

 English
English文献解读|Nat Cell Biol(17.3):三维染色质重组调节衰老过程中的 B 细胞发育
✦ +
+
论文ID
原名:Three-dimensional chromatin reorganization regulates B cell development during ageing
译名:三维染色质重组调节衰老过程中的 B 细胞发育
期刊:Nature Cell Biology
影响因子:17.3
发表时间:2024.06.12
DOI号:10.1038/s41556-024-01424-9
背 景
人们对生物体衰老过程中染色质重组的了解甚少,特别是在免疫系统中。大多数研究利用培养产生的衰老细胞作为生理性衰老的替代物。衰老细胞的染色质结构发生显著变化,包括异染色质区室 B 的去抑制、由兼性异染色质的丧失引起的从区室 B 到 A 的区域变化,以及激活炎症基因表达的染色体环路改变。衰老细胞的染色质特征在多大程度上反映生物体衰老仍不清楚。
实验设计

结 果
01

衰老骨髓祖细胞(pro)B细胞的隔室变化
研究团队建立了年轻(8-12 周)和老年(100-110 周)重组激活基因 2(Rag2)缺陷小鼠模型,染色体构象捕获(Hi-C)分析显示老年 Pro-B 细胞中区室水平相互作用(≥10 Mb)增加,而老年 Pro-B 细胞中与拓扑相关结构域(TAD)和环相关的相互作用(<10 Mb)减少(图1a-b)。在老年 Pro-B 细胞中,超过一百个基因组位点从真染色质区室 A 转变为异染色质区室 B,反之亦然[马哈拉诺比斯距离 (MD) P < 0.01](图1c)。为了探究伴随区室变化的表观遗传状态,他们进行了染色质免疫沉淀,然后用抗 H3K27ac 和抗 H3K27me3 抗体进行测序(ChIP-seq)。在从 A 转换为 B 的区域中,H3K27ac 水平降低,而在从 B 转换为 A 的区域中,H3K27ac 水平升高;然而,H3K27me3 水平没有改变(图1d)。
对年轻和老年的pro-B细胞进行转录组分析(RNA-seq)显示,位于随年龄增长从B区室转换为A区的区域中的基因平均表达水平较高(图1e)。在310个注释为随年龄增长从A区室转换为B区的基因中,只有6个在老年pro-B细胞中下调,老年pro-B细胞中A区室到B区室变化的平均基因表达没有统计学上显著差异(图1e)。
最重要的 A 到 B 转换基因组区域包含Ebf1基因,该基因编码一种转录因子,该因子是造血前体细胞向 B 谱系分化所必需的。Ebf1表达在老年pro-B 细胞中降低(图1f),同时伴有Ebf1基因座处 H3K27ac 水平降低,但 H3K27me3 不变(图1f)。他们推测,与年龄相关的Ebf1表达缺陷可能是老年小鼠 B 细胞发育受损的某些方面的原因。年轻pro-B 细胞中该区域的 Hi-C 接触图显示Ebf1启动子与标记有 H3K27ac 修饰的基因内和远端增强子之间存在多种相互作用(图1g)。这些相互作用在老年pro-B 细胞中显著减少,并由内含子和启动子远端 CTCF 结合位点之间的相互作用所取代(图1g)。
他们推测后者的相互作用反映了Ebf1表达的“关闭”状态。在祖 B 细胞阶段,随着 B 细胞的定型,Ebf1会从核边缘向更中心的位置进行径向重新定位。在老年祖 B 细胞中, Ebf1基因座移得更靠近核边缘(图1h-i)。相反,编码共同调节因子的Foxo1基因座在老年祖 B 细胞中并没有改变其径向位置或与异染色质相关 γ 卫星重复序列的接近度(图1h)。这些结果揭示了祖 B 细胞基因组在衰老过程中发生大规模构象变化,其中一些变化直接影响 B 细胞发育的关键转录调节因子。

图1. 小鼠骨髓 B 细胞祖细胞中与年龄相关的染色质区室变化。
(a) 来自年轻和老年 pro-B 细胞两个重复的组合 Hi-C 接触线性基因组距离密度图。(b) Hi-C 接触热图示例,使用 Juicebox 和覆盖率(sqrt)标准化进行可视化。 (c) 来自 Hi-C 区室分析的年轻和老年 pro-B 细胞的 PC1。(d) H3K27ac 和 H3K27me3信号的差异。 (e) 基因的差异表达。(f) 年轻和老年 pro-B 细胞中Ebf1基因座的染色质状态。(g) Ebf1基因座的 Hi-C 热图。(h) 通过 FISH 检测核定位。(i) 显示了祖 B 细胞和非 B 细胞中Ebf1基因座与核边缘之间的距离 。
02
区间变化影响Ebf1
为了评估Ebf1下调对年龄相关染色质和基因表达变化的贡献,他们首先使用Ebf1杂合子Rag2缺陷型pro-B细胞(Ebf1 +/−)进行Hi-C和转录组分析。Hi-C分析揭示了区室(≥10 Mb)、TAD和环(<10 Mb)水平的变化(图2a)。Ebf1 pro-B细胞与老年pro-B细胞有一些共同的结构特征,例如TAD内相互作用减少,但没有其他特征(图1a,图2a-b)。他们推断,Ebf1并不位于老年pro-B细胞中染色质重组的顶点。因此,Ebf1杂合性仅重现了年轻与老年pro-B 细胞之间注意到的一些差异。例如,Mmrn1基因从区室 A 转换到 B,其表达在老年pro-B 细胞和Ebf1 +/− pro-B 细胞中均降低(图2c),而在Minar1/Temed3基因座处,新的环路与老年和Ebf1 +/− pro-B 细胞中Temed3 RNA 的增加相一致(图2d)。
Ebf1 +/−祖B细胞的转录组分析也揭示了与老年祖B细胞中基因表达变化的相似性(图2e),超过50%的先前鉴定的Ebf1靶基因在老年祖B细胞中发生了改变,并且几个与B细胞发育相关的基因在Ebf1 +/−细胞和老年祖B细胞中都下调(图2f)。
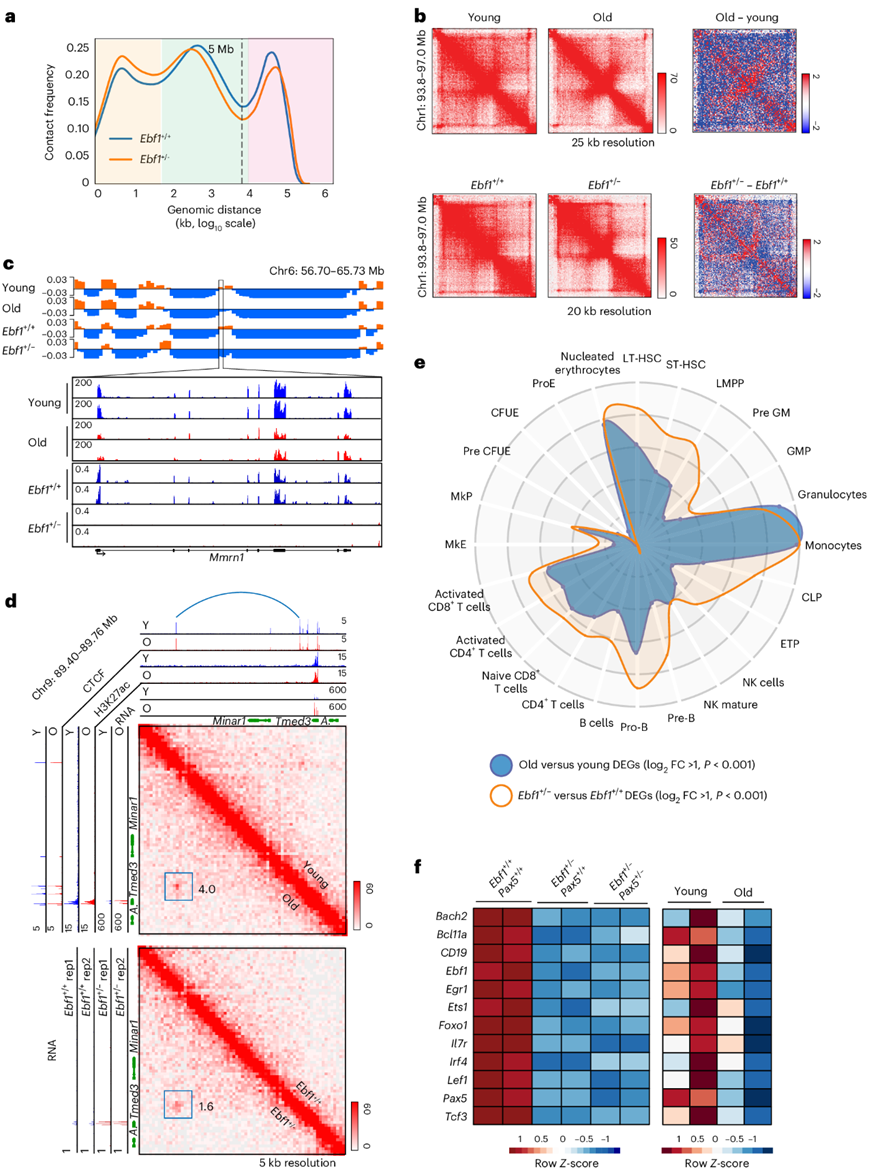
图2. 前 B 细胞中 Ebf1 表达的减少部分模仿衰老表型。
(a) Hi-C 接触线性基因组距离密度图。(b) 老年与年轻以及Ebf1 +/−与Ebf1 +/+原 B 细胞中的 TAD 变化相似。(c) 在Mmrn1基因位点,老年与年轻以及Ebf1 +/−与Ebf1 +/+原 B 细胞中的区室转换相似。(d) 老年和Ebf1+/− pro- B 细胞中相互作用增加(蓝色弧)的 9 号染色体区域 Hi-C 热图。(e) 转录变化分析。(f) 通过 RNA-seq 测量指定基因型的 pro-B 细胞以及年轻和年老(Rag2 −/− ) pro-B 细胞中关键 B 细胞基因的表达水平。
03
老年pro-B 细胞中的 TAD 改变
他们在年轻的pro-B细胞中鉴定了 2781 个 TAD。通过量化位于 TAD 边界之内或之外的双末端标签 (PET) 的比例作为 TAD“强度” (富集得分:ES)的定量,发现在老年pro-B细胞中,有 240 个 TAD 的 ES 显著较低(年轻特有),而有 84 个 TAD 的 ES 显著较高(老年特有)(图3a)。含有免疫球蛋白重链(Igh)基因位点的 TAD 在老年pro-B细胞中减少最多(图3a)。其他重要的 B 细胞调控基因,如Ebf1和Pax5,也位于减少的 TAD 内(图3a-b)。240 个年轻特异性TAD 与老年pro-B细胞中TAD体内H3K27ac减少(图3c )和TAD边界处H3K27me3减少有关(图3d)。CTCF结合没有变化;然而,老年pro-B细胞中整个年轻特异性TAD体内的Rad21水平较低(图3e,f)。老年pro-B细胞中Rad21向染色质募集的整体减少与黏连蛋白卸载蛋白Wapl表达的增加相关(图3g-h)。
几个与B细胞发育相关的基因,包括Ebf1、Pax5、Bach2和Bcl11a,位于减少的TAD内,表达水平较低(图3h),但是平均基因表达没有变化(图3i)。
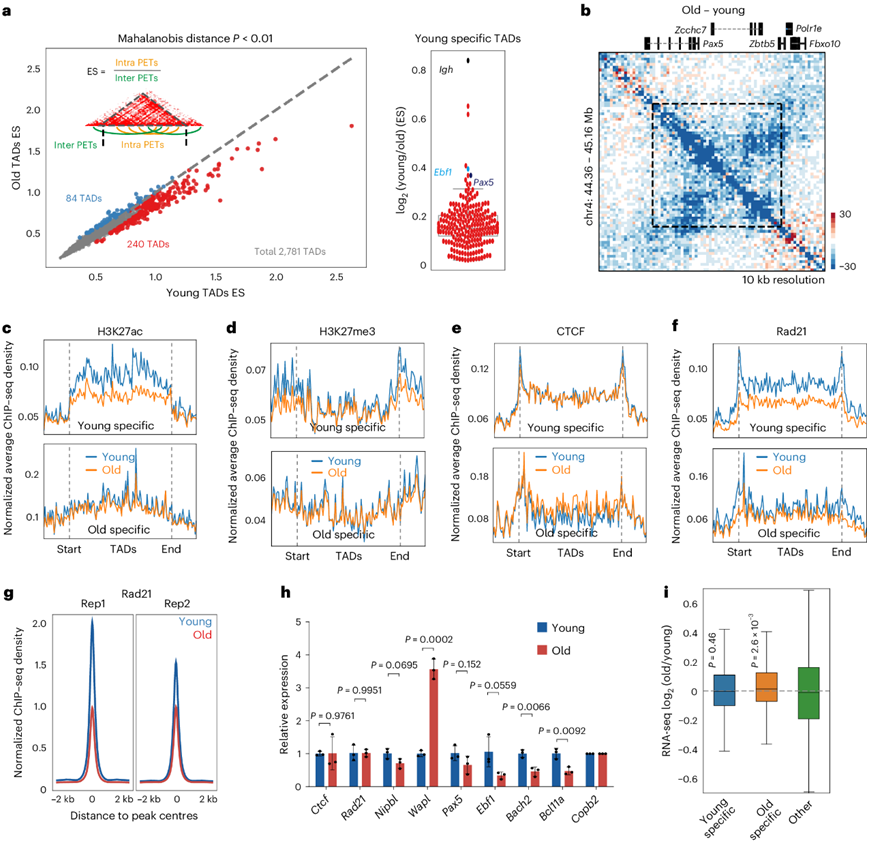
图3. 祖B细胞中TAD与年龄相关的变化。
(a) 左侧的散点图显示了基于年轻和老年Rag2−/−前 B 细胞的 Hi-C 数据的 TAD 强度的定量比较。 (b) 差异热图显示老年前 B 细胞中覆盖Pax5基因座的 TAD 形成减少。(c-f) ChIP-seq 信号密度。(g) ChIP-seq 信号密度。(h) RNA-seq 数据的定量 RT-PCR 验证。(i) 位于显著变化的 TAD 中的基因表达水平分布。
他们观察到老年pro-B 细胞中全基因组超过 10000 个位点的 H3K27ac(特征选择性)降低,大多数(78%)注释为增强子(图4a)。相反,老年pro-B 细胞中约有 1200 个位点获得了 H3K27ac(图4a)。年轻和老年pro-B 细胞中的总 H3K27ac 水平相当。年轻或老年选择性 H3K27ac 峰映射了具有相反表达模式的大部分独特基因组(图4b-d)。p300 是一种乙酰化 H3K27ac 的酶,它在老年pro-B 细胞中改变了全基因组的募集,大多数 p300 缺失的位点与老年前 B 细胞中 H3K27ac 减少的位点相吻合。相比之下,染色质重塑因子 Brg1 的募集在年轻和老年pro-B 细胞中非常相似。
他们在年轻和年老的骨髓前 B 细胞中进行了抗 H3K27ac HiChIP,以探测染色质环化。与不随年龄变化的 H3K27ac 位点相互作用相比,在年老的前 B 细胞中,涉及年轻和年老选择性 H3K27ac 位点的相互作用分别减少(年轻)和增加(年老)(图4e)。在分离增强子或启动子相关的差异 H3K27ac 位点后,这种趋势仍然保持(图4f-g)。在几个对 B 细胞发育至关重要的基因中,增强子-启动子相互作用受到年龄的影响(图4h)。

图4. 衰老过程中 H3K27ac 介导的染色质重组。
(a) 通过 ChIP-seq 在年轻和老年 pro-B 细胞中鉴定出 H3K27ac 的变化 。(b) 与年龄相关的 H3K27ac 峰注释为基因;维恩图显示包含年轻或老年选择性 H3K27ac 峰的基因之间的重叠。 (c) 包含每个 H3K27ac 峰类别的基因的表达水平。(d) 基因组浏览器跟踪包含年轻(Bcl11a和Foxo1)和老年选择性(Cebpa)H3K27ac 峰的基因。(e) H3K27ac 峰的聚合分析。(f) H3K27ac HiChIP 相互作用差异。(g) 峰的 H3K27ac ChIP-seq 信号变化与 H3K27ac HiChIP 相互作用强度之间的相关性。(h) 年轻(上)和老年(下)pro-B 细胞中Bcl11a基因座中 H3K27ac HiChIP 相互作用的热图。
04
Igh TAD 强度降低
老年pro-B细胞中减少最多的TAD包含3 Mb的Igh基因座(图5a-b)。在Igh基因座有两个显著的与年龄相关的差异。首先,老年pro-B细胞中贯穿整个TAD的“条纹”减少了(图5c)。该条纹的位置与一组ctcf结合位点(3 ' ctcf结合元件)一致,这些位点标记了Igh位点的3 '端(图5d)。其次,老年pro-B细胞中Igh TAD内的亚TAD减少了(图5c),这样的亚TAD代表了基因座中编码可变(VH)基因片段并促进远端基因片段VDJ重组的部分的某种程度的压缩。老年pro-B细胞中2.5 Mb VH区内的H3K27ac 和 Rad21 水平较低(图5d-e),与 Eμ 增强子相对应的突出 H3K27ac 峰也较低(图5d)。相比之下,CTCF 通过 VH区、基因间控制区 1 (IGCR1)和 3′ CBE的结合受年龄的影响最小(图5d-e)。荧光原位杂交 (FISH) 分析证明了老年pro-B 细胞中亚 TAD 形成减少(图5f-g)。

图5. Igh基因座染色质结构与年龄相关的变化。
(a) 小鼠Igh基因座的示意图。(b) 差异 Hi-C 热图显示 12 号染色体的一部分。(c) 年轻和年老 pro-B 细胞中Igh TAD 的接触频率热图。 (d) Igh基因座处年龄依赖性组蛋白修饰以及 CTCF 和 Rad21 结合。(e) ChIP-seq信号的定量。 (f-g) 荧光原位杂交及空间距离定量。
05
与年龄相关的Igh基因座收缩障碍的功能后果
为了更深入地探究Igh TAD内的相互作用,他们使用年轻和老年 pro-B 细胞进行了捕获 Hi-C。与全基因组 Hi-C 一致,他们发现在老年 pro-B 细胞中,TAD 内相互作用以及 3′ Igh结构域与基因座其余部分的相互作用减少(图6a)。在老年 pro-B 中,远端区域与基因座 3′ 端的相互作用减少最为显著(图6b)。为了研究 Ebf1 下调对Igh结构的影响,他们对Ebf1 +/− pro-B 细胞进行了 Hi-C 分析(图6c )。Ebf1 +/+中3′ CBE 接触到VH区中间,这可能是因为它们的增殖表型。在Ebf1 +/−细胞中,这些接触进一步减少,而与 Eμ 和 IGCR1 的相互作用不受影响(图6c-d)。Igh基因座 5′ 和 3′ 端之间的相互作用在老年pro-B 细胞中减少,部分原因是 Ebf1 表达随年龄减少。
为了确定染色质相互作用改变随年龄增长而产生的功能后果,他们对年轻和老年 C57BL/6J 小鼠纯化的 pro-B 细胞中的V H重排进行了量化。在老年 pro-B 细胞中,最近端的 V H基因片段过表达,而最远端的 V H 基因片段低表达(图6e-f)。老年 pro-B 细胞中 D H基因片段的利用率受到的影响最小。老年 pro-B 细胞中 VDJ 与 DJ 重组等位基因的比例(图6g) 和生产性 VDJ 连接的水平降低(图6h)。虽然老年 pro-B 细胞中 CDR3 区域的总数减少了,但 CDR3 长度分布并没有随着年龄的增长而改变。

图6. 与年龄相关的Igh基因座收缩障碍的功能后果。
(a,c) 捕获 Hi-C或 Hi-C的接触频率热图。(b,d) Hi-C 中得出虚拟 4C 轨迹。(e) C57BL/6J 小鼠年轻和老年 pro-B 细胞中 所有 V H基因片段的重排频率。(f) 饼状图显示年轻和老年 pro-B 细胞中近端、中端和远端 VH基因利用率的差异。(g) 显示C57BL/6J 小鼠年轻和老年 pro-B 细胞中部分(DJ H)和完全(V H DJ H)Igh重排的比例。(h) V H DJ H连接的比例。
+ + + + + + + + + + +
结 论
本项研究表明大规模染色质重组可区分年轻和年老的pro-B 细胞。这些变化导致区室水平的相互作用增加, TAD内的相互作用减少。编码 Ebf1 的基因是一种关键的 B 细胞调节因子,它会随着年龄的增长从区室 A 切换到 B。遗传上减少 Ebf1 可重现年老 pro-B 细胞的一些特征。随着年龄增长而减少最多的 TAD 包含对 B 细胞发育很重要的基因,包括Igh基因座。Igh处较弱的 TAD 内相互作用与改变的变量 (V)、多样性 (D) 和连接 (J) 基因重组相关。这些结果表明,三维染色质重组是 pro-B 细胞表型的主要驱动因素,而 pro-B 细胞表型会随着年龄的增长而损害 B 淋巴细胞生成。
+ + + + +



