

 English
English文献解读|Nat Commun(14.7):衰老小鼠蛋白质组图谱揭示了与年龄相关的转录后失调的特征
✦ +
+
论文ID
原名:An atlas of the aging mouse proteome reveals the features of age-related post-transcriptional dysregulation
译名:衰老小鼠蛋白质组图谱揭示了与年龄相关的转录后失调的特征
期刊:Nature Communications
影响因子:14.7
发表时间:2024.10.02
DOI号:10.1038/s41467-024-52845-x
背 景
衰老是大多数慢性疾病的最大风险因素,了解其潜在机制对于开发与年龄相关的疾病的预防和治疗方法至关重要。
实验设计
结 果
01

小鼠衰老蛋白质组图谱的构建
研究团队对成年、中年、老年和老年小鼠(分别为 6、15、24 和 30 月龄)的主要组织的全组织裂解物 (WTL) 和低溶解度蛋白质富集级分 (LSF) 进行定量蛋白质组学分析。总共检测并定量了 9283 种蛋白质,WTL 和 LSF 中平均分别有 4608 种和 2906 种蛋白质(图 1a)。总体而言,同一组织的WTL和LSF的蛋白质组学图谱与不同组织的蛋白质组学图谱一样不同(图 1b)。此外,质谱检测显示,位于核心基质体和线粒体膜的蛋白质,根据MatrisomeDB和MitoCarta3.0分析结果显示,在LSF中所占的蛋白质、肽和信号的比例高于WTL(图1c),表明低溶解度蛋白质富集的成功和分析全面性的提高。
与 6 月龄小鼠相比,差异表达蛋白 (DEP)数量随着年龄的增长而逐渐增加(图1d)。但值得注意的是,C57BL/6 J 小鼠的存活率在 24 月龄左右开始下降,疾病对差异蛋白质表达的影响在 30 月龄时可能明显高于 24 月龄时。虽然大多数年龄依赖性蛋白质的表达都表现出随着年龄增长而发生的变化,但是在皮肤中,相当一部分年龄依赖性蛋白质在 15 月龄小鼠中表现出瞬时表达变化(图 1e)。与 6 月龄小鼠相比,30 月龄小鼠中大多数显著差异表达的蛋白质表现出逐渐的年龄相关变化(图 1f),表明这些变化与自然衰老过程有关。细胞衰老标志物p16和 DNA 损伤标志物 γ-H2AX 的表达水平在 30 月龄小鼠也高于 24 月龄小鼠。
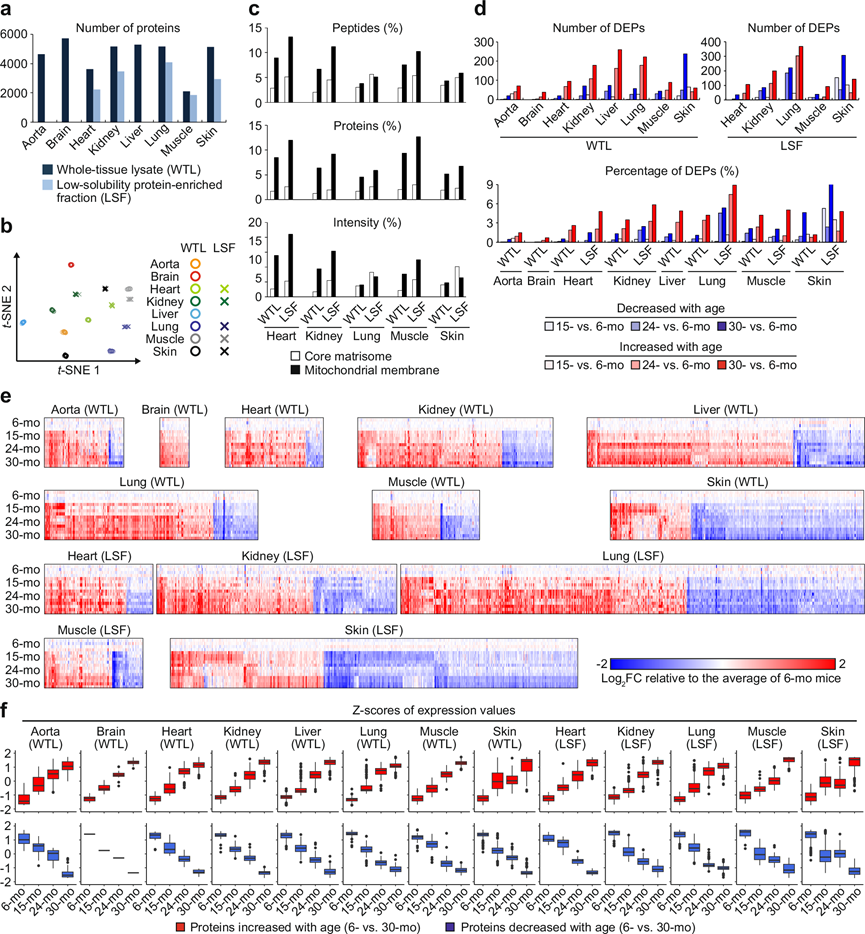
图1. 鉴定小鼠主要器官的全组织裂解物(WTL)和低溶解度蛋白质富集级分(LSF)中随年龄变化而差异表达的蛋白质。
(a) 蛋白质组学质谱分析检测和定量的蛋白质数量。(b) 使用 Seurat 流程构建的主要器官 WTL 和 LSF 蛋白质组学谱的 tSNE 图。(c) 核心基质体和线粒体膜蛋白的肽在 WTL 和 LSF 中测量的总肽中的百分比。(d) 小鼠主要器官 WTL 和 LSF 中随年龄差异表达的蛋白质数量。(e) 热图显示了 6 个月大和 15 个月大、24 个月大或 30 个月大小鼠之间差异表达蛋白质的相对蛋白质表达水平。(f) 箱线图显示了 6 个月大和 30 个月大小鼠之间差异表达蛋白质的表达水平的 Z 分数。
02
衰老过程中蛋白质组的组织和部分依赖性变化
在 30 个月大时,WTL 中的 DEP 百分比在肾脏、肝脏、肺、肌肉和皮肤中为 5–6%,在主动脉和心脏中为 2–3%,在大脑中为 0.7%。大脑中 DEP 数量较少可能是由于大脑组织的复杂性,也可能表明大脑对基因表达失调的抵抗力较低,在衰老过程中必须更好地保存。LSF 中的 DEP 百分比较高,在心脏、肾脏和肌肉中为 6-8%,在肺和皮肤中为 14-15%(图 1d)。对 LSF 的分析使 DEP 的数量翻了一倍(图 2a-b)。在 LSF 中发现的大多数 DEP 也在 WTL 中检测到,但在 WTL 中没有随年龄变化而差异表达,这表明这些蛋白质仅在特定细胞区室中受年龄影响或与其他降低其溶解度的条件相关时才受影响(图 2a)。在每种组织中检测到的蛋白质有明显的重叠(图 2c),但绝大多数 DEP 是组织依赖性的(图 2d)。
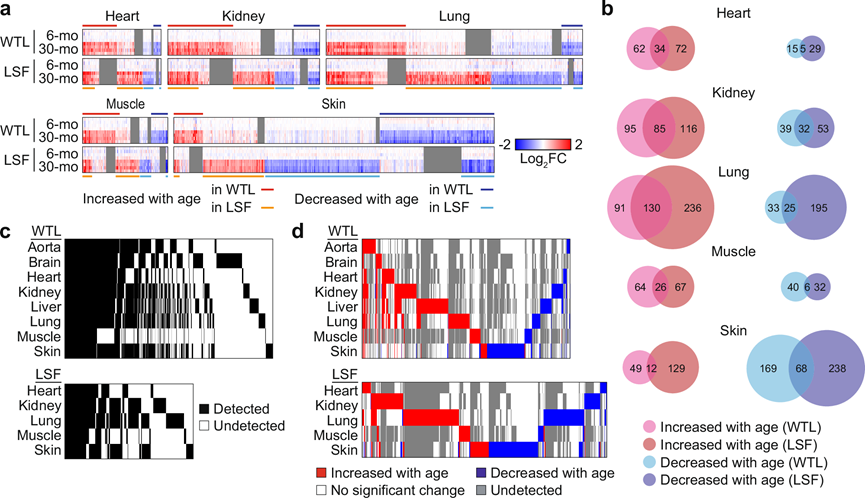
图2. 衰老过程中蛋白质组的组织和部分依赖性变化。
(a) 热图显示了在 WTL 和 LSF 中 6 个月大和 30 个月大小鼠之间差异表达的蛋白质的相对蛋白质表达水平。(b) 维恩图显示了在WTL和 LSF 中 6 个月大和 30 个月大小鼠之间差异表达的蛋白质之间的重叠。(c) 热图显示了在不同组织中检测到的蛋白质之间的重叠。(d) 热图显示了在不同组织中随年龄(6 个月 vs. 30 个月)差异表达的蛋白质之间的重叠。
然而,在每种组织的 DEP 中富集的通路彼此广泛重叠(图 3a-b)。与免疫反应相关的蛋白质在 WTL 和 LSF 中的所有组织中都随着年龄的增长而优先增加。随着年龄增长而增加的蛋白质,特别是在 LSF 中,也显示出免疫反应蛋白的富集。在 WTL 和 LSF 中,主动脉、心脏、肾脏、肝脏、肺和肌肉中的细胞外基质 (ECM) 蛋白随着年龄的增长而增加(图 3a-b)。从matrisome DB中检索到的核心基质体蛋白也富集在所有组织中随年龄增长的蛋白中,无论样本类型如何,并且在13个数据集中,有12个数据集的富集具有统计学意义(图3c)。在 LSF 中,心脏、肾脏和肺中线粒体或线粒体内膜复合物中的蛋白质随年龄不成比例地减少,但在 WTL 中没有(图 3b)。MitoCarta3.0 分析发现,心脏和肾脏中线粒体富集了随年龄减少的蛋白质(图 3d)。重要的是,虽然核心基质体和线粒体膜蛋白在多种组织中富含 DEP,但大多数 DEP 都是组织依赖性的,这表明这些区室在多种组织中随着衰老而发生显著变化,但变化方式高度依赖于组织。与免疫反应相关的 DEP 在组织中更常见,但其中相当一部分在肾脏、肝脏、肺和皮肤中是组织特异性的。
在主动脉中,随年龄增长而减少的蛋白质包括补体活化相关蛋白质。在肌肉中,随年龄增长而增加的蛋白质包括内质网 (ER) 和 ER 伴侣复合物中的蛋白质(图 3a)。以免疫反应相关基因为例,一部分蛋白质在不同组织中普遍受到年龄的影响。对于每个 WTL 和 LSF,他们将在中老年小鼠组织中显著且一致地增加或减少的蛋白质定义为“常见 DEP”(图 3e)。在 WTL 和 LSF 中,随年龄增长而增加的常见 DEP 参与 “抗原加工和呈递”和“补体和凝血级联”相关的通路。
他们在没有事先灌注或采血的情况下收获了小鼠组织样本,因此与年龄相关的血流或组织中免疫细胞的变化可能是免疫相关的常见 DEP 来源。主要在非免疫细胞中表达增加的常见DEP包括 SERPINE2 和 COL18A1(图 3e)。已知 SERPINE2 和 COL18A1 的 C 端片段具有抗血管生成作用,据报道,与年龄相关的 SERPINE2 水平增加与老年小鼠心脏血管生成受损有关。随着年龄增长而减少的常见 DEP 数量相对较少,包括肌酸激酶 (CKM)、小白蛋白 (PVALB)、主要尿蛋白(MUP3 和 MUP8)、核糖体蛋白 S17 (RPS17)、连接组蛋白 H1.5 (H1-5)、线粒体裂变过程 1 (MTFP1) 和 COX6C(图 3e)。
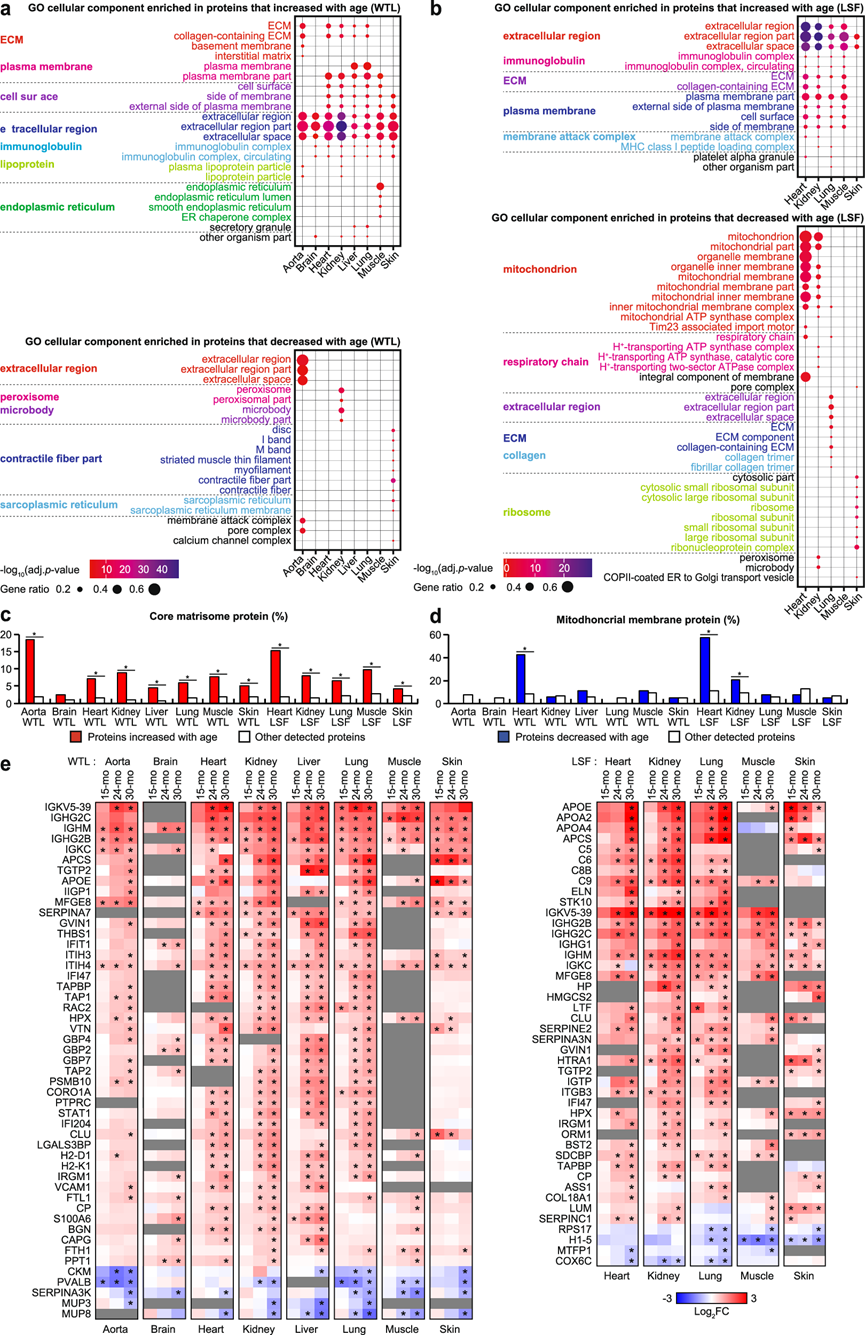
图3. DEP 的功能特性。(a-b)GO分析。
(c) 核心基质体蛋白在随年龄增加的蛋白质和其他检测到的蛋白质中的比例。(d) 线粒体膜蛋白在随年龄减少的蛋白质和其他检测到的蛋白质中的比例。(e) 热图显示了常见 DEP 蛋白质水平与年龄相关的变化。
03
性别和遗传背景的影响
接下来,他们对6个月和30个月大的雌性C57BL/6J小鼠(采用8重 TMT标记)的心脏和肾脏WTL以及4个月和32个月大的CB6F1雄性小鼠和4个月和24个月大的BALB/c雌性小鼠(采用10重TMT标记)的肾脏WTL进行了蛋白质组学分析(图 4a)。在任一性别或两种性别中鉴定的 DEP 蛋白质水平的与年龄相关的表达变化在雄性和雌性 C57BL/6 J 小鼠之间显示出良好的相关性。C57BL/6 J、CB6F1 和 BALB/c 小鼠的整体年龄相关蛋白质组学变化也相似(图4a)。蛋白质中核心基质体蛋白的富集随年龄增长而增加,这是雄性 C57BL/6 J 小鼠衰老蛋白质组的一个突出的组织独立特征(图 3c),但是在雌性 BALB/c 小鼠肾脏中没有统计学意义(图 4b)。值得注意的是,虽然不同条件之间与年龄相关的变化总体上存在相关性,但相当多的蛋白质仅在特定条件下随年龄显著差异表达。 例如,雄性 C57BL/6 小鼠肾脏的 DEP 随年龄增长而减少,而雌性 C57BL/6 小鼠肾脏中却没有富集微体和过氧化物酶体相关DEP。 另一方面,雌性 C57BL/6 小鼠肾脏的 DEP 随年龄增长而减少,而雄性 C57BL/6 小鼠肾脏中却没有富集基底外侧质膜和血影蛋白相关的细胞骨架相关蛋白。
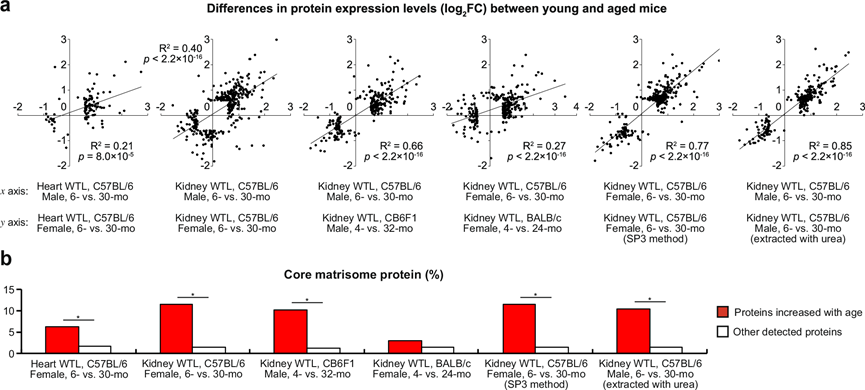
图4. 对研究结果普遍性的评估。
(a) 蛋白质表达水平差异分析。(b) 核心基质体蛋白在随年龄增加的蛋白质和其他检测到的蛋白质的比例。
04
与年龄相关的转录后失调的特征
结合对 6 月龄和 30 月龄小鼠组织的转录组学和蛋白质组学数据,他们发现只有大约 60% 的差异表达的 mRNA(DER)和 DEP 伴随其相应蛋白质和 mRNA 水平呈现显著一致变化(图 5a-b)。肾脏和肝脏中随着衰老而增加的蛋白质是例外,其中 69-80% 在 mRNA 水平上显著上调(图 5b)。下调基因中 mRNA 和蛋白质水平与年龄相关的变化之间的一致性低于上调基因。鉴于相当一部分 DEP 可能受到转录后机制的影响,他们接下来试图表征未伴随 mRNA 水平显著变化的 DEP 的特征。结果发现,在12个数据集中,仅在蛋白质水平表达显著增加的基因,其核心基质体基因比例高于在mRNA和蛋白质水平表达均增加的基因(图 5c)。另一方面,在10个数据集中,仅在蛋白质水平上显著降低表达的基因中,其线粒体膜基因的比例高于那些在mRNA和蛋白质水平上均降低表达的基因(图 5c)。非膜线粒体基因仅在蛋白质水平上随年龄减少(图 5c)。在大多数数据集中,仅在蛋白质水平上随年龄增长而减少的线粒体膜蛋白都富含OXPHOS蛋白,在肾脏和肺的LSF中具有统计学意义(图 5d)。
在 30 月龄小鼠中的DEP,如果未伴随 mRNA 水平的显著一致变化,则在 24 月龄小鼠中不太可能出现显著差异表达(图 5e)。这一结果表明,转录后机制对与年龄相关的蛋白质丰度变化的总体贡献通常在老年后变得更高。
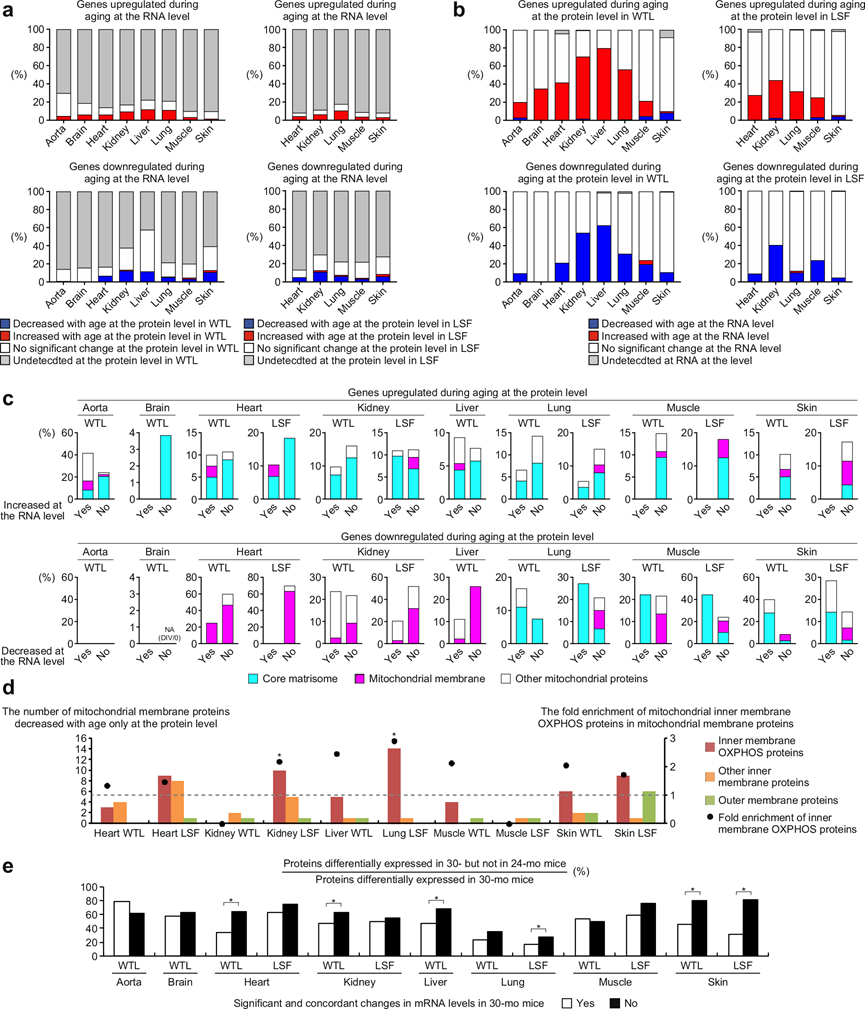
图5. 与年龄相关的转录后失调的特征。
(a)显著年龄相关变化的差异表达 mRNA (DER) 的比例。(b) 显著年龄相关变化的 DEP 的比例。(c) 核心基质体、线粒体膜和其他线粒体蛋白在 6 个月大和 30 个月大的小鼠之间差异表达的蛋白质总数中的比例。(d) 条形图显示随年龄增长而减少的线粒体膜蛋白数量。(e) 条形图显示 24 个月大小鼠中未出现差异表达的蛋白质在 30 个月大小鼠中出现差异表达的蛋白质中的比例。
为了创建一个主要通过转录后机制在高龄时失调的基因目录,他们列出了蛋白质产物随年龄显著增加但相应 mRNA 水平增加不超过 1.05 倍的基因,以及蛋白质产物随年龄显著降低但相应 mRNA 水平降低不超过 1.05 倍的基因(图6)。在 WTL 和 LSF 中发现的 DEP 分别有 198 个和 502 个符合这些标准。总共有 635 种这样的蛋白质,包括 41 种核心基质蛋白和 36 种线粒体膜蛋白。铁蛋白重链 (FTH1) 在肾脏、肝脏和皮肤的 WTL 中转录后增加,而铁蛋白轻链 (FTL1) 在主动脉和肾脏的 WTL 以及肾脏和皮肤的 LSF 中转录后增加,这表明转录后机制影响高龄时的铁稳态。他们还发现,HTRA1 是一种细胞外蛋白酶,已知其表达增加会诱导细胞外蛋白质沉积,其在主动脉和肺的 WTL 以及心脏、肺和皮肤的 LSF 中转录后增加。
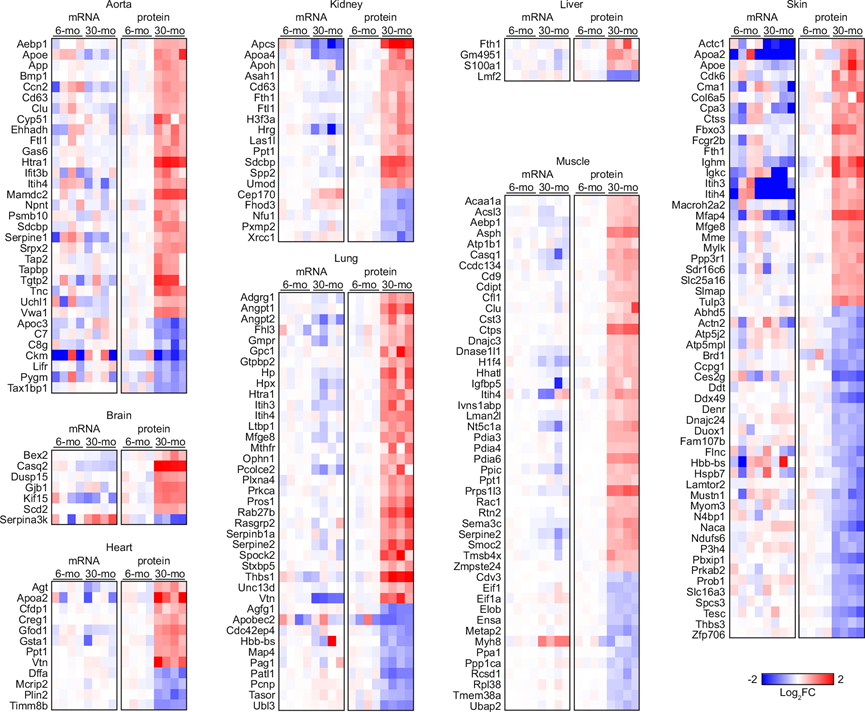
图6. WTL 中随年龄变化而出现差异表达的蛋白质,但没有伴随 mRNA 水平的类似变化。
热图显示了在 6 月龄和 30 月龄小鼠之间 WTL 中蛋白产物表达显著差异的基因的相对表达水平。
05
DEP 的细胞来源
公开的单细胞核转录组分析(RNA-seq)数据的分析表明,蛋白质随年龄增长而增加,蛋白质随年龄增长而减少,并且由不同类型的细胞产生(图 7)。30月龄小鼠中蛋白质和mRNA水平上表达水平增加的基因往往在免疫细胞中高表达,而随年龄增加的核心基质体蛋白质往往在基质细胞中高表达。另一方面,30月龄小鼠中表达水平降低的蛋白质往往在实质细胞中高表达。例如在肝脏中,蛋白质和mRNA水平上随年龄增加的蛋白质在免疫细胞中大量表达,而随年龄增加的核心基质体蛋白质在星状细胞中大量表达。相反,肝脏中几乎所有随年龄增加而减少的蛋白质主要在肝细胞中表达。
对于其他组织,在蛋白质和 mRNA 水平上随年龄增长而增加的蛋白质在心脏和肺的免疫细胞中表达最为丰富(图 7)。这一发现与本研究的结果一致,即随年龄增长而增加的常见 DEP 富含免疫相关蛋白(图 3e),并且往往在 mRNA 水平上存在差异表达。随年龄增长而增加的核心基质体蛋白在心脏的心内膜细胞、成纤维细胞、周细胞和平滑肌细胞,肾脏的成纤维细胞、近肾小球细胞、系膜细胞和足细胞,以及肌肉中的纤维脂肪形成祖细胞、平滑肌细胞和肌腱细胞中均在 mRNA 水平上高表达。30月龄小鼠中蛋白质和mRNA水平表达水平降低的基因在肾脏近端小管细胞中高表达。30月龄小鼠中蛋白质水平表达水平降低而非mRNA水平表达水平降低的基因在心脏的脂肪细胞、心肌细胞和施万细胞、肾脏的近端小管细胞以及肌肉的骨骼肌细胞中高表达(图7)。随年龄增长而降低的线粒体膜蛋白在这些细胞中也高表达(图 7)。值得注意的是,伴随mRNA水平变化的与年龄相关的蛋白质水平变化可能部分归因于组织内细胞群体的变化,尤其是免疫细胞的增加和实质细胞的减少。
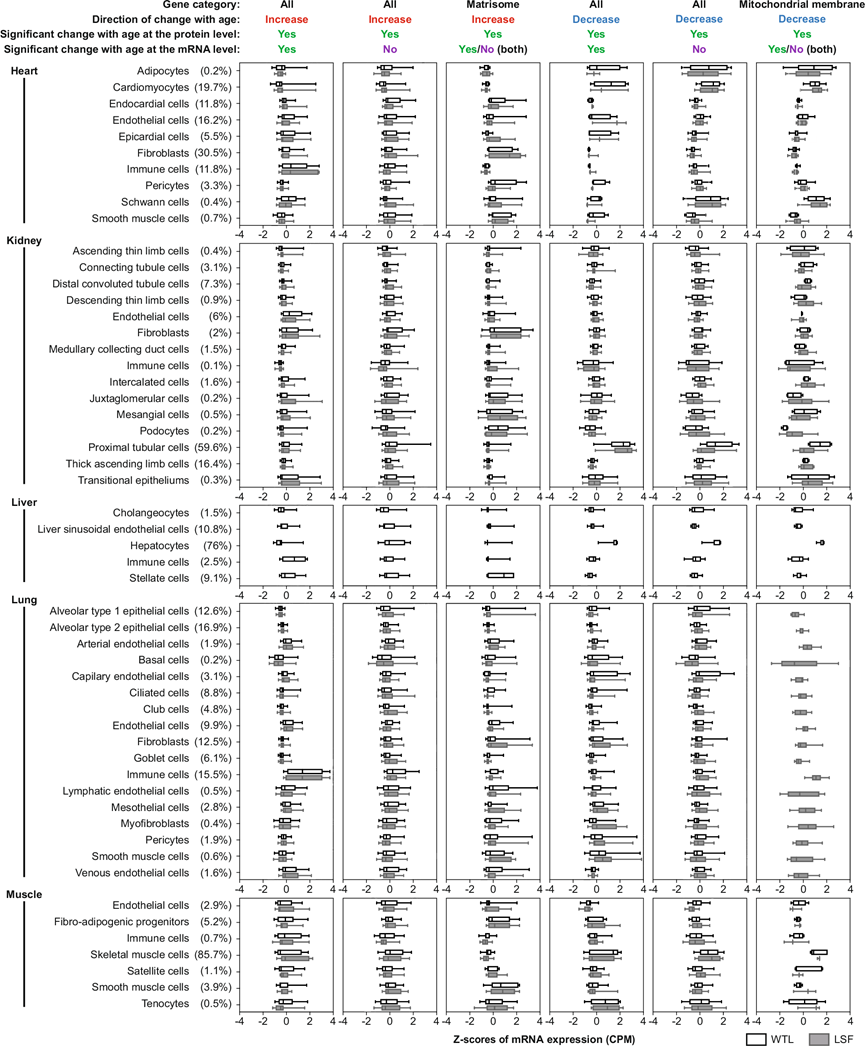
图7. 依赖于细胞类型的 DEP mRNA 表达水平。
箱线图显示了图顶部所示的DEP的细胞类型依赖性mRNA表达水平。
06
DEP 验证
最后,他们试图验证通过质谱蛋白质组学鉴定出的 DEP。首先关注 APOE,不仅因为其蛋白质水平在许多组织中随着衰老而增加,还因为对寿命和健康寿命影响最大的人类遗传多态性是 APOE 基因的多态性。APOE 主要在肝脏和大脑中产生,人们普遍认为其多态性通过影响阿尔茨海默病的发病机制来影响寿命,尽管最近的研究表明,无论阿尔茨海默病状态如何,APOE 多态性都与人类寿命有关。有趣的是,本研究的蛋白质组学数据显示,APOE 在主动脉、心脏、肾脏、肺、肌肉和皮肤中随着衰老而增加,但在大脑和肝脏中没有增加(图 3e),且通过免疫印迹法证实了这一发现(图 8a)。实时定量PCR分析显示仅在脑和肌肉中 APOE 的mRNA水平随着年龄显著增加(图8b)。免疫荧光染色显示老年心脏细胞外区域APOE增加(图 8c-d),提示循环APOE在细胞外沉积。另一种有趣的常见 DEP 是 COL18A1(图 3e),其 C 端裂解产物内皮抑素具有强大的抗血管生成活性。
他们还验证了 HSPA5 的年龄依赖性表达(图 8e-f),HSPA5 是一种组织特异性 DEP,仅在肌肉中随年龄增长而增加。最近对 8 月龄和 18 月龄小鼠组织的蛋白质组学研究也表明,HSPA5 在老年雄性小鼠肌肉中特异性增加。HSPA5是未折叠蛋白反应 (UPR) 的标志物,受 ER 应激激活转录因子 ATF6 和 XBP1调控,其上调表明UPR 的发生,而UPR 会调节衰老过程。他们研究了 UPR 通路三个主要分支的下游转录靶标,即 S1P-ATF6、PERK-ATF4 和 IRE1-XBP1随年龄相关的表达变化。ChIP-Atlas 数据库中鉴定的 XBP1 结合基因在衰老肌肉中表达增加,但 ATF4 结合基因则未出现表达增加(图 8g)。具有进化保守的 ATF6 结合序列的基因在 6 月龄和 30 月龄小鼠肌肉之间没有出现整体表达差异(图 8g)。UPR 调节因子及其已知转录靶标的实时定量 PCR 也表明,在衰老肌肉中 XBP1 通路发生激活,但 ATF4 和 ATF6 通路没有发生激活。XBP1、XBP1s(XBP1的活性剪接形式)及其靶基因Dnajb9和P4hb在衰老肌肉中上调,而 PERK 和 ATF6 通路中研究的基因均未随年龄增长而出现显著表达差异(图 8h)。为了评估 XBP1 激活在肌肉中的功能后果,他们对 15 个月大的小鼠进行了单次静脉注射,注射了编码小鼠XBP1s 的 AAV(受 MHCK7 启动子的控制)(图 8i)。转导 3 个月后,与转导含有非编码填充 DNA 的对照 AAV 的小鼠相比,转导了编码 XBP1s 的 AAV 的小鼠在转棒测试、跑步耐力测试和握力测试中的运动表现较低(图 8j-l)。这些结果表明,衰老肌肉中 XBP1s表达增加是与年龄相关的运动能力下降的原因之一。
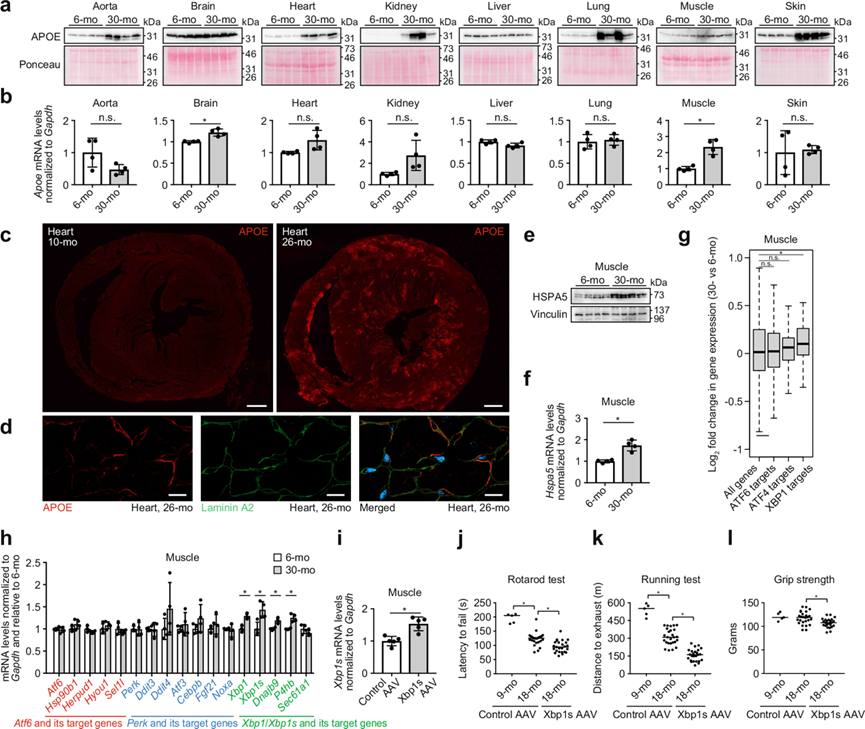
图8. 通过蛋白质组学分析确定的 DEP 的验证。
(a) 免疫印迹分析。(b) 通过实时 PCR 测量 6 个月大和 30 个月大小鼠的 Apoe mRNA 水平。(c) 代表性免疫荧光图像。(d) 小鼠心脏中 APOE 和层粘连蛋白 a2 的代表性共聚焦显微镜图像。(e)免疫印迹分析。(f) 通过实时 PCR 测量 6 个月大和 30 个月大小鼠的Hspa5 mRNA 水平。(g) 箱线图显示 6 个月大和 30 个月大小鼠肌肉中 ATF6、ATF4 和 XBP1 的假定靶基因的表达水平。(h) 通过实时 PCR 测量 6 个月大和 30 个月大小鼠的Atf6、Perk、Xbp1及其特征明确的靶基因的 mRNA 水平。(i) 通过实时 PCR 测量 18 个月大小鼠的Xbp1s mRNA 水平。(j-l) 小鼠的转棒能力、跑步耐力和握力分析。(l) (m) (n)
+ + + + + + + + + + +
结 论
本项研究提供了从 6、15、24 和 30 月龄小鼠中采集的WTL 和LSF的蛋白质组学数据。低溶解度蛋白质优先受年龄影响,而 LSF 的分析使随年龄变化而差异表达的蛋白质数量翻倍。使用相同组织匀浆同时分析蛋白质组和转录组揭示了与年龄相关的转录后失调的特征。转录后失调在 24 月龄小鼠中后尤其明显,与年龄相关的转录后失调导致核心基质蛋白的积累和多种组织中线粒体膜蛋白的减少。基于对成年、中年、老年和老年小鼠的深入蛋白质组学数据和样本匹配的转录组数据,本研究构建了小鼠衰老蛋白质组学图谱,该图谱提供了与年龄相关的基因表达变化的全面视图。
+ + + + +



