 B 细胞分别发生了显著变化(图 S4 C-D)。值得注意的是,衰老相关的 DEG 在相似的细胞类型中也表现出空间异质性。例如,在心脏中,心室心肌细胞中此类 DEG 的数量高于心房区中的此类 DEG 数量,而在肝脏中,他们注意到 1 区肝细胞中的 DEG 数量高于 2 区和 3 区肝细胞中的 DEG 数量(图 S4 C-D)。
总体而言,衰老的 DEG 与慢性炎症、线粒体功能障碍和营养感应中断密切相关(图 S4 E,图S5 A-F)。上调的 DEG 主要与淋巴结、脾脏、小肠和肺的炎症有关,特别是老化淋巴结髓质和脾脏红髓的浆细胞(图 S4 E,图S5 A)。相比之下,下调的 DEG 与淋巴结、海马和小肠的线粒体功能相关(图 S4 E,图S5 B)。例如,与线粒体功能通路相关的基因表达减弱,以及海马 DG 和 CA1-3 区域内的神经祖细胞和成熟神经细胞中上调的凋亡相关基因过度表达(图 4 E,图S5 C)。此外,基因表达的减弱与肝脏内肝小叶区域肝细胞的线粒体功能途径和脂肪酸代谢有关,突出了能量动力学和代谢功能的空间特定功能障碍如何在衰老过程中影响肝脏(图 S4 E,S5B-F)。
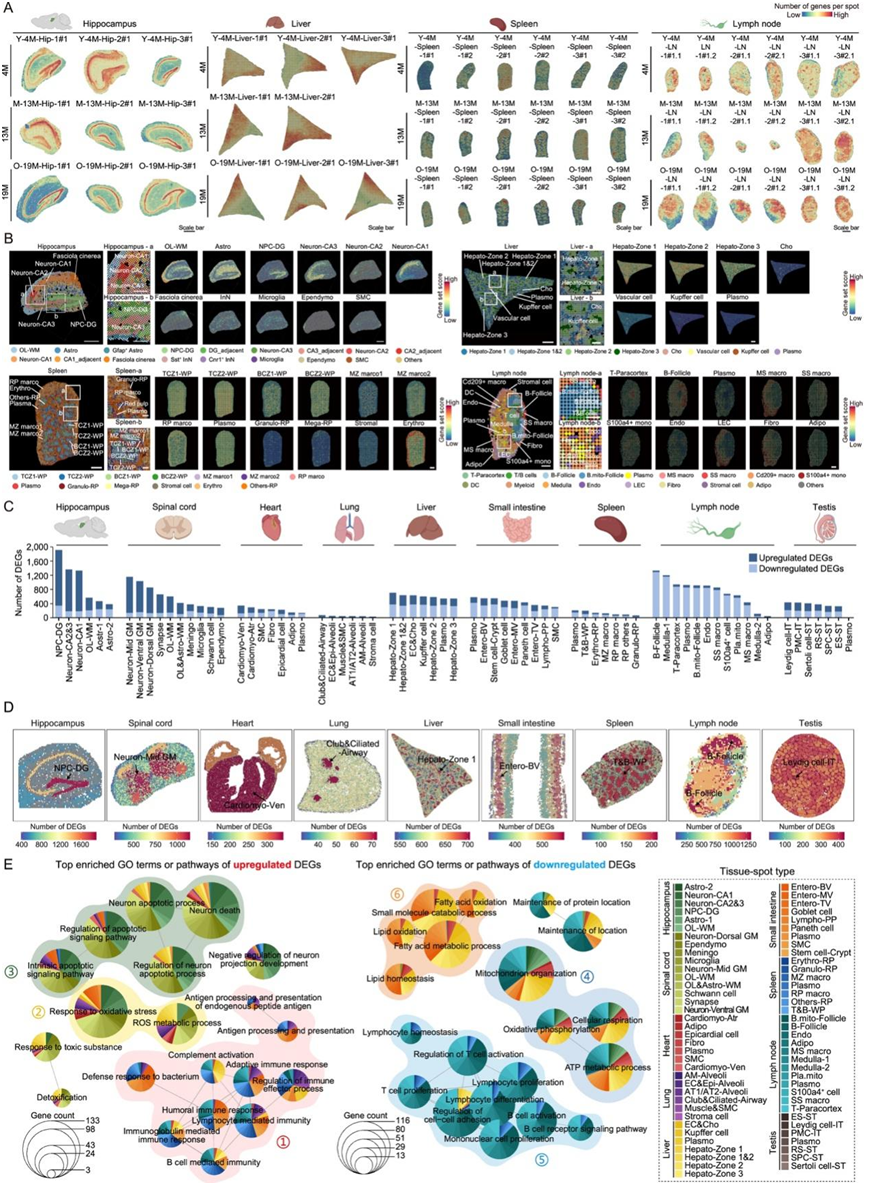
图S4. 不同年轻和年老小鼠组织中的空间尺度基因表达变化。
(A) 雄性小鼠的海马、肝脏、脾脏和淋巴结的空间转录组数据中每个点的结构和基因数量。(B) 左图为放大视图,显示 4 月龄小鼠组织中斑点类型的代表性分布。右图为放大视图,显示不同小鼠组织中所示斑点类型的标记基因得分。(C) 条形图显示空间转录组数据中不同组织的各种斑点类型中与衰老相关的差异表达基因(DEG)的数量。(D) 组织中衰老 DEG 数量的空间映射,颜色从蓝色到红色,表示 DEG 数量从低到高。(E) 网络图显示DEG 的富集通路。
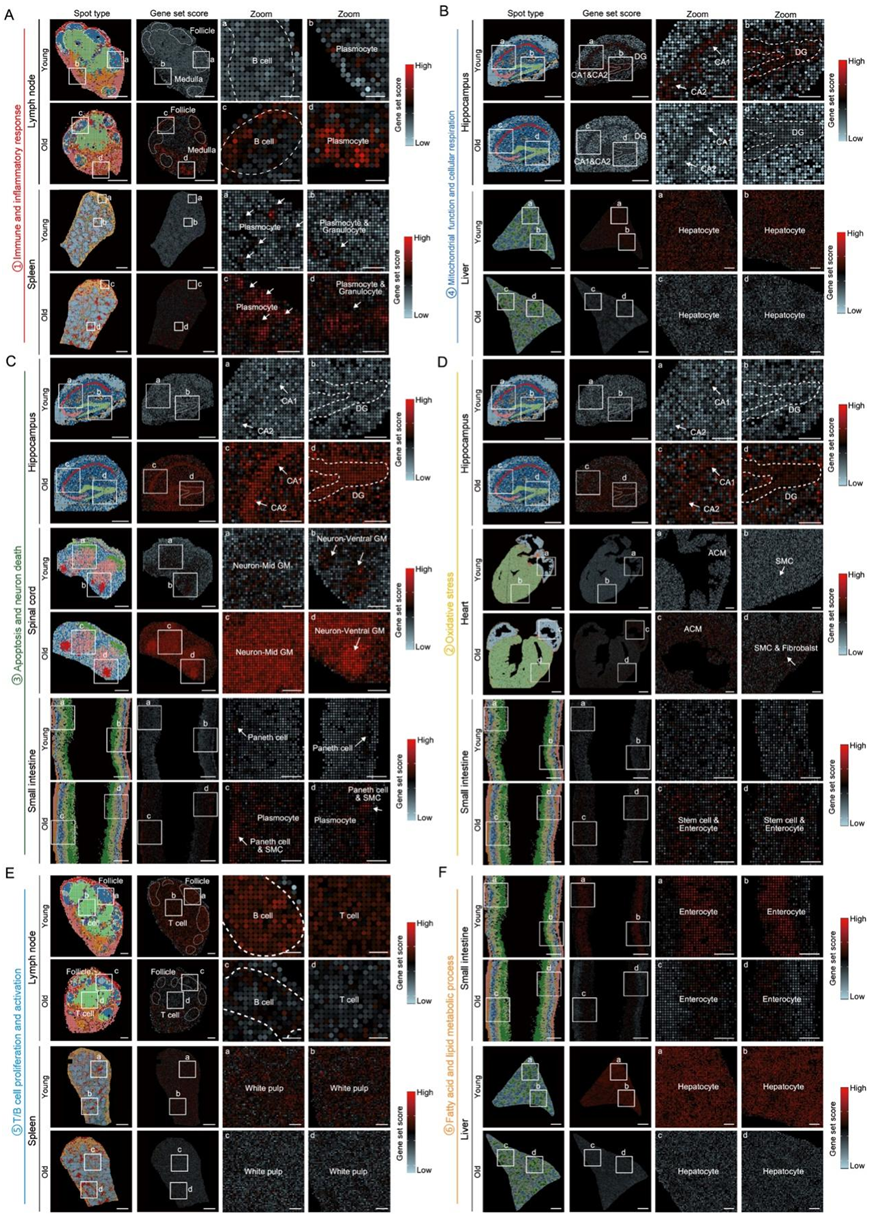
图S5. 不同年轻和年老小鼠组织之间的生物通路空间评估。
(A) 所示组织中基因集“免疫和炎症反应”得分的空间映射。(B) 所示组织中基因集“线粒体功能和细胞呼吸”得分的空间映射。(C) 所示组织中基因集“细胞凋亡和神经元死亡”得分的空间映射。(D) 所示组织中基因集“氧化应激”得分的空间映射。(E) 所示组织中基因集“T/B 细胞增殖和活化”得分的空间映射。(F) 所示组织中基因集“脂肪酸和脂质代谢过程”得分的空间映射。
04
雄性小鼠组织中衰老敏感点 (SSS)及其生态位的空间映射
接下来,他们将重点转移到精确定位衰老热点区域并描述其相邻的微环境。为了实现这一目标,他们利用了先前研究中广泛使用的五个完善的衰老基因组,并与每种组织中斑点类型特异性的 DEG 相重叠。根据衰老DEG和衰老相关基因组,他们分析了不同组织中每个空间点的衰老评分(图 3 A)。通过根据衰老评分对斑点进行排名,能够识别不同组织中对衰老高度敏感的区域。他们将每个老化样本中衰老评分最高的前 5% 的斑点指定为 SSS(图 3 B),衰老评分的强度在 SSS 中心达到峰值,并以距离依赖的方式逐渐向周围细胞降低(图 3 C)。有趣的是,OSE 分数增加和细胞身份分数降低的区域往往聚集在 SSS 附近(图3D-F)。
然后,他们根据与 SSS 的接近程度对基因表达进行排序,从而剖析了 SSS 周围的分子梯度特征(图3G-H)。在 SSS 中,体液免疫反应和补体激活升高,并且与 SSS 的接近程度与更高的 SASP 评分相关(图3G-H)。TNF 信号始终随着与 SSS 距离的增加而逐渐减弱。相反,与 ATP 生物合成过程和细胞周期相关的基因的表达水平随着与 SSS 距离的增加而恢复(图3G-H)。他们在老年小鼠的多个器官中始终观察到这种空间模式。这些结果表明,SSS 是加剧炎症的中心,以距离依赖的方式损害周围细胞。
接下来,他们在空间分辨率上研究了 SSS 的细胞身份。在海马体中,他们检测到少突胶质细胞 (OL) 和 CA2&CA3 神经元是最常见的 SSS(图 3 I)。在衰老的海马体中,SSS 的空间模式与 SPiDER-βGal 阳性细胞的分布相吻合,这些细胞优先位于神经元区域内。在脊髓中,几乎一半的 SSS 是中 GM 区域的神经元斑点,而在心脏中,心肌细胞占主导地位(图 3 I)。同样,肺中的肺泡上皮细胞、肝脏中的 2 区肝细胞和睾丸中产生雄激素的 Leydig 细胞是 SSS 的主要成分(图 3 I)。
浆细胞和终末分化 B 细胞构成了脾脏和淋巴结中的主要 SSS 部分,分别占每个组织中 SSS 的约 50% 和 30%(图 3 I)。具体而言,他们发现浆细胞 SSS 围绕着脾脏的白髓和淋巴结的髓质区(图 3 I)。此外,基于淋巴结和脾脏中的组织学染色,大多数 SPiDER-βGal 阳性细胞与 CD138 阳性细胞共定位,并且他们注意到在老化的淋巴结髓质区、脾脏的红髓和肠绒毛中积累了浆细胞。
值得注意的是,4、13 和 19 月龄时的空间转录组评估一致证实了 SSS 附近的 OSE 水平增加,并且这些区域内的细胞特征评分趋于降低(图 3 J-L)。此外,该数据集证实浆细胞是脾脏和淋巴结中 SSS 周围微环境的重要细胞成分(图 3 M)。
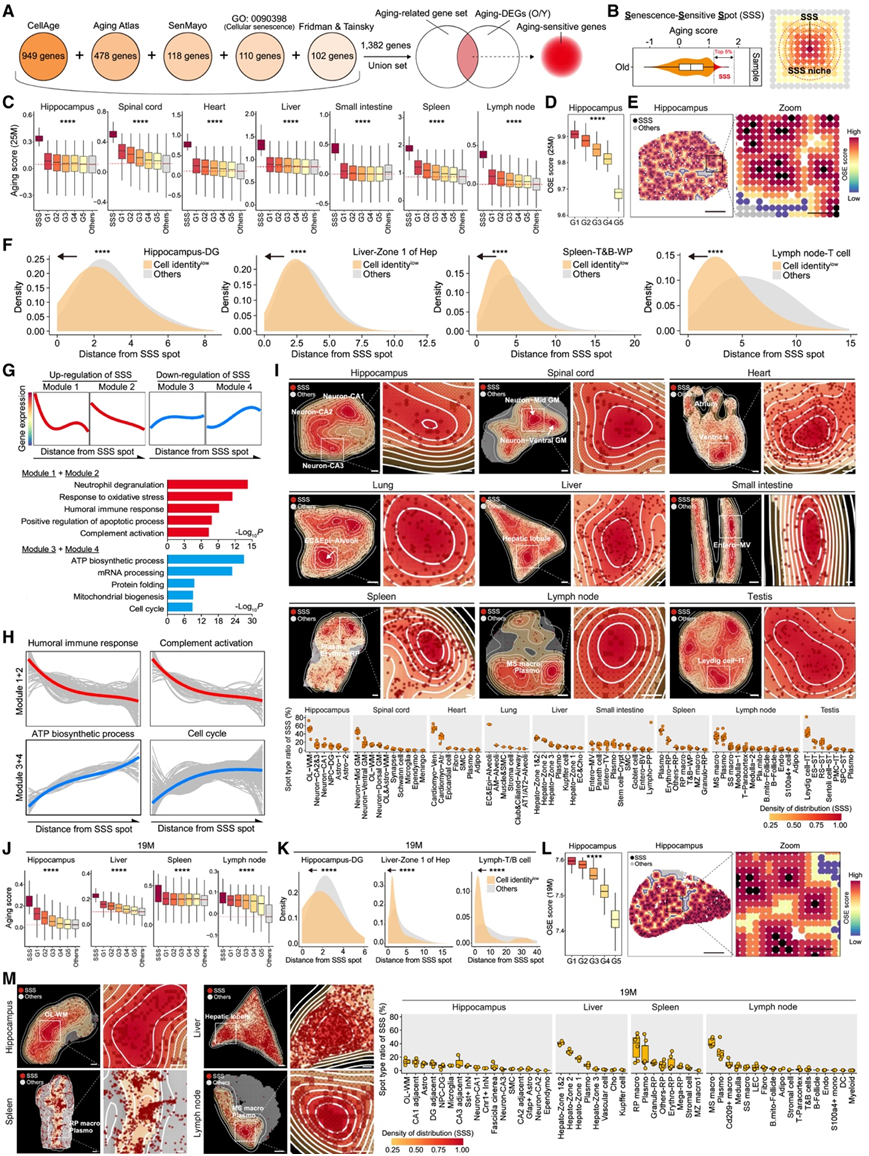
图3. 雄性小鼠不同组织中衰老敏感点的特征。
(A) 示意图显示衰老相关基因集的基因数目和衰老敏感基因的定义。(B) 显示 SSS 以及 SSS 生态位定义的示意图。(C) 箱线图显示老年小鼠(25 月龄)不同距离 SSS 的衰老评分。(D) 箱线图显示老年小鼠海马体 SSS 周围的熵值降低。(E) 空间映射显示老年小鼠海马体 SSS 周围的熵增加。(F) 密度图显示 SSS 周围细胞身份的丧失。(G) 将 SSS 周围的基因变化归类为不同的基因表达模块。(H) 模块 1 + 模块 2 和模块 3 + 模块 4 中所示通路所涉及基因表达水平的拟合曲线。(I) 上:空间映射显示 SSS 的密度分布。下:箱线图显示旧样本中 SSS 的斑点比例。(J) 箱线图显示 19 月龄小鼠衰老评分下降,同时与 SSS 的距离增加。(K) 密度图显示 19 月龄小鼠 SSS 周围细胞身份的丧失。(L) 箱线图(左)和空间映射(右)显示 19月龄小鼠海马体 SSS 周围的熵增加。(M) 左:显示 SSS 密度区域的空间映射。右:箱线图,显示 19 月龄鼠样本中 SSS 的斑点比例。
05
免疫球蛋白表达细胞和SSS建立共生系统
为了识别可能在功能上对 SSS 及其微环境产生影响的罪魁祸首基因,他们分析了最常出现的 DEG 并检查了它们与 SSS 的空间关系。在组织中,上调最多的基因是编码免疫球蛋白的基因,例如Igkc、Jchain (Igj)、Ighg1和Igha,以及编码补体蛋白的基因,例如C4b和C1qa(图 4 A)。其中,编码免疫球蛋白轻链恒定区的Igkc和由所有分泌聚合免疫球蛋白的浆细胞合成的Jchain在老化组织中增加的频率最高,尤其是在淋巴结、脾脏和小肠中(图 4 B)。他们还发现,其他免疫球蛋白相关基因(如编码 IgG 重链的Ighg2c、Ighg2b和Ighg3 )的表达在衰老组织中升高,尤其是在脾脏、淋巴结和小肠中。总体而言,在衰老组织中, Igkc基因高表达与重链编码基因表达相结合的斑点(简称Ighigh点)的比例也显著升高(图 4 C-D)。此外,大多数 Igkc 阳性细胞也在海马、小肠、脾脏和淋巴结等组织中表达 IgG。随着衰老组织中免疫球蛋白浸润的增加,老年动物血浆中的 Igkc 和 IgG 水平也升高,类似于经典的衰老相关分泌因子 IL-6(图 4 E)。
接下来,他们试图剖析免疫球蛋白生成细胞和 SSS 之间的空间关系。他们将Ighigh点分为三类:与 SSS 共定位的、与 SSS 相邻的以及与 SSS 有一定距离的相对较少的。在脾脏和淋巴结中,Ighigh点要么直接与 SSS 共定位,要么与 SSS 距离最近(图 4 F)。当对 CD138 和 Igkc 进行共染色时,在老化的淋巴结和脾脏中,大多数 Igkc 阳性细胞是 CD138 阳性浆细胞,并且 Igkc 和 CD138 双阳性细胞群在老化组织中急剧增加。
在海马、脊髓、心脏、肺、肝脏、肠道和睾丸中,与淋巴结和脾脏不同,主要的 SSS 成分不是浆细胞(图 3 I)。然而,他们注意到大量Ig高点始终位于最靠近 SSS 的外周区域(图 4 F)。此外,在这些衰老组织中,Ighigh点倾向于与富集的 SSS 聚集(图 4 F)。值得注意的是,在这些组织中,Ighigh点并非完全是浆细胞;它们是海马和脊髓中的 OL,以及心脏中的心肌细胞。空间转录组数据进一步证实了免疫球蛋白编码基因,特别是免疫球蛋白重链基因和Igkc,在各种衰老组织和器官(海马、肝脏、脾脏和淋巴结)中持续激活(图 4 G-J)。

图4. 免疫球蛋白表达细胞在雄性小鼠的整个生命周期中积累在多个组织中。
(A) 热图显示不同斑点类型中高频上调的衰老相关 DEG。(B) 免疫球蛋白结构示意图。(C) 组织中Ighigh点的比例。(D) Igkc表达和Ighigh点的空间映射。(E) ELISA 检测年轻和老年小鼠血浆中的 Igkc、IgG 和 IL-6 水平。(F) 箱线图显示Ighigh点相对于其与 SSS 距离的比例。(G) 热图显示衰老过程中上调的免疫球蛋白相关基因的表达。(H) 箱线图显示了Ighhigh点和Igkchigh点相对于它们与 SSS 距离的比例。(I) 不同年龄中Ighhigh点和Igkchigh点的空间映射。(J) 小鼠衰老过程中Ighhigh点和Igkchigh点比例增加的线性拟合。
06
免疫球蛋白积累是衰老的标志
为了动态绘制免疫球蛋白表达随年龄变化的情况,他们分析了雄性小鼠在五个生命阶段的多种组织:2、4、13、19 和 25月龄。正如预期的那样,衰老特征[例如 SPiDER-βGal 阳性细胞、P21、4-HNE、γH2AX 阳性细胞和 MMTV(人类 HERVK 的小鼠同源物)阳性细胞]随着年龄的增长在各种组织中上调。此外,他们注意到聚集体和纤维化的扩张,以及炎性细胞因子水平的升高,而层蛋白 B1 信号随着年龄的增长而减弱。与经典的衰老生物标志物相似,他们检测到雄性小鼠组织中 IgG 表达随年龄增加而增加(图 5 A-B)。他们对 Igkc 和 SPiDER-βGal 进行了共染色,发现在衰老的脾脏和淋巴结中,SPiDER-βGal 阳性细胞由 Igkc 阳性细胞紧密包围(图 S9 K)。为了验证免疫球蛋白随年龄增长的积累是否与性别无关,他们对 4、13 和 19 月龄的雌性小鼠的样本进行了染色,并观察到在衰老过程中多个器官中相似的 IgG 和 Igkc 积聚(图5 C)。相比之下,既定的抗衰老策略,如长期运动和接触年轻体液,可减轻衰老组织中 IgG 和衰老细胞的积累。
更重要的是,他们证明了 IgG 在人类淋巴结、肝脏、脾脏和大脑中与年龄相关的积累(图5D-E)。这些结果已确定了跨多个生命阶段、性别和物种的组织内 IgG 积累的一致模式。
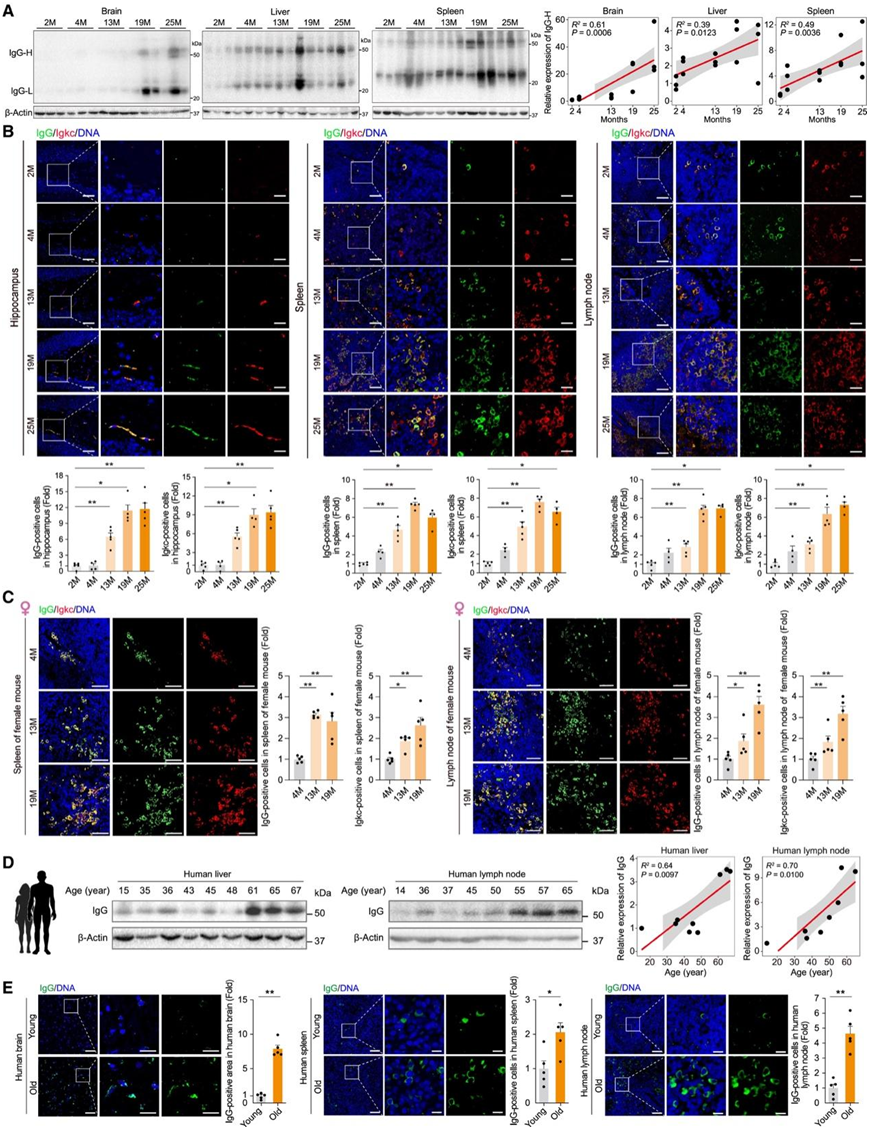
图 5.小鼠和人类多种组织中IgG 与年龄相关的积累。
(A)免疫印迹分析。(B) 5 个不同年龄组雄性小鼠海马、脾脏和淋巴结中的 IgG 和 Igkc 共染色。(C) 3 个不同年龄组雌性小鼠脾脏和淋巴结中的 IgG 和 Igkc 共染色。(D) 免疫印迹分析。(E)年轻和老年组人脑、脾脏和淋巴结中的 IgG 免疫荧光染色。
07
IgG 是促进衰老的因子
为了研究过高的 IgG 水平是否以及如何导致组织衰老,他们接下来试图确定能够识别和响应免疫球蛋白的细胞类型。在各种组织中,他们观察到组织巨噬细胞中存在的免疫球蛋白受体编码基因表达上调。此外,通过免疫荧光染色,他们观察到巨噬细胞和 IgG 阳性细胞在老年小鼠的脾脏和淋巴结中彼此非常接近。此外,相对于年轻巨噬细胞,衰老组织中巨噬细胞的转录谱显示与免疫球蛋白介导的免疫反应相关的基因上调。基于这些观察结果,他们假设与年龄相关的组织内免疫球蛋白积累很可能导致巨噬细胞转变为衰老状态。
为了验证该假设,他们分离并用 IgG 处理三种不同的巨噬细胞群(来自骨髓、脾脏和腹腔的巨噬细胞群)(图 6 A)。IgG 诱导这些巨噬细胞衰老,表现为 SA-β-Gal 活性和 P21 表达增加,同时 MMTV 阳性细胞上调、侵袭性小体形成,H3K9me3 和层蛋白 B1 水平降低(图 6 B-G)。同时,他们观察到 IgG 处理的巨噬细胞中活化的核因子 κB (NF-κB)/P65 (p-P65)、STAT1 和促炎因子分泌显著增加(图 6 H)。此外,在 IgG 治疗后,他们观察到诱导型一氧化氮合酶 (iNOS) 表达和一氧化氮 (NO) 产生增加,这两者都是促炎巨噬细胞的标志(图 6 C)。敲低编码 IgG Fc 受体的基因Fcgr4可减轻 IgG 诱导的巨噬细胞衰老(图 6 I-K),支持细胞表面的 IgG 受体在介导 IgG 的促衰老信号传导中发挥作用。同样,他们观察到 IgG 治疗可诱导人类巨噬细胞衰老(图 6 L-O)。IgG 促进巨噬细胞衰老的功效取决于其活性蛋白构象,因为热灭活的 IgG 无法诱导细胞衰老。总之,这些数据表明,当暴露于升高的 IgG 水平时,巨噬细胞会趋向于衰老和促炎状态。
接下来,他们将研究范围扩大到小胶质细胞(一种大脑中的特殊巨噬细胞),发现在衰老过程中,编码免疫球蛋白受体的基因在小胶质细胞中上调。与在巨噬细胞中的观察结果类似,用 IgG 治疗小鼠和人类小胶质细胞都会诱导一系列衰老相关标志物的表达和炎症因子的分泌(图 6 P-T)。
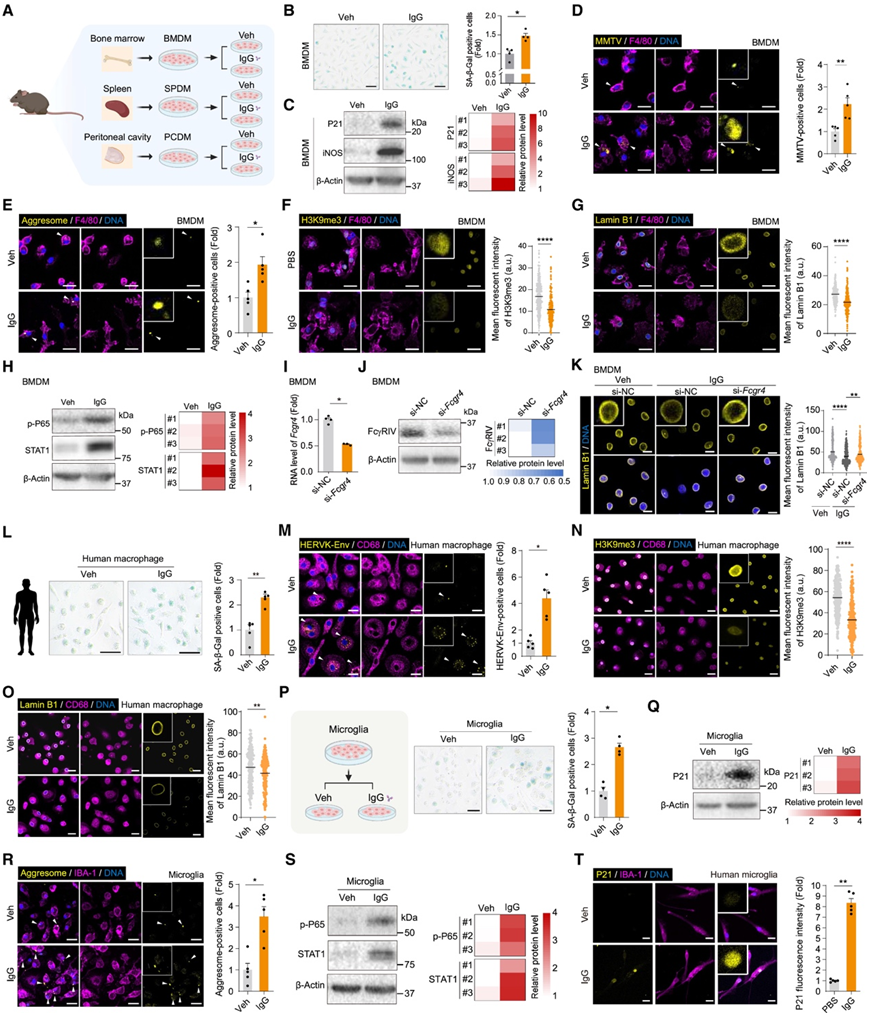
图6. IgG 引发巨噬细胞和小胶质细胞衰老。
(A) 从小鼠骨髓(骨髓来源的巨噬细胞,BMDM)、脾脏(脾来源的巨噬细胞,SPDM)和腹腔(腹腔来源的巨噬细胞,PCDM)分离的巨噬细胞的分离和处理示意图。(B) 用载体 (Veh) 或 IgG 处理的 BMDM 的 SA-β-Gal 染色。(C) 免疫印迹分析。(D) 用 Veh 或 IgG 处理的 BMDM 中的 MMTV 和 F4/80 共染色。(E) 用 Veh 或 IgG 处理的 BMDM 中的聚集体和 F4/80 共染色。(F) 左:用 Veh 或 IgG 处理的 BMDM 中 H3K9me3 和 F4/80 的共染色。右:显示 Veh 或 IgG 处理组中 H3K9me3 的荧光强度值。(G) 左:用 Veh 或 IgG 处理的 BMDM 中层蛋白 B1 和 F4/80 的共染色。右:显示 Veh 或 IgG 处理组中层蛋白 B1 的荧光强度值。(H) 免疫印迹分析。(I) 用对照或 siRNA 处理的 BMDM 中Fcgr4表达水平的 RT-qPCR 分析。(J) 免疫印迹分析。(K)左:siRNA 转染的巨噬细胞 (BMDM) 经 Veh 或 IgG 处理后的层蛋白 B1 的免疫荧光染色。右:显示各组(si-NC Veh、si-NC IgG 或 si-Fcgr4 IgG 处理)中层蛋白 B1 的荧光强度值。(L) 用 Veh 或 IgG 处理的人类巨噬细胞的 SA-β-Gal 染色。(M) 用 Veh 或 IgG 处理的人类巨噬细胞中 HERVK-Env 和 CD68 的共染色。(N) 左:用 Veh 或 IgG 处理的人巨噬细胞中 H3K9me3 和 CD68 的共染色。右:显示 Veh 或 IgG 处理组中 H3K9me3 的荧光强度值。(O) 左:经 Veh 或 IgG 处理的人类巨噬细胞中层蛋白 B1 和 CD68 的共染色。右:显示经 Veh 或 IgG 处理的组中层蛋白 B1 的荧光强度值。(P) 用 Veh 或 IgG 处理的小胶质细胞的 SA-β-Gal 染色。(Q) 用 Veh 或 IgG 处理的小胶质细胞的 SA-β-Gal 染色。(R) 用 Veh 或 IgG 处理的小胶质细胞中的聚集体和 IBA-1 共染色。(S) 免疫印迹分析。(T) 用 Veh 或 IgG 处理的人类小胶质细胞中的 P21 和 IBA-1 共染色。
接下来,他们试图验证全身 IgG 水平升高是否足以引发多种组织衰老。他们给 6 月龄的小鼠腹膜内注射 IgG,然后评估各种与衰老相关的生物标志物(图 7 A)。值得注意的是,经过 100 天的 IgG 处理后,他们观察到脾脏、淋巴结、海马、肺和心脏中 IgG 的积累(图 7 B-C)。这种 IgG 积累导致明显的衰老表型,其特征是衰老标志物 P21 水平升高以及促炎细胞因子和纤维化指标的存在(图 7 D-G)。
组织 IgG 水平通过 FcRn(Fcgrt编码)受体介导的再循环进行调节,已知 FcRn 受体的消耗会降低组织 IgG 水平。然后,他们设计了一种针对Fcgrt的反义寡核苷酸 (ASO) ,并将其腹膜内注射到 19 月龄的小鼠体内(图 7 H)。经过 40 天的处理,在脾脏、淋巴结、海马、肺和心脏中观察到 FcRn 表达显著下调,同时伴随这些组织内的 IgG 水平降低(图 7 I-L)。同时,在这些组织中观察到衰老标志物的减少(图 7 M-P)。这些结果验证了 IgG 积累在组织衰老中起关键作用的观点,并表明减少 IgG 表达的干预措施可能会改善系统性衰老。
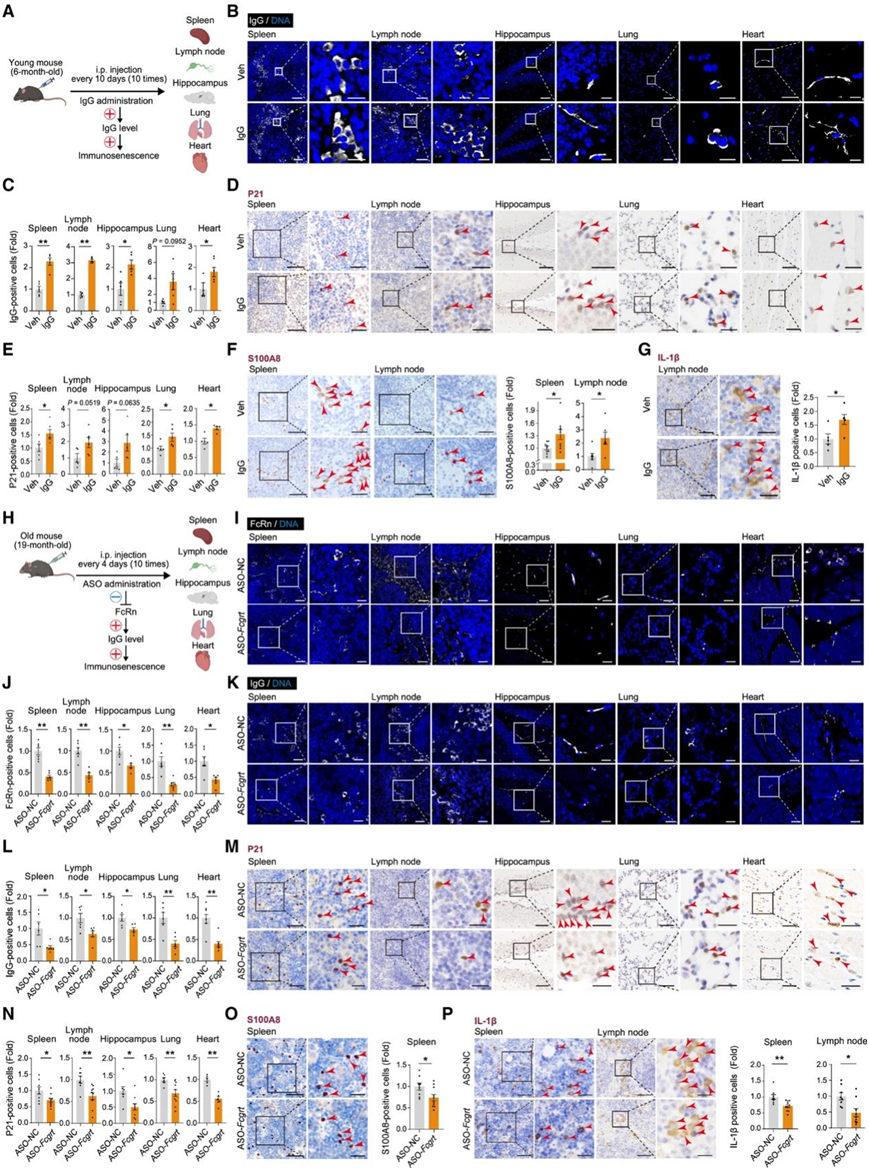
图7. IgG 的积累导致雄性小鼠多组织衰老。
(A) 用 Veh 或 IgG 处理 6 月龄小鼠的示意图。(B) 用 Veh 或 IgG 处理的小鼠多种组织中的 IgG 免疫染色。(C) 对多种组织中 IgG 阳性细胞的变化进行统计分析。(D) 用 Veh 或 IgG 处理的多种组织中 P21 表达的免疫组织化学分析。(E) 对多种组织中 P21 阳性细胞的变化进行统计分析。(F) 用 Veh 或 IgG 治疗的幼鼠脾脏和淋巴结中的 S100A8 免疫组织化学染色。(G) 用 Veh 或 IgG 处理的淋巴结中的 IL-1β 免疫组织化学染色。(H) 用 Veh 或 IgG 处理 19 月龄小鼠的示意图。(I) 用 ASO-NC 或 ASO- Fcgrt治疗的老年小鼠的多种组织中 FcRn 的免疫染色。(J) 对多种组织中 FcRn 阳性细胞的变化进行统计分析。(K) 用 ASO-NC 或 ASO-Fcgrt处理的老年小鼠的多种组织中的 IgG 免疫染色。(L) 多种组织中 IgG 阳性细胞变化的统计分析。(M) 用 ASO-NC 或 ASO- Fcgrt处理的老年小鼠的多种组织中 P21 表达的免疫组织化学分析。(N) 对多种组织中 P21 阳性细胞的变化进行统计分析。(O) 对用 ASO-NC 或 ASO- Fcgrt处理的小鼠脾脏中 S100A8 表达进行免疫组织化学分析。(P) 用 ASO-NC 或 ASO-Fcgrt处理的老年小鼠脾脏和淋巴结中的 IL-1β 免疫组织化学染色。
+ + + + + + + + + + +
结 论
本项研究对衰老过程中雄性小鼠的九种组织进行了深入的空间转录组学分析,SSS与组织结构中的熵升高共定位,而免疫球蛋白表达细胞的聚集是 SSS 周围微环境的一个特征。IgG在雄性和雌性小鼠的衰老组织中积累,在人体组织中也观察到了类似的现象,这表明免疫球蛋白异常升高可能是衰老过程中进化保守的特征。此外,IgG 可以在巨噬细胞和小胶质细胞中诱导促衰老状态,从而加剧组织衰老,并且有针对性地减少 IgG 可以减轻雄性小鼠各种组织的衰老。
+ + + + +



