

 English
English文献解读|Cell Res(44.1):单细胞时空转录组学鉴定胎儿肝脏中的 HSC/MPP 扩增单位
✦ +
+
论文ID
原名:Identification of HSC/MPP expansion units in fetal liver by single-cell spatiotemporal transcriptomics
译名:单细胞时空转录组学鉴定胎儿肝脏中的 HSC/MPP 扩增单位
期刊:Cell Research
影响因子:44.1
发表时间:2021.08.02
DOI号:10.1038/s41422-021-00540-7
背 景
在哺乳动物中,造血干细胞和多能祖细胞 (HSC/MPP) 在造血系统中至关重要,它们表现出多谱系分化和自我更新能力。
高度血管化的胎儿肝脏 (FL) 是各种造血细胞 (HC) 的短暂发育场所。在小鼠中,早期定植于 FL 的 HC 包含卵黄囊红髓系祖细胞 (EMP),它将产生红系 - 髓系谱系,包括巨噬细胞。随后,主动脉-性腺-中肾 (AGM) 来源的 HSC 迁移到 FL 中进行成熟、扩增和分化。FL 结构生态位细胞,包括内皮细胞 (EC)、基质细胞和成肝细胞,通过多种生长因子、细胞因子和趋化因子与 HSC/MPP 相互作用。
然而,对HSC/MPP在其天然生态位内扩增的细胞和分子机制的了解有限,阻碍了基于干细胞的疗法在血液恶性肿瘤中的应用。
实验设计

结 果
01

发育中的小鼠 FL 的单细胞转录组图谱
为了在转录组学水平上解决HSC/MPP及其生态位的时间动态,研究者团队以胚胎日(E) E11.5和E14.5之间的一天间隔生成发育中的FL的单细胞分辨率细胞图谱(图1a)。总共收集了 32449 个单细胞并应用用均匀流形近似和投影 (UMAP) 可视化(图 1b)。根据特征基因表达,他们注释了 21 个细胞聚类(图 1c),包括18个HC聚类和3个结构生态位细胞聚类,并发现它们的比例与发育阶段动态相关。HSC/MPP 聚类(C)1、C2、C3的特征是干细胞标志基因Mycn、Hlf和Mecom的高表达。淋巴祖细胞(C4)的特征在于Ccr9 、Il7r和Flt3表达。骨髓祖细胞 (C6) 的特征在于Mpo、Spi1和Ccl9的表达。三个结构生态位细胞群包括C19内皮细胞(Lyve1、Kdr和Pecam1)、C20基质细胞(Pdgfra、Alcam和Ncam1)和C21肝母细胞(Afp、Alb和Ttr)(图1c)。
他们接下来对四个阶段的差异表达基因 (DEG) 进行分析,并对 HSC/MPP 中的 DEG 进行基因本体 (GO) 分析,发现淋巴发育和免疫反应在 E11.5 和 E12.5 富集,造血发育和有丝分裂细胞周期调节在E13.5和E14.5处富集(图 1d-e),这表明HSC/MPP在FL发育过程中的转录动力学是HSC/MPP成熟和扩增的基础。
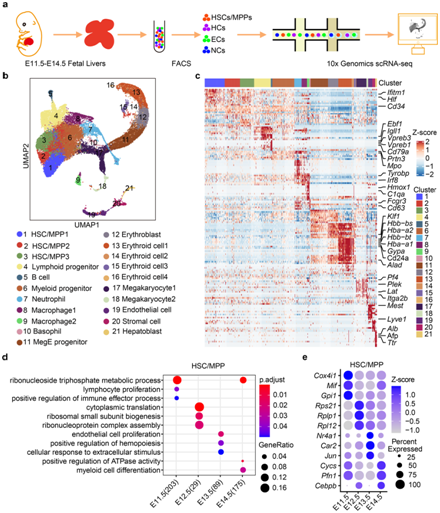
图1. 发育中的小鼠FL的单细胞转录组(scRNA-seq)图谱。
(a) FL的组织处理、细胞分离和scRNA-seq分析程序示意图。 (b) 用细胞团在FL中着色的所有单个细胞的UMAP可视化。 (c) 热图显示每个聚类前 20 个 DEG 的表达。(d) GO分析。(e) FL 发育过程中 HSC/MPP 中富集 DEG 的点图。
02
富含 CD93 的 HSC/MPP 表现出增强的干细胞特性
为了进一步剖析 HSC/MPP 的转录组特征,他们重点关注 HSC/MPP 的1-3 亚型并探讨它们之间的潜在差异。GO和DEG分析分别表明HSC/MPP的1-3亚型中造血调节和分化、主动翻译过程和细胞周期调节的富集(图 2a-b)。HSC/MPP1中Mycn、Mecom、Hlf表达富集;在HSC/MPP2中,代谢和谱系特异性基因高表达;在HSC/MPP3中,细胞周期相关基因表现出高度富集的表达(图2b-c)。为了进一步评估异质 HSC/MPP 的干细胞特性,他们计算了hscScore分数,并发现 HSC/MPP1 得分最高(图 2d)。同时,HSC/MPP亚群和下游骨髓和淋巴祖细胞之间的轨迹分析显示,HSC/MPP1占据造血发育的顶部(图 2e)。
他们鉴定了一个细胞表面标记基因CD93,其表达在HSC/MPP1中相对于HSC/MPP 2-3上调(图 2f)。
鉴CD93(也称为AA4.1)已证明在造血祖细胞中表达,于是他们想联合CD93与其他HSC/MPP标志物,如Lineage-Sca-1+c-Kit+(LSK)、Flt3-LSK和CD150+ CD48-LSK (SLAM-LSK)以进一步鉴定和纯化具有强大干细胞特性的HSC/ MPP。
为了评估CD150+CD48–LSK (SLAM-LSK)CD93+SLAM-LSK联合纯化的HSC/MPP的移植能力,他们进行原代移植试验。结果表明,E14.5 CD93+SLAM-LSK细胞比CD93- SLAM-LSK细胞表现出更高的重构和自我更新能力(图2g-j)。

图2. 富含 CD93 的 HSC/MPP 表现出增强的干细胞特性。
(a) 三种HSC/MPP亚型间的GO分析。(b) 三种 HSC/MPP 亚型中富集 DEG 的点图。 (c) HSC/MPP1 与 HSC/MPP2(左)或 HSC/MPP3(右)中所有基因表达的散点图。差异表达的转录因子是彩色的。 (d) 箱线图显示三种 HSC/MPP 亚型中的 hscScore 分布。(e) 通过PHATE进行轨迹分析,分别重建HSC / MPP向淋巴祖细胞和髓祖细胞的谱系分化。(f) 热图显示三种HSC/MPP亚型中HSC/MPP特征基因和Cd93的表达模式。 (g) 使用 CD93+ SLAM-LSK (CD93+ HSC) 和 CD93– HSC进行初次移植后 8 周,受体外周血 (PB) 中供体衍生的嵌合。 (h) 使用 CD93 + HSC 和 CD9 – HSC进行初次移植后 20 周,受体 PB 中出现供体衍生的嵌合。 (i) 使用 CD93 + HSC 和 CD93 – HSC进行二次移植后 8 周,受体 PB 中出现供体衍生的嵌合。(j) 使用 CD93 + HSC 和 CD93 – HSC进行二次移植后 16 周,受体 PB 中出现供体衍生的嵌合。
03
单细胞转录组分析(scRNA-seq) 和空间转录组学 (ST) 的整合分析 HSC/MPP 和生态位细胞之间的细胞间相互作用
在胚胎发生过程中,FL 为 HSC/MPP 发育提供了支持性生态位,然而,HSC 与其生态位成分动态相互作用的方式仍然未知。为此,他们使用 CellPhoneDB方法构建了一个HSC/MPP-生态位细胞相互作用网络,重点关注结构生态位细胞和巨噬细胞(图3a)。重要的是,他们发现了几种以前未鉴定的配体-受体相互作用(图 3b-c),包括与细胞生长相关的相互作用(FLT1-VEGFB、HGF-CD44、MDK-LPR1、IGF1-IGF1R 和 PTN-PTPRS)、细胞因子识别(CCL3–IDE 和 CCL4–SLC7A1)和 Notch 信号传导(DLK1–NOTCH3 和 DLK–NOTCH4)。
为了确定预测的信号相互作用是否确实存在于解剖学组织的细胞中,他们使用 E14.5 胚胎沿背腹轴的四个切片进行了ST,重点关注 FL 区域(图 3d- f)。去卷积后,根据前两种细胞类型的富集分数显示两个斑点图案(第一个图案和第二个图案),然后映射到原始FL区域(图3g-h)。正如预期的那样,红细胞和成肝细胞在大多数点上显示出最高的富集分数,表明它们是 FL 的主要细胞成分。
总而言之,ST 描述了 FL 的空间组织,以便进一步解分析 scRNA-seq 预测的细胞间相互作用。
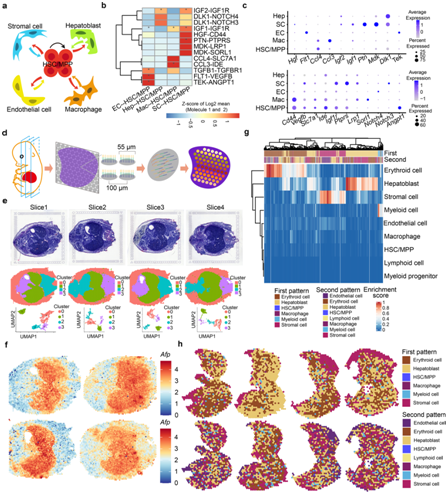
图3. scRNA-seq 和 ST 的整合分析 HSC/MPP 和微环境细胞之间的细胞间相互作用。
(a) HSC/MPP 与微环境细胞(内皮细胞、基质细胞、成肝细胞和巨噬细胞)之间的细胞间相互作用。 (b) CellPhoneDB 分析显示主要的 HSC/MPP-生态位细胞相互作用对。 (c) 主要HSC/MPP-生态位细胞相互作用对的配体(顶部)和相应受体(底部)表达模式的点图。 (d) 10×Visium ST 实验流程示意图。 (e) ST 分析中包含的胚胎天数 (E)14.5 胚胎组织切片(上)的苏木精和伊红 (HE) 染色。(f) 组织切片中肝母细胞标记物Afp的表达模式。(g) FL 截面中两个点图的空间特征图。
为了定量比较 HSC/MPP 和不同生态位细胞之间的相互作用,他们根据每种斑点类型的富集得分中位数与所有斑点的富集得分中位数的比率定义了富集倍数。他们发现巨噬细胞在点内富集11.52倍,在点间富集1.31倍,EC在点内富集1.62倍。而肝母细胞和间质细胞在点内富集较少(图4c-d)。
在分子水平上,他们检测了交互信号的空间表达,发现编码配体的基因,如MDK和PTN,在点内和点间的生态位细胞中高表达(图4e-i),以及与受体相关的基因,例如LRP1和PTPRS,在HSC/MPP定位点中富集(图 4e-i)。这些结果表明,空间邻近性促进了信号相互作用,从而在功能上支持 HSC/MPP 扩增。
鉴于点内和点间的特点是细胞之间的空间接近性和丰富的交互信号,他们将它们定义为扩增单元,其中HSC/MPP位于点的核心并与周围的生态位细胞点相互作用(图4f-i)。综上所述,他们证明 FL HSC/MPP 在多个单元中扩增,其中巨噬细胞和多种生长因子(包括 MDK 和 PTN)高度富集。
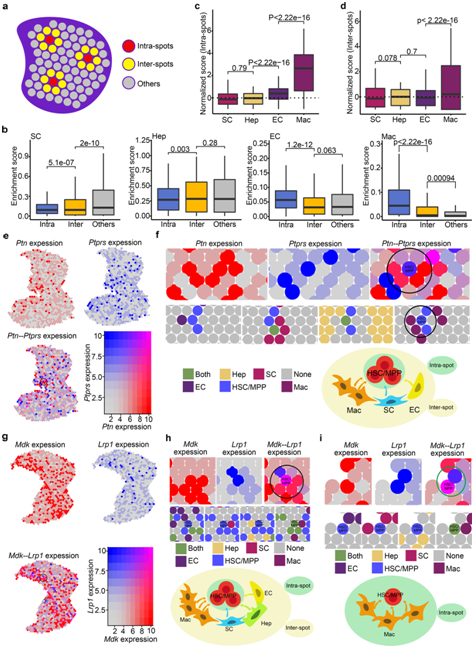
图4. HSC/MPP 扩增单元的识别
(a) 表示点内、点间等的示意图。 (b) 箱线图显示点内、点间和其他位置中生态位细胞的富集分数。(c) 箱线图显示点内生态位细胞的标准化(富集)分数。 (d) 箱线图显示点间生态位细胞的标准化(富集)分数。 (e) FL 部分中Ptn和Ptprs的共表达模式。 (f) Ptn-Ptpr的空间特征图-富集的HSC/MPP扩增单元。(g) FL 部分中Mdk和Lrp1的共表达模式。 (h) 富含Mdk−Lrp1的 HSC/MPP 扩增单元的空间特征图(点间)。 (i) 富含Mdk-Lrp1的 HSC/MPP 扩增单元的空间特征图
04
微环境细胞在促进 HSC/MPP 扩增中作用的功能验证
为了确定巨噬细胞如何调节 FL 中的 HSC/MPP 扩增,他们首先检测了 E14.5 时HSC/MPP 与巨噬细胞之间的空间关系。免疫荧光分析揭示了两种空间模式,一种是HSC/MPP附着近端巨噬细胞,另一种是HSC/MPP巨噬细胞包围HSC/MPP(图 5a-d)。值得注意的是,后一种模式类似于斑马鱼 FL 中的造血干细胞和祖细胞以及 EC 之间的模式,表明巨噬细胞可能通过形成“口袋状”结构来促进 HSC/MPP 扩张。从功能上讲,E14.5 HSC/MPP 与巨噬细胞共培养,或在补充有巨噬细胞来源的 MDK 的限定培养基中培养,发现在巨噬细胞共培养或补充 MDK 的系统中 HSC/MPP 扩增得到增强,这表明巨噬细胞可以通过分泌 MDK 因子促进 HSC/MPP 扩增(图 5e-g )。然后他们发现FL细胞的绝对数量和HSC/MPP(LSK细胞和SLAM-LSK细胞)的比例在E14.5时下降(图 5h-l)。
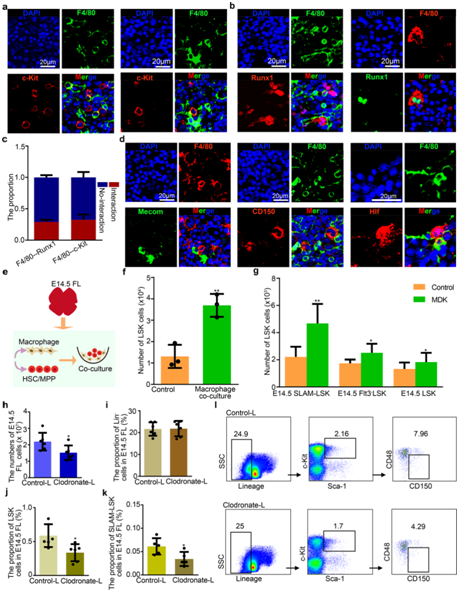
图5. 巨噬细胞促进 HSC/MPP 扩增。
(a) 免疫荧光分析显示 E14.5 FL 冰冻切片中 F4/80(代表巨噬细胞)和 c-Kit(代表 HSC/MPP)的表达。(b) 免疫荧光分析显示 E14.5 FL 冰冻切片中 F4/80(代表巨噬细胞)和 Runx1(代表 HSC/MPP)的表达。 (c) 分别具有和不具有细胞相互作用的 HSC/MPP 的比例。 (d) 免疫荧光分析显示 E14.5 FL 冰冻切片中 F4/80(代表巨噬细胞)、CD150(代表 HSC/MPP)、Mecom(代表 HSC/MPP)和 Hlf(代表 HSC/MPP)的表达。 (e) HSC/MPP 与巨噬细胞共培养的程序示意图。(f) 巨噬细胞与HSC/MPP共培养体系中获得的HSC/MPP (LSK细胞)数量。 (g) 通过添加MDK (50 ng/mL)培养100个HSC /MPP (LSK、Flt3-LSK和SLAM-LSK细胞)获得的HSC /MPP(LSK细胞)数量。(h) 氯膦酸盐脂质体和对照脂质体处理后 E14.5 FL 中的总细胞数。 (i) 氯膦酸盐脂质体和对照脂质体处理后,E14.5 FL 中谱系– (Lin–) 细胞的比例。(j) 氯膦酸盐脂质体和对照脂质体处理后 E14.5 FL 中 LSK 细胞的比例。 (k) 氯膦酸盐脂质体和对照脂质体处理后 E14.5 FL 中 SLAM-LSK 细胞的比例。 (l) 流式分析。
免疫荧光检测显示EC包围HSC/MPP(图 6a-b),并且大部分HSC/MPP位于EphrinB2标记的肝动脉门静脉附近,少数HSC/MPP位于E14.5 EphB4标记的肝静脉周围(图6c-d)。这些结果表明 HSC/MPP 显示出与 FL 中假定的门静脉血管生态位附近优先的空间关联。
为了从功能上确定来自结构生态位细胞的预测相互作用信号是否可以调节HSC/MPP的扩增,他们在E14.5时分别在添加IGF1、IGF2、HGF和PTN的培养基中进行了HSC/MPP扩增实验。鉴于IGF的功能是由其载体蛋白IGFBP调节的,他们也研究了IGFBP1/5/7的作用,它们的基因表达分别在肝母细胞,基质细胞和内皮细胞中富集(图6e)。与IGF1-、IGF2-、HGF-、IGFBP1-和igfbp7补充培养体系相比,IGFBP5-和PTN-补充培养体系中HSC/MPP的扩增明显增强(图6f-h)。综上所述,功能分析验证了潜在的细胞间相互作用和支持HSC/MPP扩增的潜在分子机制。

图6. 促进 HSC/MPP 扩增的微环境细胞结构。
(a-b) 免疫荧光分析显示 E14.5 FL 冰冻切片中 Lyve1(代表 EC)和 Runx1(代表 HSC/MPP)的表达。(c) 免疫荧光分析显示c-Kit(代表HSC /MPP)、Runx1(代表HSC/MPP)和EphrinB2(代表动脉门静脉血管)在E14.5 FL冷冻切片中的表达。 (d) 免疫荧光分析显示c-Kit(代表HSC /MPP)、Runx1(代表HSC /MPP)和EphB4(代表静脉)在E14.5 FL冷冻切片中的表达。 (e) Igfbp1、Igfbp5和Igfbp7在三种结构生态位细胞(内皮细胞、基质细胞和肝母细胞)中的表达模式的点阵图。(f) 来自 E14.5 FL 的 100 个 SLAM-LSK 细胞补充因子(每种因子 50 ng/mL)培养物的 HSC/MPP(LSK 细胞)数量。 (g) HSC/MPP(LSK 细胞)的数量源自补充因子(每种因子 50 ng/mL)的 100 个 Flt3 – E14.5 FL 的 LSK 细胞培养物。 (h) 来自 E14.5 FL 的 100 个 LSK 细胞补充因子(每种因子 50 ng/mL)培养物的 HSC/MPP(LSK 细胞)数量。
05
小鼠与人FL造血的跨物种分析
为了分析小鼠和人类之间 FL 造血作用的保守性和差异,他们将本研究中的 scRNA-seq 数据与已发布的人类 FL scRNA-seq 数据进行了比较。对受孕后11 周 (11PCW) 的人类 FL 的总共 6325 个单细胞和 E14.5 的小鼠 FL 的 8404 个细胞进行无监督 UMAP 分析(图7a)。综合分析表明,FL细胞类型和HSC/MPP多谱系(红细胞/粒细胞/淋巴细胞)分化途径在这两个物种之间很好地混合(图 7b)。
为了比较小鼠和人类之间 FL HSC/MPP 的转录组特征,他们对每个物种的 HSC/MPP 和其他细胞聚类进行了 DEG 分析,并鉴定了在人类和小鼠中保守的 39 个典型 HSC/MPP 特征基因。此外,他们对小鼠和人类 HSC/MPP 之间的 DEG 进行了 GO 分析。结果表明, “DNA损伤反应的调节”、“白细胞介素6生物合成过程的调节”和“端粒酶活性的调节”等相关通路在人类HSC/MPP中富集(图7c),证明了人类HSC/MPP 容易受到 DNA 损伤。相比之下, “炎症反应的正向调节”、“对 I 型干扰素的细胞反应”和“病毒进入宿主细胞的负向调节”等相关通路在小鼠 HSC/MPP 中腹肌,证明了小鼠 HSC/MPP对炎症信号有反应(图 7d)。鉴于小鼠和人类的生活环境不同,小鼠 HSC/MPP 可能对病原体衍生的炎症具有更强的抵抗力,而人类 HSC/MPP 可能更容易受到辐射引起的 DNA 损伤。因此,HSC/MPP 的这些不同转录组特征的存在表明,尽管总体相似,但也存在对外部刺激的适应性反应的物种特异性机制。
为了确定小鼠中 HSC/MPP 扩增的机制在人类中是否保守,他们还使用 CellPhoneDB 在人类 FL 中构建了 HSC/MPP-生态位细胞相互作用网络。他们发现HSC/MPP与巨噬细胞和EC的相互作用比与其他两种微环境细胞的相互作用更强(图 7e)。此外,他们在小鼠中,发现与细胞生长、细胞粘附、细胞因子识别和Notch信号传导相关的相互作用也存在于人类中(图 7f)。
在功能上,他们进行了免疫荧光成像,并验证了 11 PCW 人类 FL 中巨噬细胞(由 CD68 标记)和 HSC/MPP(由 CD34 标记)之间的细胞-细胞相互作用(图 7g-i)。同时,将人脐带血(CB) CD34+ HSC/MPP与小鼠巨噬细胞共培养。经过四天的共培养,发现共培养的 CB CD34 + HSC/MPP 的数量比没有巨噬细胞共培养的 CB HSC/MPP 多1.7倍以上,证明巨噬细胞人类 HSC/MPP 扩增中的保守作用(图 7j)。总而言之,这些数据与人类和小鼠之间 FL HSC 扩增的许多机制是保守的模型一致,但 HSC/MPP 中也存在物种特异性转录组特征。

图7. 小鼠和人类之间 FL 造血作用的跨物种分析。
(a) 小鼠 E14.5 FL 中所有细胞类型的 UMAP 可视化。 (b) 来自小鼠(E14.5)和人类(受孕后 11 周)的 FL 细胞群的 UMAP 可视化。(c) 对人类 HSC/MPP 中与生物过程相关的顶级 DEG 进行 GO 分析。 (d) 对小鼠 HSC/MPP 中与生物过程相关的顶级 DEG 进行 GO 分析。(e) 配体-受体相互作用网络显示人类 HSC/MPP 与微环境细胞之间的潜在通信。 (f) 热图显示主要的人类 HSC/MPP−生态位细胞相互作用对。(g-h) 免疫荧光分析显示人类受孕后 11 周 FL 切片中 CD68(代表巨噬细胞)和 CD34(代表 HSC/MPP)的表达。 (i) 分别具有和不具有细胞相互作用的 HSC/MPP 的比例。(j) 来自小鼠巨噬细胞和 HSC/MPP 共培养系统的人 CD34 + HSC/MPP数量。
+ + + + + + + + + + +
结 论
本项研究构建了小鼠胎儿肝脏(FL) 的时空转录组图谱,单细胞转录组学揭示了三个转录异质性 HSC/MPP 亚群,其中富含 CD93 的亚群表现出增强的干细胞特性。此外,通过采用单细胞和空间转录组学的联合分析,他们鉴定了新的 HSC/MPP “口袋状”单位(HSC PLUS),由生态位细胞(成肝细胞、基质细胞、内皮细胞和巨噬细胞)组成,富含生长因子。出乎意料的是,巨噬细胞在 HSC PLUS 中富集了 11 倍。在功能上,巨噬细胞-HSC/MPP共培养测定和候选分子测试分别验证了巨噬细胞和生长因子(MDK、PTN和IGFBP5)在HSC/MPP扩增中的支持作用。最后,跨物种分析和功能验证显示小鼠和人类 FL HSC/MPP 之间存在保守的细胞间相互作用和扩增机制,但转录组特征存在差异。总而言之,这些结果为了解 FL 中 HSC/MPP 的发育提供了新的理论依据,并为离体功能性 HSC/MPP 扩增提供了新的见解。
+ + + + +



