

 English
English文献解读|Nature(48.5):人类母胎界面单细胞时空分析
✦ +
+
论文ID
原名:Single-cell spatiotemporal dissection of the human maternal-fetal interface
译名:人类母胎界面单细胞时空分析
期刊:Nature
影响因子:48.5
发表时间:2026.04.08
DOI号:10.1038/s41586-026-10316-x
背 景
人类母胎界面(MFI)是一种短暂的半同种异体融合体,其中母体蜕膜基质细胞(DSC)支持胎盘附着,募集免疫细胞,并为胎儿滋养层细胞侵袭模式的形成创造耐受性微环境。在胎盘发育过程中,漂浮绒毛中的胎儿绒毛滋养层细胞(VCT)融合形成合体滋养层细胞(SCT),后者介导营养物质和代谢废物的交换以及激素和生长因子的分泌,并限制胎儿皮质醇的暴露。或者,它们形成锚定绒毛,其细胞柱产生侵袭性绒毛外滋养层细胞(EVT)。EVT侵入蜕膜和子宫螺旋动脉。在妊娠早期末期,这些血管的EVT重塑形成低阻力动脉,从而使血液能够高速流向胎盘。MFI以母体细胞和胎儿细胞的镶嵌式交错为特征。然而,其潜在的细胞、分子和空间调控机制仍未完全阐明。
实验设计
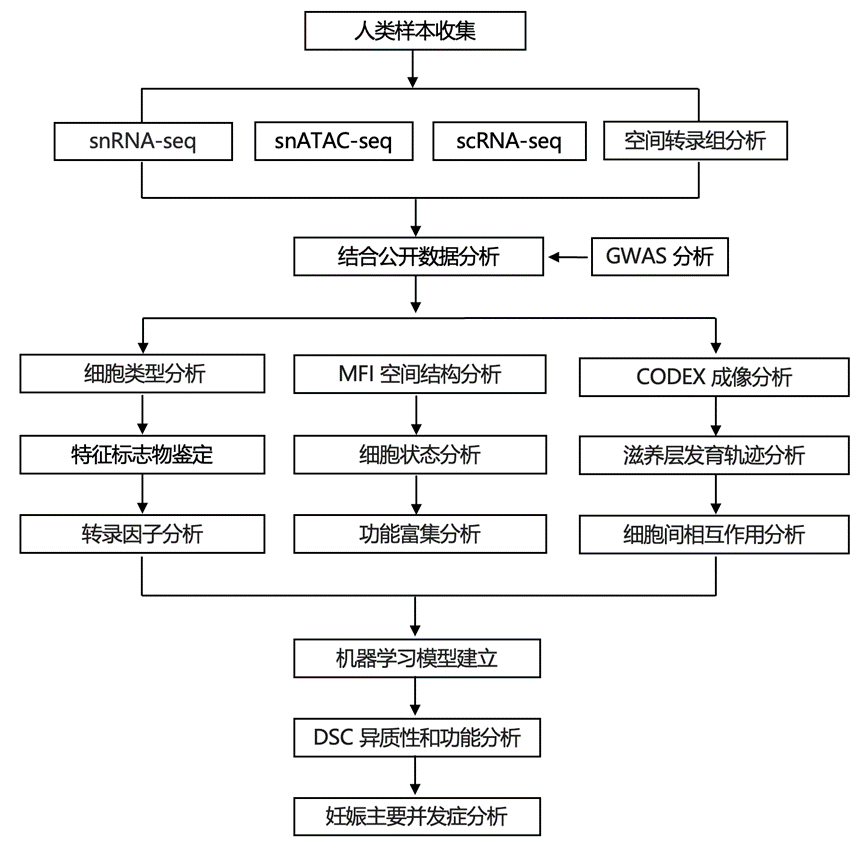
结 果
01

MFI单细胞图
本研究采用单细胞核多组学分析方法,对已知正常(足月妊娠)或推测正常(妊娠早期/中期)妊娠的MFI进行分析。样本包括妊娠5至39周的蜕膜基底层(含滋养层细胞,妊娠早期)和基底板(妊娠中期/晚期)(图1a)。研究团队进行了配对的单细胞核转录组分析(snRNA-seq)和单细胞核转座酶可及染色质高通量测序(snATAC-seq),经过严格的质量控制,获得了高质量的数据集,分别包含221380个细胞核(snRNA-seq)和210191个细胞核(snATAC-seq)。其中,191735个细胞核同时具有snRNA-seq和snATAC-seq的测序结果。平均每个样本分析了8336个细胞核的多组学数据。
他们基于已知标记对开放染色质表观基因组和转录组空间中的细胞聚类进行了注释(图1b),结果显示具有很强的细胞类型特异性(图1c),且两种模式之间总体上具有一致性,本研究发现了更多新的细胞亚型和状态。由于微观解剖结构(尤其是螺旋动脉分布)在不同的活检部位存在差异,因此在缺乏空间背景信息的情况下,跨样本的细胞类型比例比较可能会产生误导。因此,他们重点关注细胞类型内的分子特征,并且仅在有空间信息或需要空间信息时才进行成分分析。他们使用 Souporcell技术对超过 95% 的细胞进行了母源或胎源性鉴定,并通过 Y 染色体标记进行了验证(图1d)。某些细胞类型同时包含两种来源(例如,图1d中的成纤维细胞,FB),而血管内皮细胞则形成了明显的母源和胎源细胞聚类(图1b),表明存在分子分化。
细胞类型特异性基因与ATAC-seq峰的细胞类型特异性启动子分析结果进行比对。利用chromVar技术,他们推断了每种细胞类型中ATAC-seq伪整体峰中富集的转录因子结合基序。snATAC-seq足迹分析进一步解析了伪整体峰内约20 bp大小的转录因子结合位点(图1e)。然后,他们将开放染色质峰映射到实验定义的增强子。在HLA-G基因座(一种EVT标记)处,他们检测到一个EVT特异性启动子峰和三个上游开放染色质峰,其中包括一个位于上游约10 kb处的已知增强子和两个推定的远端增强子。这些分析共同揭示了广泛的细胞类型特异性调控重编程。
他们使用 CellOracle 技术整合 snATAC-seq 和 snRNA-seq 数据,重建了每种细胞类型的基因调控网络 (GRN)。重点关注滋养层谱系,以鉴定引导祖细胞 VCT 分化为终末细胞(EVT 或 SCT)的调控重编程。与 VCT 相比,分别鉴定出 71 个和 30 个在 EVT 和 SCT 中特异性上调的转录因子(图1g)和EVT 与 SCT 的重叠区域(图1h)。CellOracle 推断了这些转录因子的靶基因。在EVT中,上调最显著的20个转录因子包括ASCL2、FOS、KLF6和STAT1,它们是已知的VCT向EVT分化的调控因子,介导了167个高置信度调控相互作用,涉及116个靶基因(图1g)。其中,FOS正向调控经典的EVT基因,例如HLA-G、KRT8和FN1(图1g)。在EVT中激活的相同转录因子也与一些在EVT中下调的基因呈负相关,正如预期的那样,但其中许多实际上是已知的SCT标志物(例如CGA、TFPI2和PLAC4)(图1g)。在全基因组范围内,与 EVT 转录因子呈正相关的基因在 EVT 中表达上调,而呈负相关的靶基因在 EVT 中表达下调,但在 SCT 中富集(图1i)。在 EVT 中,EVT 转录因子与 SCT 基因之间的这些负相关性(预测为潜在的抑制性相互作用)与近期报道一致,即 EVT 特异性转录因子在 EVT 中功能性地抑制 SCT 程序。对 SCT 特异性转录因子的相同分析显示了相反的模式:激活 SCT 程序,同时在 SCT 中与 EVT 富集的基因呈负相关(图1i)。综上所述,这些观察结果提示了一种双稳态切换开关模型,该模型强制细胞分化为EVT或SCT谱系,将细胞锁定在一种命运,同时主动抑制另一种命运(图1j)。值得注意的是,这两个谱系之间仅有少数激活转录因子是共有的(图1h),包括GCM1,其缺失会损害EVT和SCT的形成。即使是这些共有的因子,在两种细胞类型中也激活了不同的靶基因集,这凸显了广泛的调控重编程,从而确保了互斥的滋养层细胞命运。
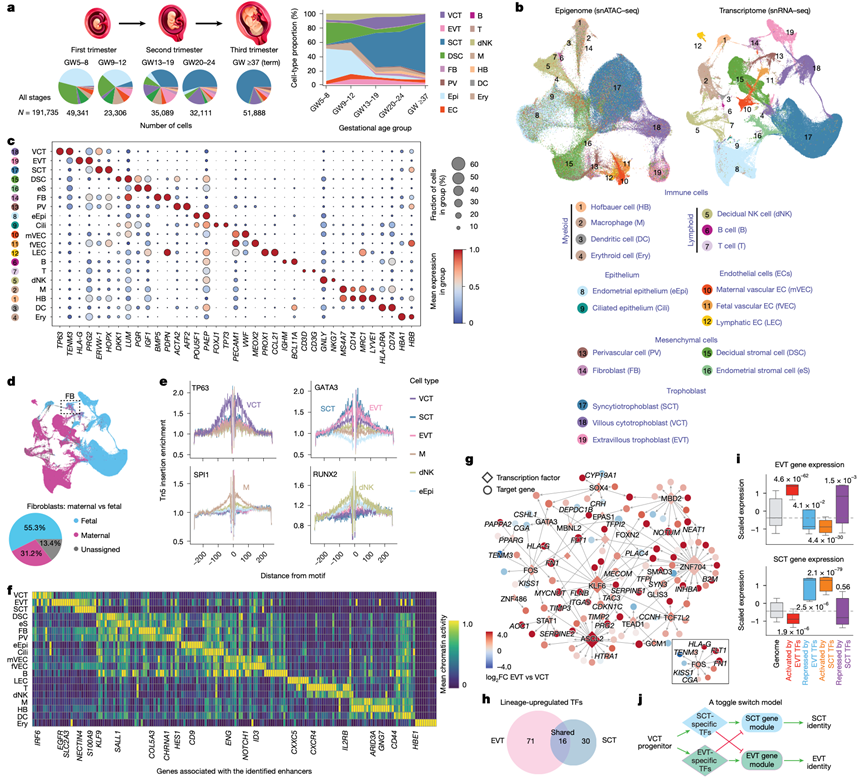
图1. 人类妊娠期 MFI 单细胞多组学数据概览。
(a) 实验设计。(b) 19种广泛鉴定的细胞类型(图例)的snATAC-seq(左)和snRNA-seq(右)的UMAP投影图。(c) 19种细胞类型中选定标记基因的表达(snRNA-seq)散点图。(d) 根据基因型相位将每个细胞核分配为母体或胎儿细胞。(e) ATAC-seq足迹分析鉴定出细胞类型特异性转录因子的结合位点。(f) 染色质可及性热图显示了十个活性最高的细胞类型特异性增强子的活性。(g) VCT分化为EVT和SCT谱系过程中基因调控网络(GRN)分析。(h) 维恩图显示了与VCT相比,EVT或SCT中特异性上调的转录因子(TF)。(i) 转录因子及其靶基因的表达模式反映了两个滋养层谱系间的协同调控。(j) 示意性模型,展示了滋养层命运的强化和替代程序的抑制。
02
从单细胞到组织结构
为了在天然组织环境中对深度测序的细胞状态进行空间映射,他们使用 STOmics Stereo-seq对来自正常妊娠的 16 个宽幅基底板活检切片生成了亚微米(0.5 µm)空间全转录组图谱,每个切片均来自不同的供体(图2a)。Stereo-seq能够从全组织切片中分割出单个细胞,并通过聚合以 0.5 µm 空间分辨率捕获的转录本来重建单细胞全转录组。他们重点关注 EVT 侵袭和螺旋动脉重塑的高峰期,仅分析了妊娠中期基底板样本,这些样本包含清晰的 MFI,并具有锚定绒毛和丰富的螺旋动脉。该技术允许在转录组分析之前进行免疫染色。大多数样本(16个样本中的12个)使用抗泛细胞角蛋白 (CK) 抗体进行免疫染色以标记滋养层细胞,其中一部分含有大量血管的样本还使用抗CD31抗体进行共染色以标记血管系统。其余 4 个样本特意未进行免疫染色,以评估其对转录组质量的影响,并验证仅使用转录组进行注释的结果。广泛的质量控制证实了数据质量,并验证了制造商的建议,即预运行免疫染色不会影响 Stereo-seq 的性能。细胞类型分配在不同的技术差异(例如染色方案和化学试剂版本)下保持一致,表明不同制备方法之间具有一致性,并证明了整合所有 16 个全切片样本进行后续分析的合理性(图2a)。
空间分析重建了所有样本中约110万个细胞;它们的转录组经过协调处理,并嵌入到统一的均匀流形近似和投影(UMAP)模型中,该模型重现了主要细胞类型(图2b)。他们进行了空间共现分析,以识别蜕膜中显著共定位的细胞群,结果显示蜕膜基质细胞和免疫细胞之间存在最强的关联,提示基质细胞与免疫细胞之间存在相互作用。接下来,他们进行了空间细胞群落分析,以识别基本的“邻域”,即在不同组织中反复出现且具有共同特征细胞类型组成的相邻细胞的空间微环境。该分析揭示了六个主要细胞群落,每个群落对应于一个独特的解剖学微环境:两个蜕膜微环境(D1 和 D2)、母体动脉和胎儿绒毛核心血管微环境、母胎连接处以及漂浮绒毛微环境(图2c)。每个微环境都具有其特有的细胞类型组成(图2d)。
蜕膜微环境D1和D2截然不同。D1富含EVT,而D2的EVT较少,主要由蜕膜基质细胞和免疫细胞组成(图2d)。血管群落也存在差异。母体动脉微环境富含可能参与血管重塑的EVT,而绒毛核心血管微环境则富含ACTA2+血管周围细胞。在母胎交界处,锚定绒毛的VCT和SCT与蜕膜基质细胞和内皮细胞交织在一起,而漂浮绒毛微环境则由SCT、VCT和缺乏血管周围鞘的胎儿内皮细胞组成(图2d),这与滋养层基底膜下的毛细血管网络相符。因此,空间群落分析勾勒出了构成人类 MFI 基本结构单元的反复出现的细胞生态位。
对于每个细胞,他们计算了其到两个解剖标志的像素距离(像素大小为 0.5 µm):最近的 MFI 部分和最近的母体螺旋动脉。在每个样本中,首先将界面定义为胎盘和蜕膜之间致密锚定绒毛末端形成的边界。然后,根据到界面的距离对细胞进行分组(图2e),并分析了跨样本的按距离分层的细胞类型组成谱(图2f)。总体而言,EVT 在浅层蜕膜中比在深层蜕膜中更丰富,这与只有最具侵袭性的 EVT 才能到达这些深度相一致。然而,免疫细胞和 DSC 在深层蜕膜中富集。绒毛膜绒毛内的胎儿细胞类型组成变异性较小,与其到界面的距离无关(图2f)。
在所有切片中,CD31免疫染色与PECAM1表达(CD31由PECAM1编码)共同勾勒出62条子宫血管。详细的观察证实了螺旋动脉的形态以及邻近CK+ EVT的预期富集。以血管壁为零距离,他们定量了同心100像素距离区间内的细胞类型组成(图2g),将正距离赋值给管腔外细胞,负距离赋值给管腔内细胞。延伸至绒毛膜绒毛的远端细胞将排除在分析之外。管腔和血管周围区域富含母体内皮细胞和滋养层细胞,仅有少量免疫细胞(图2h)。向外延伸,内皮细胞减少,滋养层细胞仍然存在;蜕膜细胞和免疫细胞是主要的邻近细胞群。
为了定量评估EVT的聚集情况,他们以随机抽取的、大小相当的、不含子宫血管的蜕膜区域为参照,估算了EVT的密度,结果显示子宫血管周围EVT显著富集(图2i)。来自同一样本的两条代表性血管的放大图突显了整合免疫染色和空间转录组学在捕捉EVT介导的血管重塑不同阶段方面的强大能力(图2j)。第一条螺旋动脉(图2j)显示出基本完整的CD31+内皮细胞层,仅有少量CK+ EVT聚集在一侧(图2j),表明其处于重塑的早期阶段。相比之下,第二条血管(图2j)呈现出嵌合型EVT表型:NCAM1+(血管内EVT标志物;图2j)EVT共表达不同水平的HLA-G(图2j)和AOC1(间质EVT标志物)(图2j)。残余的CD31+内皮细胞沿血管壁稀疏且呈碎片状分布,这与EVT介导的移位和晚期重塑相一致。蜕膜基质细胞和散在的免疫细胞也存在,提示血管重塑过程中存在多细胞协同作用。
在取自螺旋动脉的010-GW20样本中,他们鉴定出17个穿过蜕膜的横截面,每个横截面上血管壁内皮细胞和EVT的密度各不相同(图2k)。单个样本中的这一系列横截面使得他们能够详细绘制内皮细胞状态转变图谱,而不会受到样本间差异的干扰。随后,在其他样本中重复了这些转变,并在蛋白质水平上进行了验证。他们使用了PDE3A,一种近期发现的动脉内皮标记物,并通过重新分析和子宫螺旋动脉的免疫染色对其进行了独立验证,以确认这些血管内衬有动脉内皮。对这些动脉内PECAM1+动脉内皮细胞进行全转录组分析,并进行拟时间重建,解析出四个连续状态(图2l)。相邻状态之间的差异表达分析表明,PDE3A和VIM的共表达区分了每种细胞状态(图2m):典型的动脉内皮细胞(caEC;VIMhiPDE3Ahi),依次经历 R0(VIMlowPDE3Ahi)、R1(VIMlowPDE3Alow)和 R2(VIMhiPDE3Alow)(图2l-m),阐明了内皮细胞对 EVT 介导的重塑的反应。功能富集分析证实了这一轨迹方向:与 caEC相比,R2中上调的基因与细胞凋亡显著相关(图2n),证实 R2 为终末状态。在独立的组织样本中观察到了相同的细胞状态(R0、R1 和 R2)(图2o-p)。值得注意的是,促凋亡基因GADD45G的高表达特异性地出现在 R2 终末状态(图2o)。空间测量进一步验证了这一轨迹(图2l)。他们计算了每个内皮细胞与血管壁的绝对距离。caEC 和 R0 细胞彼此靠近,R1 细胞略微偏离,而 R2 细胞距离最远(图2q)。因此,螺旋动脉重塑过程中 caEC 的丢失始于 R1 状态下的血管壁移位,导致 R2 细胞完全脱离、凋亡并最终清除。
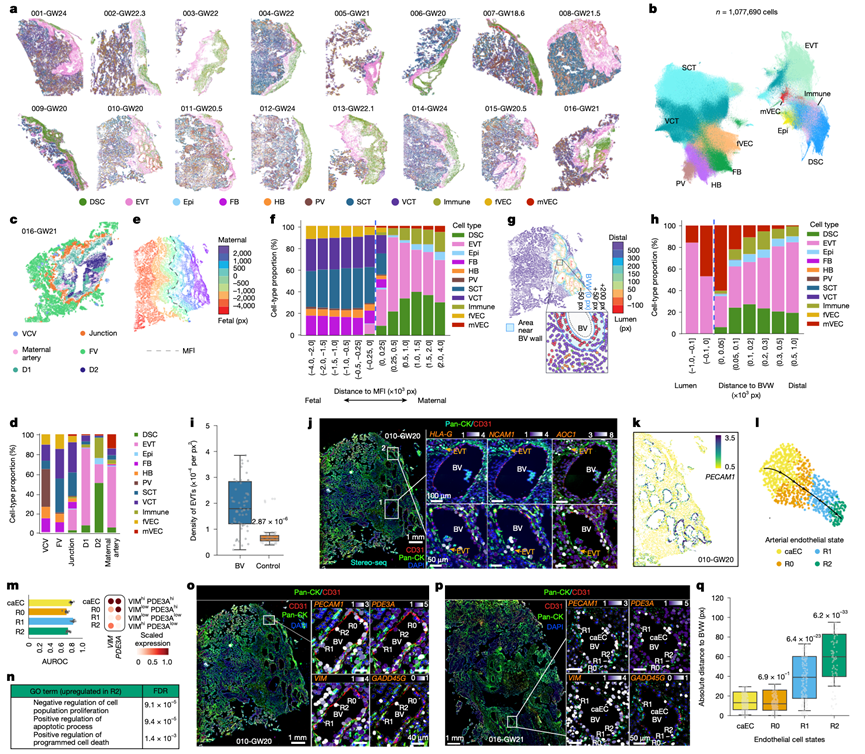
图2. MFI 的亚微米空间全转录组分析概述。
(a) 对16例正常妊娠中期样本的基底板切片进行STOmics空间测序分析。(b) 对16个样本中约110万个单细胞空间转录组谱进行UMAP投影。(c) 空间细胞群落分析鉴定出六个重复出现的群落。(d) 每个空间群落均表现出特征性的细胞类型组成。(e) 每个细胞到MFI的距离(以0.5 μm像素为单位)。(f) 按MFI分组的细胞类型比例。(g) 蜕膜中细胞与最近血管壁的空间距离。(h) 蜕膜中细胞类型与血管壁(BVW)的接近程度比例。(i) 血管壁附近滋养层细胞(EVT)密度的定量,并 与相同大小的随机蜕膜区域进行比较。(j) 通过免疫染色和空间转录组学联合可视化,展示了处于EVT介导的重塑不同阶段的两条代表性螺旋动脉。(k) 代表性GW20样本中EVT细胞活跃侵袭。(l) 单细胞转录组分析揭示了内皮细胞状态的转变。(m) PDE3A和VIM能够高精度地区分R0-R2状态。(n) R2内皮细胞富集凋亡相关基因。(o-p) 通过配对的空间转录组学和泛CK及CD31免疫染色。(q) 17条重塑螺旋动脉(孕10-20周)内皮细胞状态与血管壁的绝对像素距离。
03
CODEX 对螺旋动脉 EVT 重塑过程中动脉内皮状态逐步转变的验证
接下来,他们采用 CODEX 成像技术,通过多重检测九种细胞类型特异性抗体进行蛋白质水平验证,该方法能够准确检测人 MFI 处的主要细胞类型(图3a),包括 CD31 ( PECAM1 )、PDE3A 和波形蛋白 (VIM),用于标记不断变化的内皮细胞状态。在蛋白质水平上,在血管重塑不同阶段的动脉中观察到了细胞状态(R0、R1 和 R2)(图3b-e)。这些 CODEX 观察结果已通过对独立样本的免疫定位进行了验证。
为了进行功能表征,他们将每种内皮细胞状态与 caEC的空间转录组进行了比较(图2l)。R0 细胞表现出连接基因和抗原呈递通路的下调(图 3j),这定义了一种血管壁黏附的“启动”状态(图2q),其结构稳定性降低,免疫信号传导减弱。R1 细胞下调了PDE3A和其他动脉标志物(例如EFNB2、HEY1和DLL4)(图3k),表明动脉特性丧失,并伴有早期从血管壁脱离(图2q)。R2,即终末EVT重塑状态,表现出凋亡基因激活(图2n)和完全脱离(图2q,图3f-i),提示发生失巢凋亡。值得注意的是,caEC距离EVT最远,而EVT逐渐靠近R1和R2,这支持了一种空间梯度,即EVT距离的增加驱动内皮细胞从失去动脉特性(R1)到凋亡(R2)的连续转变(图3l)。
在该模型(图3l)中,SA-A细胞(图3b)处于重塑前的准备状态(R0)。SA-B细胞的重塑程度更高,上壁分布着R1细胞,下壁则分布着由EVT细胞取代的R2细胞。图3d中的动脉代表了接近终末期的重塑过程,此时EVT细胞已基本取代了内皮细胞。
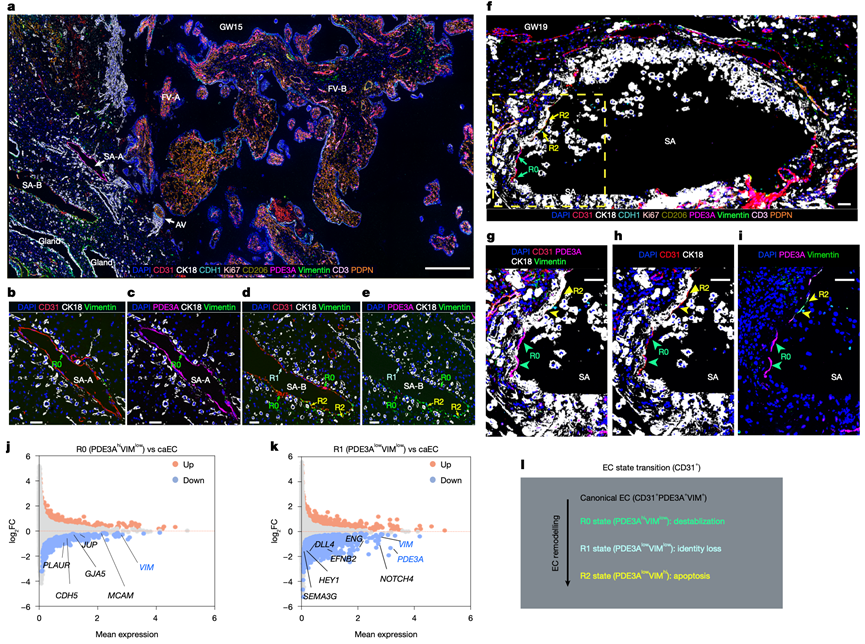
图3. CODEX 对螺旋动脉 EVT 重塑过程中动脉内皮状态逐步转变的验证。
(a) 对人类基底板样本(妊娠15周)代表性组织切片进行多重CODEX成像的概述。(b-e) 螺旋动脉 SA-A 的高倍率 CODEX 成像。(f) GW19 基底板样本中广泛重塑的螺旋动脉附近的 CODEX 成像概览。(g-h) 显示了选定通道中不同内皮细胞状态的细节。(i) R0细胞(绿色箭头)和 R2 细胞(黄色箭头)在空间上是分离的。(j-k) 差异表达基因分析比较了空间转录组谱。(l) 提出的螺旋动脉重塑过程中内皮细胞(CD31 +)状态转变的逐步模型。
04
滋养层发育轨迹
深度覆盖的snRNA测序能够灵敏地识别滋养层中间状态,随后利用空间转录组学对其进行原位定位。基于snRNA-seq数据对95872个滋养层细胞进行聚类,鉴定出由典型标记区分的VCT、SCT和EVT亚群(图4a-c)。在每个谱系中,他们发现了多个随妊娠阶段变化的中间细胞状态(图4d)。使用Palantir 进行的伪时序分析重建了VCT向SCT和EVT的典型分化过程(图4e),并解析了滋养层状态在发育连续体中出现的时间(图4f)。首先,他们重点研究了EVT谱系(图4e),该谱系起源于一种VCT亚型——锚定绒毛VCT(图4d-f),并与另一种亚型——主要分化为SCT的漂浮绒毛VCT(图4d-f)有所区别。他们鉴定出一个以ITGA2、ITGB6和VIT表达为标志的EVT祖细胞群(图4e),并进一步区分出三种终末EVT亚型:血管内EVT(eEVT)、间质EVT(iEVT)和血管周围EVT(pEVT),每种亚型均由已知的标志物定义(图4e)。此外,他们还鉴定出一个以CD81为标志的潜在滋养层巨细胞群(图4d)。这些细胞也表达典型的iEVT 标记,表明它们是由 iEVT 形成的。
接下来,他们利用聚合的空间数据集,根据与MFI和血管壁的距离,对EVT亚型进行定位(图4g-h)。EVT祖细胞局限于界面附近的区域,包括一个仅限于细胞柱的ITGB6 +亚群(图4i)。eEVT的比例随蜕膜深度的增加而增加,iEVT是蜕膜各层中的主要亚型(图4g),并在离开细胞柱时开始表达HLA-G(图4i)。在iEVT中,PRG2 +和AOC1 +亚群(图4e)在空间上相互交错(图4i)。AOC1在发育较晚的iEVT中富集(图4e ),这与其作为晚期EVT标志物的作用相符。根据血管邻近程度进行分层,NCAM1+ eEVT局限于管腔内和血管周围区域,同时伴有 iEVT 减少;而SLIT2+ NCAM1− pEVT(图4e)则特异性地富集于血管周围区域(图4h)。这些观察结果揭示了一种镶嵌式的 EVT 组织结构,其中HLA-G+iEVT包裹血管,SLIT2+pEVT占据血管周围区域,而NCAM1+ eEVT则分布于管腔和血管壁(图4i)。
他们比较了终末分化的eEVT、iEVT和pEVT的转录组(图4j),CellChat 分析整合了配体-受体共表达和空间邻近性,揭示了eEVT和母体血管内皮细胞(mVEC)之间的相互信号传导(图4k)。内皮细胞向eEVT发出的信号可能将eEVT募集到螺旋动脉,而eEVT向内皮细胞发出的信号可能促进caEC向R2的转变(图3f)。该分析还揭示了iEVT和DSC之间的相互作用(图4k)。此外,iEVT对自然杀伤 (NK) 细胞表现出强烈的向外信号,表明 iEVT在调节 NK 细胞方面发挥着作用。
此前,他们提出了一种假血管化模型,其中螺旋动脉的eEVT重塑涉及其对内皮细胞表型的模拟,并由其CDH5表达所支持。在所有空间转录组样本中(图2a),他们验证了该模型在全转录组范围内的有效性。随着EVT接近血管壁,它们从间质表型转变为血管内表型,PRG2表达下调, NCAM1表达上调。同时,内皮相关基因的表达在靠近血管壁的eEVT中达到峰值(图4l,左)。此外,基于在mVEC中富集程度最高的100个基因(来自snRNA-seq)(图1b),为每个EVT分配一个“内皮样”评分,该评分与EVT距血管壁的距离呈显著负相关(图4l,右),表明EVT在占据螺旋动脉微环境的过程中逐渐获得内皮细胞功能。因此,空间分辨数据支持了假血管化模型,表明eEVT系统性地获得了内皮样特性。此外,eEVT与其他EVT亚型共聚(图4d)表明,在内皮细胞模拟的同时,滋养层细胞特性也得以保留。
EVT表现出侵袭性的异质性,这促使他们检验单细胞转录组是否能够预测其侵袭潜能。本分析基于以下三点考虑。首先,通过聚焦于MFI,深层蜕膜EVT在空间上与浅层子宫肌层分离,从而减少了可能已停止侵袭的子宫肌层周围EVT的干扰。其次,大多数浅层蜕膜EVT(约77.3%)表达AOC1(图4i),AOC1是成熟EVT的标志物,这限制了发育异质性的干扰。第三,由于EVT转录组整合了细胞自主和微环境信号,基于转录组的推断能够捕捉到与侵袭相关的两种变异来源。
他们考虑了与血管无关的滋养层细胞(即eEVT和pEVT),并训练了一个学习模型来比较不同蜕膜深度下的滋养层细胞转录组。该模型从3192个滋养层细胞富集基因(图4e)中随机选择了54个,并将其表达聚合为侵袭性评分(iScore)。在随机保留的测试集(未用于训练的滋养层细胞)中,iScore与滋养层细胞侵袭深度密切相关(图4m),并且在iScore高的细胞中,上调基因富集于细胞迁移和细胞外基质(ECM)重塑程序,证实了其生物学相关性。在独立的单细胞数据集中,来自EVT(已知其侵袭性极低)确实表现出较低的iScore(图4n)。相反,来自胎盘植入谱系妊娠(以过度侵袭为特征)的EVT,无论取自蜕膜附着区还是非附着区,其iScore均较高(图4n)。这些独立的验证证实了生物学特异性。在空间数据中,高iScore的EVT通常位于深层蜕膜,而低iScore的EVT则位于表层(图4o,左)。异常值包括表层高iScore iEVT(持续侵袭)(图4o,中间)和深层低iScore iEVT(侵袭减弱)(图4o),表明iScore不仅反映了空间位置,还反映了侵袭状态的异质性。延伸至eEVT,它们在血管附近表现出较高的iScore(图4o)。根据snRNA-seq数据对EVT进行评分,祖细胞的iScore最低,而eEVT和pEVT的iScore最高。iEVT呈现出阶段依赖性模式:随着HLA-G+细胞进入蜕膜,iScore急剧上升,然后随着AOC1+表达和成熟而下降(图4p)。因此,iEVT 的侵袭潜力与发育成熟度呈负相关;这种拮抗作用表明子宫侵袭受到严格控制的机制基础。
snRNA-seq数据证实了先前描述的SCT祖细胞群(proSCT)(图4d),其以合胞素-2(ERVFRD-1)和SLC26A2为标志。紧随其后的是SCT-A亚型,其KLRD1(编码CD94)表达增强(图4q),以及以GPC5为标志的终末SCT-B状态(图4r)。CD94免疫染色将SCT-A定位于漂浮绒毛的合胞体,而RNAscope和空间转录组学分析显示GPC5在合胞体结节中富集(图4s-t)。与妊娠晚期合胞体结的积累一致,SCT-B 核在妊娠后期富集,验证了合胞体结中从 proSCT 到 SCT-A 最终到 SCT-B 的轨迹。
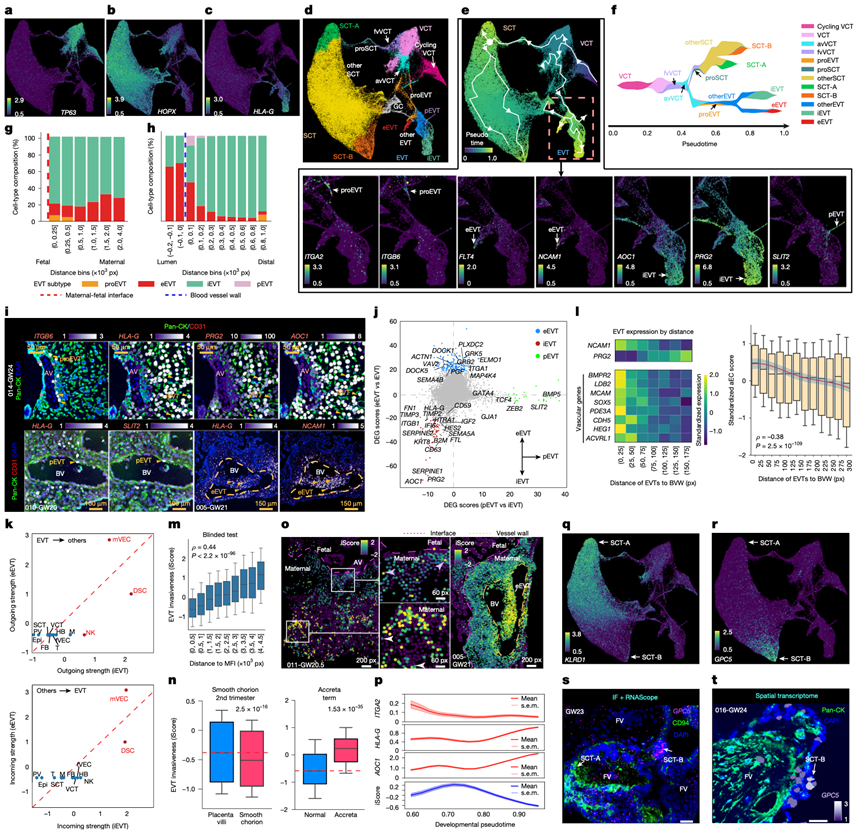
图4. 人类滋养层发育和功能的分子和空间特征。
(a-c) 滋养层亚型的UMAP图谱(基于标记表达)。(d) 滋养层亚型的聚类。(e) UMAP图谱重建的拟时间轨迹揭示了漂浮绒毛和锚定绒毛发育路径的差异(上图)。下图,UMAP图谱上EVT亚型特异性标记的表达。(f) VCT分化过程的流图。(g-h) 蜕膜中EVT亚型的比例,按距MFI或母体血管壁的距离进行分组。(i) 利用泛细胞角蛋白(pan-CK)和CD31免疫染色进行空间转录组学分析。(j) 差异表达分析(pEVT与iEVT以及eEVT与iEVT)鉴定出亚型特异性特征。(k) CellChat识别出不同的输出(上图)和输入(下图)相互作用、相互信号传导。(l) EVT 假血管化模型的验证。(m) 学习模型基于单细胞转录组预测 EVT 的侵袭性。(n) 左图:光滑绒毛膜 EVT 的侵袭性降低。右图:胎盘植入谱系中 EVT 的侵袭性增加。(o) 比较 EVT 在浅层和深层蜕膜隔室(左图和中图)以及血管附近(右图)的侵袭性。(p) 侵袭性(iScore;底部)以及沿重建轨迹的EVT成熟阶段ITGA2(祖细胞)、HLA-G(细胞柱出口)和AOC1(高级成熟)的表达。(q-r) 鉴定出一种新的SCT终末状态,其在SCT-A和SCT-B中KLRD1(编码CD94)和GPC5的表达存在差异。(s-t)免疫荧光分析。
05
新型DSC状态
对妊娠期20579个DSC进行snRNA-seq,鉴定出五个不同的细胞聚类(DSC0–DSC4)(图5a)。伪时序分析揭示了两条以DSC0为起点的进化轨迹:路径A(DSC0 → DSC1 → DSC3)和路径B(DSC0 → DSC2 → DSC4)(图5b)。DSC0在妊娠早期占主导地位,而DSC3和DSC4则出现较晚(图5c)。先前的早期妊娠图谱仅描述了路径A;因此,路径B及其末端DSC4细胞群代表了一个先前未识别的蜕膜谱系。值得注意的是,DSC4在非妊娠的人类子宫内膜(增殖期和分泌期)中也不存在,表明其出现具有妊娠特异性。
标记物分析支持了这些轨迹。DSC0 细胞表达高水平的ACTA2和低水平的IGFBP1和PRL,表明其处于未蜕膜化状态;而 DSC3 和 DSC4 细胞则表现出低水平的ACTA2表达和高水平的IGFBP1表达,表明其已发生蜕膜化(图5d)。SERPINF1 (编码 PEDF)沿路径 A 特异性上调(图5d-e),从而区分了 DSC3(PEDF+ IGFBP1+)和 DSC4(PEDF- IGFBP1+)。免疫定位显示,ACTA2+ DSC0 细胞位于蜕膜深处,而 DSC3(PEDF+IGFBP1+)细胞则广泛分布(图5f),常位于血管周围区域。他们还观察到少量CK +细胞滋养层细胞对PEDF呈免疫染色阳性(图5f)。相比之下,新型DSC4亚型(PEDF- IGFBP1+)定位于蜕膜浅层,许多细胞邻近锚定绒毛的末端(图5f),这一模式随后经空间转录组学证实。
接下来,他们对两条 DSC 发育通路进行了功能表征(图5e)。沿通路 A 上调的基因富集于内皮细胞迁移,这与 DSC3 频繁的血管周围定位以及其他多种功能相一致。在通路 A 的末端(DSC3),他们观察到其分叉为两个亚聚类,DSC3.1 和 DSC3.2(图5b),这两个亚簇均表达IGFBP1。主要在妊娠第 22 周后出现的 DSC3.2 细胞广泛下调血管生成抑制因子、MHC I 类分子、补体成分和抗凋亡基因。这些特征支持衰老样蜕膜状态,与之前的报道一致,即衰老的 DSC 在妊娠后期积累并促进分娩。
相比之下,沿路径 B 上调的基因富集于 ECM 组织(图5e)。DSC4 细胞特异性表达SEMA3A、WNT5A和CNR1(图5d)。SEMA3A和WNT5A促进 ECM 刚性和稳定性,从而抑制细胞运动。标记物CNR1编码大麻素受体 CB1。由于 DSC4 细胞位于表层蜕膜,通常靠近锚定绒毛的末端(图5f),而EVT在此处启动侵袭,因此 DSC4 标记物的表达表明 DSC4 介导人 MFI 处的内源性大麻素信号传导,并可能作为一种局部抑制机制来限制 EVT 的侵袭。
他们绘制了DSC状态的空间分布图,然后利用iScore指标(图4m)定量了DSC4邻近EVT的侵袭性,使用来自snRNA-seq的基因特征评分+,在空间分布图上从IGFBP1+ DSC中识别出DSC3和DSC4亚型,其空间定位与免疫染色结果一致(图5f)。ACTA2+VIM+未蜕膜化基质细胞和SERPINF+ IGFBP1+DSC3细胞(图5g)在深层蜕膜中富集。CNR1+ IGFBP1+ DSC4细胞通常位于锚定绒毛附近(图5g)。DSC3距离MFI最远,而DSC4距离MFI最近(图5h)。当映射到空间生态位时(图2c-d),DSC4 在富含 EVT 的 D1 生态位中富集(图5i),提示 EVT 与 DSC4 之间存在相互作用。为了验证这一结果,他们将 DSC4 邻近的 EVT 的 iScore 与以下两组进行比较:(1)与其他 DSC 亚型邻近的 EVT;以及(2)深度匹配的 EVT。结果显示,DSC4 邻近的 EVT 的 iScore 显著低于这两个比较组(图5j),表明 DSC4 介导了 EVT 侵袭性的局部抑制。
由于大麻素受体CB1(由CNR1编码)在DSC4细胞中特异性表达(图5d-g),且CB1可由内源性大麻素花生四烯酸乙醇胺(AEA)激活,而AEA在MFI处的平衡对妊娠至关重要,因此他们研究了CB1激动剂如何影响DSC4及其对滋养层细胞侵袭的调控。利用建立的体外蜕膜化模型,将人子宫成纤维细胞(HuF)分化为蜕膜化基质细胞,并暴露于0.5 μM甲酰胺(mAEA,一种AEA类似物)或溶剂对照72小时。对 33088 个细胞(13493 个对照组细胞,19595 个 mAEA 暴露组细胞)进行单细胞转录组分析(scRNA-seq),鉴定出未蜕膜化的ACTA2+细胞聚类和蜕膜化的IGFBP1+细胞聚类,以及根据原代组织中这些细胞类型的标记物定义的 DSC3 样和 DSC4 样亚型(图5k)。DSC4 样细胞保留了强烈的CNR1表达(图5k),表明该模型重现了 DSC3 和 DSC4 的分化过程。
鉴于 DSC3 中CNR1的缺失及其在 DSC4 中的特异性表达,他们在 mAEA 处理后进行了亚型特异性差异表达分析。基因集富集分析揭示了不同的反应(图5l)。在 DSC3 中,mAEA 倾向于上调促凋亡基因,这与先前关于 mAEA 诱导 DSC 细胞凋亡的报道一致。在 CB1⁺ DSC4细胞中,凋亡过程显著下调,包括促凋亡基因(DAPK3、FADD和GADD45G)的表达,表明 CB1 介导了 mAEA 处理后细胞凋亡的保护作用。
他们检测了内源性大麻素信号是否改变了DSC对原代人滋养层细胞侵袭的调控。将蜕膜化的DSC用mAEA(CB1激动剂)、利莫那班(SR141716A,CB1拮抗剂)、二者联合处理或溶剂对照处理72小时。洗涤后,收集条件培养基24小时,并将其应用于Transwell侵袭实验中妊娠中期的人滋养层细胞(图5m)。mAEA处理的DSC的条件培养基降低了滋养层细胞的侵袭能力,而利莫那班处理的DSC的条件培养基则增加了侵袭能力;联合处理使侵袭能力恢复至接近对照组水平(图5n)。这些数据共同表明,内源性大麻素暴露可保护 DSC4 免于凋亡(图5l),并调节 DSC4 旁分泌信号(图5n),从而增强其限制邻近 EVT 侵袭性的能力(图5o)。
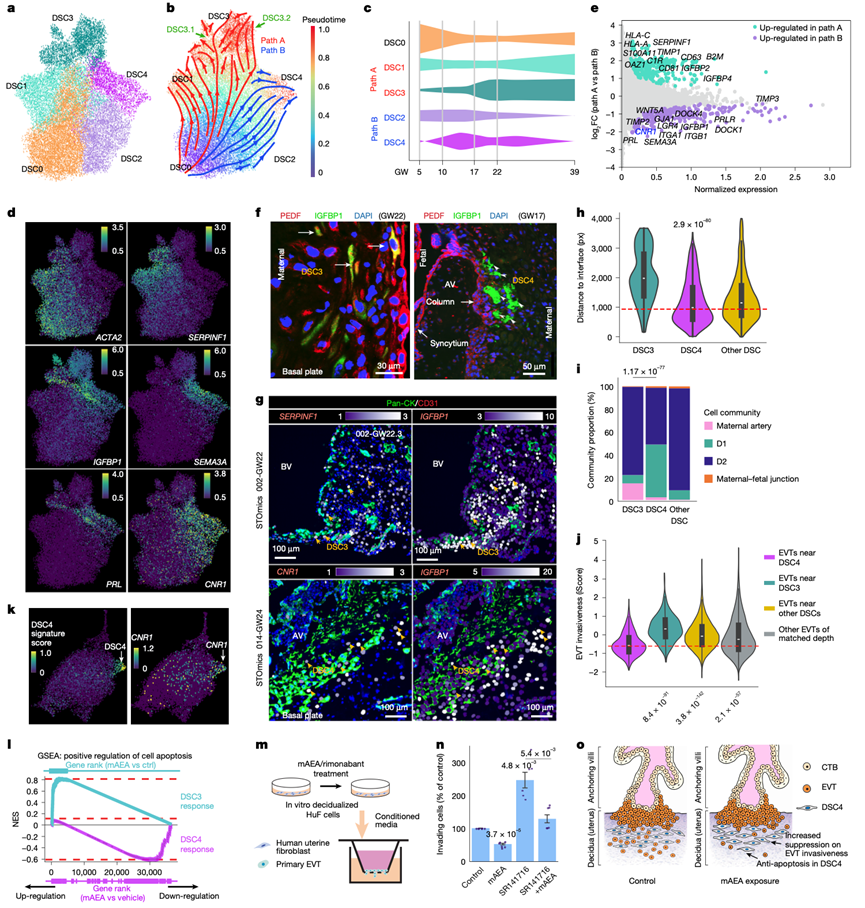
图5. DSC 异质性和功能的分子和空间表征。
(a) 基于snRNA-seq的五种蜕膜干细胞亚型(DSC0-DSC4)的UMAP投影。(b) 主要蜕膜化路径的重建轨迹:路径A(DSC0 → DSC1 → DSC3)和路径B(DSC0 → DSC2 → DSC4)。(c) 蜕膜干细胞亚型在妊娠期(GW5-39)的分布。(d) 蜕膜干细胞亚型特异性标记物在UMAP上的表达。(e) 差异表达分析表明,SERPINF1(编码PEDF)是区分路径A和路径B蜕膜干细胞的关键标记物。(f) DSC亚型的免疫定位。(g) 空间定位。(h) 空间距离分析。(i) 比较不同细胞群落中 DSC3 和 DSC4 的比例。(j) 与 DSC4 相邻的 EVT 的 iScore 低于 DSC3 附近的 EVT、其他 DSC 或处于相似蜕膜深度的 EVT(16 个样本)。(k) 体外蜕膜化的 HuF 细胞重现了 DSC4 的转录组特征。(l) 基因集富集分析(GSEA)。(m) Transwell侵袭实验示意图。(n) 原代细胞滋养层侵袭的定量分析。
06
疾病中最易感的细胞类型
接下来,他们研究了单细胞图谱是否能够解释主要妊娠并发症的遗传结构,使用 SCAVENGE将单细胞开放染色质结构与大规模(n > 10000)母胎全基因组关联研究 (GWAS) 数据整合,这些研究针对先兆子痫、自发性早产和散发性流产,从而生成了每种疾病的细胞类型解析的遗传风险图谱(图6a)。传统的先兆子痫 GWAS 样本量庞大,但仅发现了少数效应值较小的位点,导致大部分遗传力无法解释。相比之下,SCAVENGE 将每个细胞开放染色质区域的 GWAS 信号聚合为一个基于每个细胞的性状相关性评分。他们将母体细胞与母体先兆子痫 GWAS进行配对,并将胎儿细胞与胎儿先兆子痫 GWAS进行配对,结果鉴定出 6221 个母体细胞和 8232 个胎儿细胞具有显著的先兆子痫风险富集。在胎儿细胞中,只有 EVT(特别是 iEVT)富集了先兆子痫风险(图6b)。在母体细胞中,富集的细胞群包括 DSC3 亚型、动脉内皮细胞(非静脉内皮细胞)、血管周围细胞、成纤维细胞和 T 细胞,而蜕膜巨噬细胞和 NK 细胞则未富集(图6c)。一种独特的POU5F1+LGR5+子宫内膜上皮细胞群,仅限于妊娠早期,并表达干细胞标志物,也显示出显著富集,提示早期子宫内膜缺陷与先兆子痫相关。使用 DisGeNET收录的151个先兆子痫基因进行验证,证实了这些易感细胞类型中的表达富集。阴性对照实验将相同的GWAS与胎儿脑或成人脑单细胞数据集配对,未检测到显著富集,表明其特异性。需要注意的是,该分析侧重于群体遗传风险(常见变异),其他细胞类型可能通过罕见变异、体细胞变异或非遗传因素发挥作用。
接下来,他们将相同的策略应用于母体自发性早产和散发性流产的全基因组关联研究(GWAS),并对母体细胞类型进行配对分析。在这两种情况下,只有POU5F1+LGR5 +子宫内膜上皮细胞显示出显著富集(图6d-e),这一关联在包含233290名女性的独立自发性早产队列中得到了验证。这种上皮细胞群在先兆子痫、自发性早产和流产中的共同脆弱性支持了“子宫内膜谱系疾病”的概念。
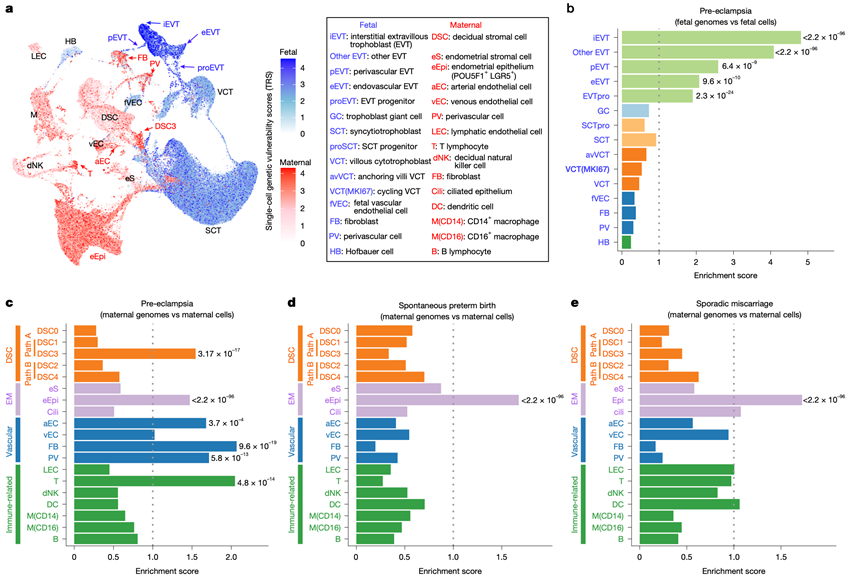
图6. 妊娠主要并发症的单细胞风险图。
(a) 通过将高风险变异映射到单细胞的开放染色质上,将单细胞多组学与母体和胎儿先兆子痫全基因组关联研究(GWAS)相结合。(b-c) 与先兆子痫显著相关的母体或胎儿细胞类型和亚型。(d-e) 针对自发性早产和散发性流产的母体全基因组关联研究(GWAS)风险。
+ + + + + + + + + + +
结 论
本研究通过整合多组学以及CODEX多重蛋白成像技术,构建了从妊娠早期到足月正常妊娠期间人类母胎界面的综合图谱,显著提升了先前研究的时空分辨率。该图谱描绘了胎儿和母体区域中常见的和瞬时的细胞类型、状态和空间微环境,重建了指导滋养层细胞和蜕膜基质细胞分化的转录调控机制,并解析了构成该界面的重复出现的结构单元。本研究鉴定出细胞滋养层介导的螺旋动脉重塑过程中先前未曾发现的动脉内皮状态转变,并开发了一种机器学习模型,该模型能够根据转录组特征预测细胞滋养层的侵袭性。进一步发现了一种蜕膜基质细胞亚型,该亚型通过内源性大麻素信号通路抑制人类母胎界面处的细胞滋养层侵袭。通过将该图谱与全基因组关联数据整合,本研究精确定位了最易受先兆子痫、早产或流产影响的母体和胎儿细胞。该研究提供了人类胎盘和蜕膜的全面、空间分辨的单细胞多组学参考,并为解码其正常和异常发育过程提供了一个框架。
+ + + + +



