

 English
English文献解读|Nature(58.39):全小鼠大脑中细胞类型的高分辨率转录组和空间图谱
✦ +
+
论文ID
原名:A high-resolution transcriptomic and spatial atlas of cell types in the whole mouse brain
译名:全小鼠大脑中细胞类型的高分辨率转录组和空间图谱
期刊:Nature
影响因子:58.39
发表时间:2023.12.13
DOI号:10.1038/s41586-023-06812-z
背 景
哺乳动物的大脑极其复杂,控制着机体的多种活动,包括活力、体内平衡、睡眠、意识、感觉、先天行为、目标导向行为、情感、学习、记忆、推理和认知。这些活动由高度专业化且错综复杂的集成神经回路控制,该神经回路由具有不同分子、解剖和生理特性的多种细胞类型组成,因此有必要全面了解构成大脑分子和解剖结构的细胞类型和回路。
实验设计
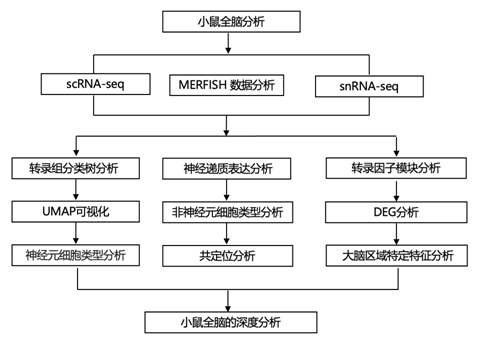
结 果
01

小鼠脑细胞类型图谱的创建
研究者团队利用单细胞转录组分析(scRNA-seq)和单细胞核转录组分析(snRNA-seq)数据建立了完整的小鼠大脑的高分辨率转录组和空间细胞类型图谱,具有四个嵌套的分类级别:34个类、338个亚类、1201个超类型和5322个聚类或类型(图1)。他们将这些类别分为七个邻域,以便对相关细胞类型子集进行更深入的分析。他们提供了该图谱的几种表示形式以供进一步分析:(1)亚类分辨率的树状图以及显示各种元数据信息的条形图(图1a);(2)单细胞分辨率下的均匀流形近似和投影(UMAP),用不同类型的元数据信息着色(图1b-e);(3) 亚类分辨率的星座图,描绘不同亚类之间的多维关系。

图1. 整个小鼠大脑的转录组细胞类型分类。
(a)以树状图组织的 338 个亚类的转录组分类树。(b-e) 所有细胞类型的 UMAP 表示,分别按类别、亚类、大脑区域和主要神经递质类型着色。
02
小鼠大脑中的神经元细胞类型
神经元细胞类型在全脑细胞类型图谱中占很大比例,包括 6 个邻域、29 个类(85%)、315 个亚类(93%)、1156 个超类(96%)和 5205 个聚类(98%)。神经元类型具有高度的区域特异性,并表现出高度可变的相似性和差异性。为了进一步研究每个主要大脑结构内的神经元多样性,他们为上述神经元类型的邻域生成了重新嵌入的 UMAP(2D 和 3D),以揭示大脑区域内和之间的神经元类型之间的细粒度关系MERFISH 数据(图2),并发现了不同神经元亚类之间的转录组特异性和相关性以及空间特异性和相关性之间的显著对应关系。
来自所有大脑皮层结构[包括同皮质、海马形成(HPF)区 、嗅觉区(OLF)和皮质底板(CTXsp)]的谷氨酸能神经元形成一个独特的大脑皮层-谷氨酸邻域,其中包括亚类1-38 和总共 517 个聚类(图2a-b),每个神经元亚类都表现出层和/或区域特异性(图2a-b)。
一组发育中的大脑皮层下衍生的 GABA 能(产生 γ-氨基丁酸)神经元亚类,包括在大脑皮层结构和大脑皮层下的小脑核(CNU) 中发现的所有 GABA 能神经元,包括背侧 STR (STRd) 和腹侧 STR (STRv)、外侧间隔复合体(LSX)、背侧 PAL (PALd)、腹侧 PAL (PALv) 和内侧 PAL (PALm),形成皮质下-GABA 邻域(图2c-d)。根据每个亚类的分子特征和区域特异性,亚类-GABA 邻域(亚类 39-90,总共 1051 个聚类)共分为 7 个类,这些类可能与其不同的发育起源相关(图2c-d)。
HY-EA-Glut-GABA邻域(包括66亚类和73-144亚类,共1404个聚类)包含一组来自整个下丘脑的密切相关的神经元亚类,以及CNU的纹状体样杏仁核(sAMY)和尾部螺旋体(PAL)区域,也称为延伸杏仁核(图2e-f),这一区域的谷氨酸能和γ -氨基丁酸能神经元亚类表现出逐渐的由前向后的转变,因此分为6个亚类(图2e-f)。下丘脑最前部的POA神经元类型与sAMY和PAL的神经元类型高度相似。因此,CNU-HYa GABA类也包括在上述的丘脑下-GABA邻区中,以显示它们与纹状体/苍白质类型的相关性和连续性(图2c-d)。
第四个神经元邻区TH-EPI-Glut(145-154亚类,共148个聚类)包含位于丘脑的所有谷氨酸能神经元亚类,以及内侧束(MH)和外侧束(LH),它们共同组成上皮体(EPI)(图2g-h)。
第五个神经元邻域,中脑(MB)-后脑(HB)-谷氨酸-血清素-多巴胺,包含MB和HB中所有的谷氨酸能、血清素能和多巴胺能神经元类型(图2i-j)。
第六个也是最后一个神经元邻域,MB-HB-CB-GABA,包含位于中脑、后脑和小脑的所有 GABA 能亚类(图2k-l)。
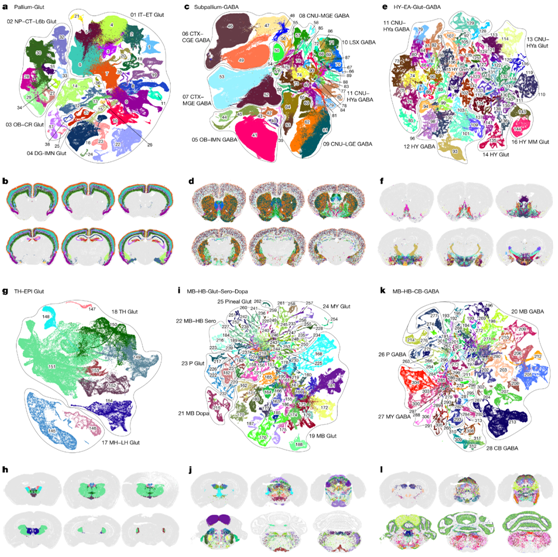
图2. 神经元细胞类型分类和整个大脑的分布。
(a,c,e,g,i,k) UMAP 可视化。(b,d,f,h,j,l) Pallium-Glut的代表性MERFISH切片。
03
神经递质和神经肽表达
他们根据典型神经递质转运蛋白基因和合成酶的共表达,并考虑替代神经递质释放机制,系统地将神经递质身份分配给每个细胞聚类(图3a-c)。这些标记基因表明大多数神经元聚类释放单一神经递质——谷氨酸或 GABA。中脑和后脑中的许多 GABA 能神经元聚类共同释放甘氨酸。他们鉴定了62个具有谷氨酸-GABA双重递质(Glut-GABA)的聚类,其中大多数表达谷氨酸转运蛋白基因Slc17a6或Slc17a8。这些聚类广泛分布在大脑的不同部位。它们包括等皮质和海马中的四个聚类以及苍白球内部节段(GPi)中的三个聚类,这可能对应于这些区域中先前充分表征的谷氨酸-GABA 共同释放神经元类型。
他们还系统地鉴定了产生调节性神经递质的所有聚类(图3a-c)。胆碱能神经元主要分布于PAL腹侧的58亚类(11个聚类),但也包括LSX的2个聚类、MH的8个聚类、PPN的3个聚类、迷走神经背侧运动核(DMX)和孤立束核(NTS)的5个聚类,以及大约13个聚类散在分布于其他髓质核。胆碱能神经元常同时释放谷氨酸(24个聚类),有时释放γ -氨基丁酸(7个聚类),同时释放谷氨酸和γ -氨基丁酸(3个聚类),或释放多巴胺(DMX中的1个聚类)。
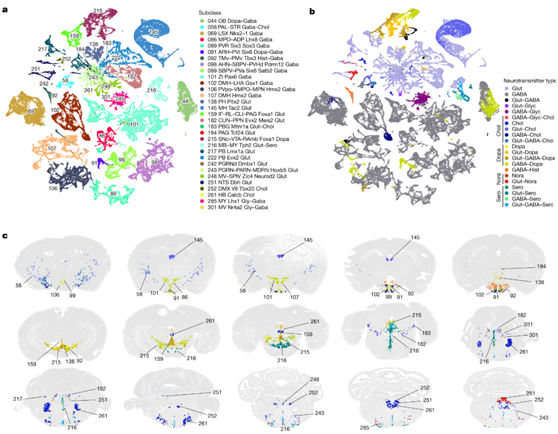
图3. 调节性神经递质类型及其在整个大脑中的分布。
(a-b) 神经元亚类,包含释放调节性神经递质及其与谷氨酸和/或 GABA 的各种共释放组合的聚类。UMAP 按亚类和神经递质类型进行着色。 (c) 代表性MERFISH切片显示表达调节性神经递质的神经元类型的位置。
04
非神经元和未成熟的神经元细胞类型
与上面定义的六个神经元邻域不同,第七个也是最后一个邻域NN-IMN-GC数据包含高度不同的非神经元细胞类型、未成熟神经元类型和颗粒细胞类型的混合集合(图4a)。它有9个类,包括5个非神经元类(Astro-Epen、OPC-Oligo、OEC、Vascular and Immune)和4个颗粒和未成熟神经元类(DG-IMN Glut、OB-IMN GABA、松果体Glut和CB Glut)。鼠脑中的所有非神经元细胞类型可分为5类,23个亚类,45个超聚类和117个聚类(图4a),可通过各层级高度特异性的标记基因进行区分。Astro-Epen类是最复杂的,包含10个亚类,其中5个亚类代表不同脑区的星形胶质细胞:Astro-OLF、Astro-TE(端脑),Astro-NT(非端脑),Astro-CB和Bergmann胶质,而其他5个亚类代表室管膜细胞类型:星形室管膜细胞、室管膜细胞、扩张细胞、室管膜细胞和脉络丛(choroid plexus, CHOR)细胞(图4a-c)。OPC - Oligo类包含两个亚类,少突胶质细胞前体细胞(OPC)和少突胶质细胞(Oligo)。
在所有的非神经元细胞类型中,Astro-Epen类表现出最多样化的空间模式。区域特异性星形胶质细胞Astro-OLF、Astro-TE、Astro-NT和Astro-CB在UMAP中按照从前到后的顺序排列(图4b),与它们的空间模式一致。室管膜的五个亚类——室管膜、室管膜、伸长细胞、室管膜和CHOR——分布在整个大脑脑室的不同部位,其中的聚类表现出精细的空间特异性(图4c)。脑室周围器官(CVO)是位于第三、第四脑室周围的特殊结构,介导脑、血液和脑脊液(CSF)之间的通信。它们是高度血管化的,内衬室管膜细胞和伸长细胞,作为脑与血液和/或脑脊液之间的选择性屏障。伸长细胞是一种特化的室管膜细胞,分布在下丘脑的第三脑室(V3)和正中隆起(ME)。他们确定了与每种亚型相对应的聚类:聚类5245/5246、5247、5249和5250分别为α1、α2、β1和β2(图4e)。他们还发现了长管细胞样室管膜细胞聚类,这些细胞聚类特异性位于其他CVO(图4c):聚类5243位于穹基下器官(SFO), 聚类5244位于终板血管器官[OV(也称为OVLT)], 聚类5240位于最后区(AP),以及聚类5263(Sspo标记)位于亚联合器官(SCO)。
VLMC类型还表现出高度特异的空间和共定位模式。聚类5296-5299位于软脑膜,而聚类5300和5301则广泛分布在大脑中(图4d)。值得注意的是,他们发现在VLMC聚类5303和脑室膜细胞聚类之间(图4e)、在VLMC聚类5302和室管膜和CHOR聚类之间(图4f)、以及在软脑(脊)膜特异性VLMC聚类和ILA之间存在高度特异的空间共定位。
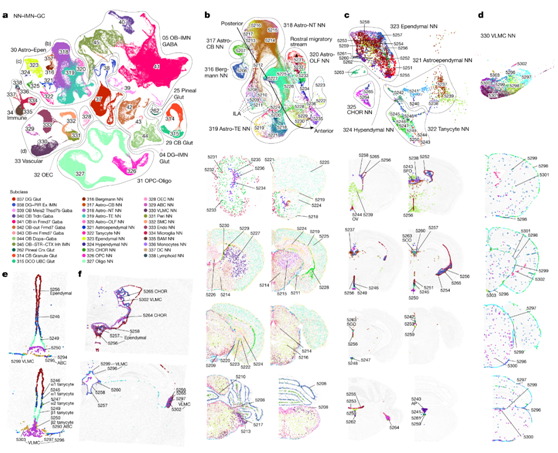
图4. 非神经元细胞类型和未成熟神经元类型。
(a) 按亚类着色的 NN-IMN-GC 邻域的 UMAP 表示。(b-d) 星形胶质细胞、室管膜细胞和 VLMC的亚群分析。 (e) V3 和 ME 周围单细胞、室管膜细胞和 VLMC 聚类的共定位。 (f) VLMC、CHOR 和室管膜细胞聚类在各个心室中的共定位。
05
定义细胞类型的转录因子
转录因子是细胞类型身份的关键调节因子。为了评估转录因子表达与转录组细胞类型的对应关系,他们计算了每对神经元与非神经元类、类、亚类或亚类内的聚类对之间差异表达的转录因子的数量(图5a )。然后,使用基于所有 8460 个差异表达基因(DEG)、534 个转录因子标记基因、541 个功能基因、编码粘附分子的基因和 534 个随机选择的 DEG 构建的分类器,比较了亚类和聚类召回的交叉验证准确性(图5b)。与功能基因、粘附分子和所有DEG相比,转录因子标记基因在所有聚类中显示出最低的基因表达相关性(图5c),这表明转录因子具有最大的区分细胞类型的能力。这些结果量化了转录因子在定义细胞类型身份方面的主要作用。
他们鉴定了一大类转录因子共表达模块(52 个模块),它们在所有层级的特定细胞类型组中选择性表达,因此可以定义这些细胞类型组的身份(图5d)。皮层谷氨酸能特异性模块包括Tbr1和Satb2,它们在不同亚类中也表现出差异表达。直接早期基因Egr3和Nr4a1在大脑皮层谷氨酸能神经元中高表达,而Fos和Fosb表达更为均匀。
虽然许多转录因子同源物是共表达的(图5d),但它们也可以表现出不同的表达模式。他们检测了几个转录因子家族的表达模式,包括叉头框(Fox), Krüppel-like因子(Klf), LIM同源盒(Lhx),Nkx同源盒(Nkx),核受体(Nr),配对盒(Pax), POU结构域(POU),正调节结构域(Prdm),SRY相关的HMG-box (Sox)和T-box (Tbx),所有这些转录因子家族存在于发育过程中的空间模式,细胞类型的分化和分化中发挥重要作用。
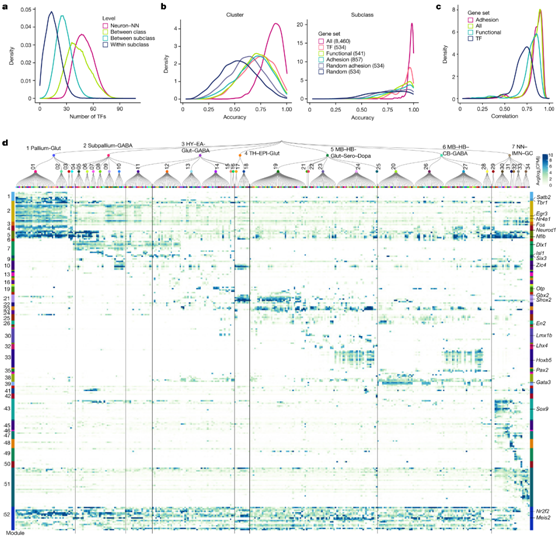
图5. 整个小鼠大脑的转录因子模块。
(a) 神经元和非神经元类之间、类之间、亚类之间和亚类内差异表达转录因子(TF)数量的分布。 (b) 使用基于所有 8460 个标记基因(全部)、534 个转录因子标记基因、541 个功能标记基因、857 个编码粘附分子的标记基因构建的分类器。 (c) 密度图,显示使用所有标记、粘附标记基因、功能基因和转录因子的聚类之间标记基因表达相关性的分布。 (d) 分类树中每个亚类的关键转录因子的表达,组织在转录因子共表达模块中,在热图两侧显示为彩条。
06
大脑区域特定的细胞类型特征
为了比较所有细胞类型的全局空间分布模式以及转录组相似性和空间邻近性之间的关系,他们使用全脑MERFISH数据集,在所有中间本体水平的脑区中量化了所有细胞亚类的全脑空间分布模式。结果表明,所有神经元亚类都局限于特定的脑区,而非神经元亚类则分布更广泛。在转录组学上,更相似的细胞类型在空间上彼此位于更近的位置——例如,同一类中的神经元亚类大多位于相同的主要脑区。相反,在转录组学上,距离越远的细胞类型在空间上彼此距离越远。每个主要的大脑区域都有自己特定的谷氨酸能和氨基丁酸能神经元亚类,这些亚类大多是局部定位的。他们进一步使用Gini系数和Shannon多样性指数来衡量各亚类空间分布的变异程度,两者均显示出各神经元亚类空间分布的高度不平等(即高度局域化模式)(图6a)。
他们发现,来自不同区域的聚类的数量与scRNA-seq所描绘的细胞数量没有相关性(图6b-c),相反,局部特征占主导地位。下丘脑、中脑和后脑区域包含最多的细胞聚类,表明细胞类型的高度复杂性,与这些广泛的区域具有许多小而异质性的亚区一致。相比之下,尽管由于其包含的许多亚区(包括异皮质、HPF、OLF和CTXsp,每个都包含多个亚区),其总体体积比其他主要脑结构大4到15倍,他们在苍白质中发现了更多的细胞。
他们计算了大脑区域内每对集群之间的DEG数量,根据相似性(即相似性越高,DEG数量越少)将这些数字分为9个分位数,并按分位数绘制了它们的分布(图6d)。在聚类数量较多的区域(即下丘脑、中脑和后脑),聚类在每个区域内彼此之间更加相似,这表明这些区域的细胞类型具有较低水平的转录组差异和较低的等级。相比之下,在聚类数较少的区域(即小脑、丘脑和苍白脑),聚类之间的DEG数量差异很大;因此,这些区域的细胞类型似乎更加多样化和分层,CNU表现出中等水平的多样性。他们基于MERFISH数据集计算了每个聚类的三维空间跨度,并汇总了每个脑区域内所有聚类的跨度(图6e)。同样,不同的区域表现出不同的特征,苍白球、CNU和小脑的聚类跨越更大,表明分布更广泛,而下丘脑、中脑和后脑的聚类跨越更小,表明定位更局限。
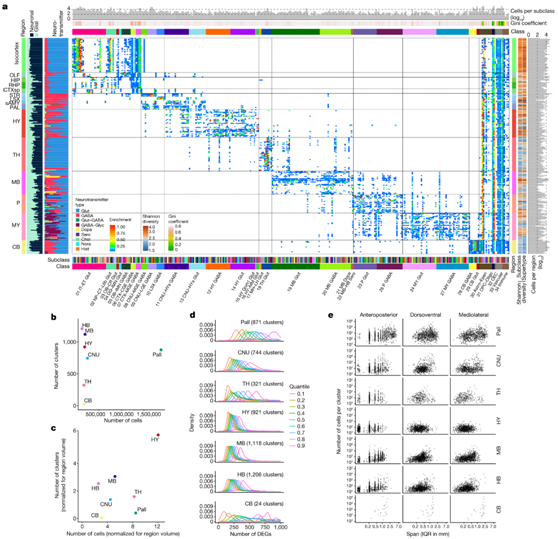
图6. 细胞类型的大脑区域特定特征。
(a) 热图显示 MERFISH 细胞的每个亚类(x轴)中的CCFv3 区域分布(y轴)。 (b) 散点图显示每个主要大脑区域识别的神经元聚类数量与相应区域中 scRNA-seq 分析的神经元细胞数量。(c) 与(b)相同,不同之处在于聚类和轮廓细胞的数量按区域体积标准化。(d) 每个主要大脑区域内每对神经元聚类之间的 DEG 数量的分布。 (e) 散点图显示映射到给定神经元聚类的细胞数量与基于 MERFISH 数据集的前后轴、背腹轴和中外侧轴的 3D 坐标跨度的关系,按主要大脑区域分层。
+ + + + + + + + + + +
结 论
本项研究报告了整个成年小鼠大脑的全面且高分辨率的转录组和空间细胞类型图谱。该细胞类型图谱是通过使用多重分析技术将约 700 万个细胞的scRNA-seq数据集与约 430 万个细胞的空间转录组数据集相结合而创建,系统地分析了大脑中的神经元和非神经元细胞类型,并确定了每种细胞类型的转录组身份和空间特异性之间的高度对应性。结果揭示了不同大脑区域细胞类型组织的独特特征,特别是大脑背侧和腹侧部分之间的二分性。背侧部分包含相对较少但高度分化的神经元类型,而腹侧部分包含更多数量的神经元类型,彼此关系更密切。这项研究还揭示了不同细胞类型中神经递质和神经肽表达以及共表达模式的多样性和异质性。最后,他们发现转录因子是细胞类型分类的主要决定因素,并确定了定义大脑所有部分细胞类型的组合转录因子代码。全小鼠大脑转录组和空间细胞类型图谱为哺乳动物大脑的细胞、发育和进化的综合研究建立了参考图谱和理论依据。
+ + + + +



