

 English
English文献解读|Cell Host Microbe(30.3):婴儿粪便微生物群和代谢物的昼夜节律:婴儿配方奶粉的随机对照干预试验
✦ +
+
论文ID
原名:Diurnal rhythmicity of infant fecal microbiota and metabolites: A randomized controlled interventional trial with infant formula
译名:婴儿粪便微生物群和代谢物的昼夜节律:婴儿配方奶粉的随机对照干预试验
期刊:Cell Host & Microbe
影响因子:30.3
发表时间:2024.04.02
DOI号:10.1016/j.chom.2024.02.015
背 景
婴儿肠道中的微生物群变化受到饮食的影响,母乳喂养和母乳低聚糖可促进有益双歧杆菌的定植。几乎所有生物体都具有生物钟,可以预测反复出现的环境变化。近年来,某些肠道细菌种类的丰度在个体和群体水平上表现出随时间变化的波动。越来越多的证据表明节律性丧失与疾病病理学之间存在联系。哺乳动物的昼夜节律系统在子宫内发育,在衰老过程中继续发展,并在发育过程中发挥重要作用。肠道菌群节律可以影响宿主体内生物钟基因的表达,细菌与宿主之间昼夜节律串扰的干扰会影响胃肠道健康和代谢。细菌的内源性时钟机制是否能够驱动复杂微生物群的节律性,从而影响宿主生理仍是未知的。
实验设计
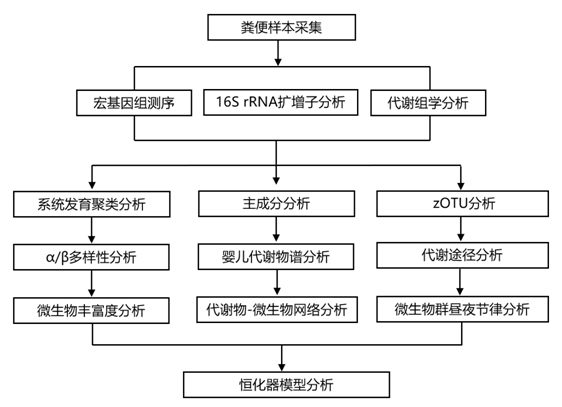
结 果
01

婴儿微生物群的时间发育分析
母乳喂养和母乳低聚糖可促进有益双歧杆菌的定植。研究团队在婴儿配方奶粉中添加了双歧杆菌或复合低聚糖,尤其是低聚半乳糖 (GOS),以模仿母乳。他们通过对 265 名孕妇进行资格评估,将 223 名婴儿纳入干预研究。在这些婴儿中,210 名积极参与并随机分配到四个不同配方奶粉组之一(图 1)。63名婴儿在干预期间(1年)接受母乳喂养。35名婴儿随机分配到不含额外补充剂的安慰剂组(配方A), 39名婴儿随机分配到含有长双歧杆菌和短双歧杆菌的配方B组,36名婴儿随机分配到含有GOS的配方C组, 37名婴儿随机分配到含有双歧杆菌和GOS的配方D组(图1)。他们在婴儿出生第一年的五个时间点收集粪便样本(图 2 A)。婴儿和父母粪便的纵向采样分析表明,婴儿的微生物群随着其父母微生物组成而发展(图2B-D)。β多样性分析表明,与亲本样本相比,3 - 24个月的微生物群特征发生了显著变化,但显示出明显不同的模式,而早期采样时间点(0.5个月、1个月和3个月)的特征是微生物群特征无法区分(图2B)。基于广义UniFrac距离的无监督分析发现,微生物群分布的显著异质性在很大程度上与喂养方式或分娩组(剖腹产与阴道分娩)无关(图2C)。亲本样本出现在两个不同的系统发育聚类中,两侧是来自较早采样时间点的样本,其中拟杆菌门和芽孢杆菌门占主导地位。放线菌的相对丰度在早期采样时间点变化很大,并随着年龄的增长而下降(图 2 C)。婴儿微生物群逐渐发育的特点是丰富度和 α 多样性的增加,在 3 个月大时达到显著水平。婴儿和父母之间平均个体丰富度的显著差异在 24 个月后消失(图 2D)。
无监督层次聚类在出生第一年的所有婴儿样本中识别出三个不同的聚类(图 2 E)。这些聚类的特点是双歧杆菌(聚类 1)、肠杆菌(聚类 2)和拟杆菌(聚类 3)的相对丰度 (%) 较高且不成比例(图 2 F)。双歧杆菌类占主导地位的聚类富集于母乳喂养的样本,主要由早期时间点的粪便样本组成。配方奶粉摄入在3个月大时达到峰值,母乳喂养同时随着配方奶粉摄入的增加而减少(图2G)。与 3 个月后配方奶粉摄入量的减少一致(图 2 G),97.5% 的婴儿在第 7 个月时接受了固体食物。来自干预措施和母乳喂养婴儿的样本中的聚类 2 和聚类 3 中的比例相同(图 2 H)。此外,通过剖腹产出生的婴儿的粪便样本的聚类2比例过高,而通过阴道分娩的婴儿粪便样本中聚类3占比过高。

图1. 参与者入组和随机化的研究示意图。
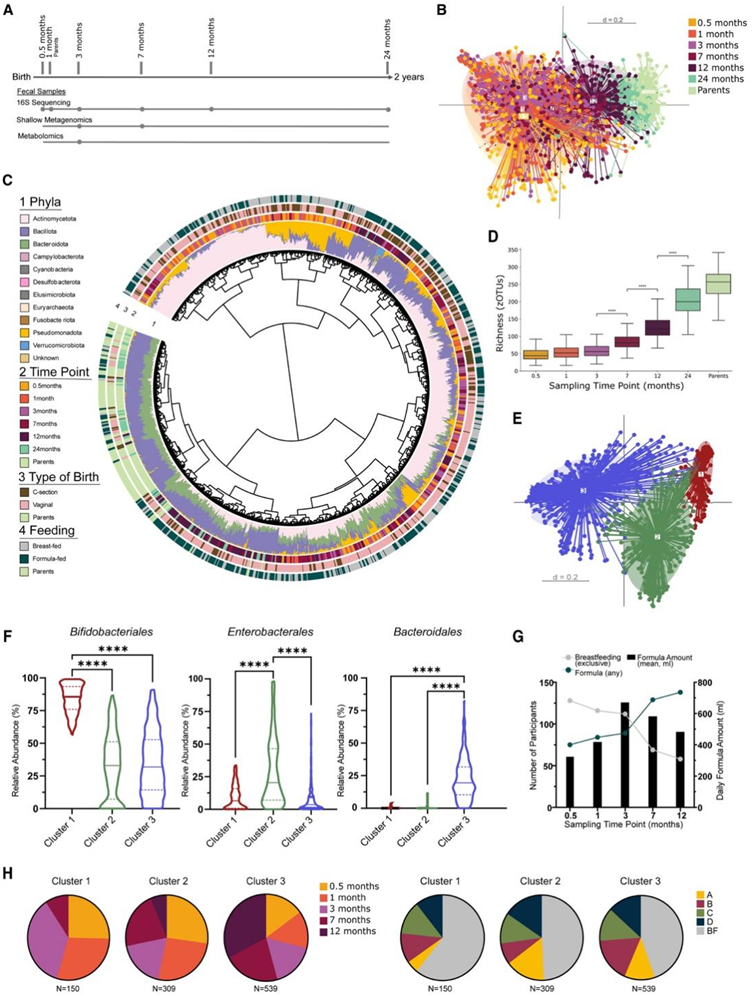
图2. 婴儿微生物群的时间发育。
(A) 研究粪便样本采集和粪便样本分析时间点的示意图。(B) 根据婴儿年龄(月)或父母样本分组的粪便样本的非度量多维标度 (NMDS) 图。(C)所有研究粪便样本(来自 210 名婴儿 [多个时间点,包括随访] 和 152 名父母 [单个时间点] 的 1222 份粪便样本)的从头系统发育聚类显示为树状图。 (D) 不同婴儿采样时间点和父母样本的 Alpha 多样性。(E) 所有婴儿样本的 β 多样性(无监督聚类)的度量多维标度 (MDS) 图,产生三个不同的聚类。(F) 前 3 个相对丰度 (%) 聚类主导细菌顺序小提琴图。(G)平均每日配方奶粉摄入量(mL,条形图)和纯母乳喂养率与配方奶粉消费者的对比。(H)关于采样时间点和喂养类型的描述性聚类组成。
02
双歧杆菌和 GOS 干预对婴儿微生物群变化的影响
广义UniFrac距离显示,四种不同补充婴儿配方奶粉(A-D)和纯母乳喂养组的群落结构之间没有显著差异(图3A),这表明个体间的差异在很大程度上超过了基于喂养方式的微生物群分层。随着婴儿年龄的增长,不同喂养组的物种丰富度增加相同(图3B)。水平斑点图总结了前10个分类群的相对丰度随时间的变化(图3C)。双歧杆菌是所有婴儿粪便样本中最丰富的属(41.2%),在婴儿出生后3 个月大时达到峰值丰度(图 3 C)。双歧杆菌的平均丰度在安慰剂 A 组中最低(32.3%),其次是补充双歧杆菌的 B 组(39.7%)。拟杆菌、埃希氏志贺氏菌、韦永氏球菌、克雷伯菌、梭状芽孢杆菌、链球菌和肠球菌在非常早期的微生物群落(0.5、1和3个月)中的丰度水平明显较低(图 3 C)。在第7个月和第12个月,拟杆菌属和布劳特氏菌属的平均丰度明显高于中位数(图 3 C)。尽管随着时间的推移存在一些差异,但配方奶喂养的婴儿比母乳喂养的婴儿平均高出29%的韦永氏球菌丰度(图 3 C)。在第3个月和第7个月,与对照组A相比,干预组C组和D组补充GOS显著增加了双歧杆菌的平均丰度(图3D),但配方中补充长双歧杆菌和短双歧杆菌(B组)未能增加双歧杆菌的丰度。母乳喂养和配方奶粉喂养婴儿之间双歧杆菌丰度的比较显示,在3个月时,任何干预组之间的显著性完全消失,表明样本之间的内在变异性高于此时间点干预的效果。干预措施之间未发现其他差异。浅宏基因组分析发现,长双歧杆菌、短双歧杆菌、大肠杆菌和两歧双歧杆菌是第3个月和第7个月最丰富和流行的5种细菌(图3E)。一些母乳喂养的婴儿缺乏最常见的双歧杆菌菌株,反而含有不同的韦永氏球菌和粪肠球菌(图3E)。基于宏基因组学的相关分析确定了与氨基酸生物合成、核苷和核苷酸生物合成、细胞结构生物合成以及辅因子、载体和维生素生物合成相关的途径(图3F)。
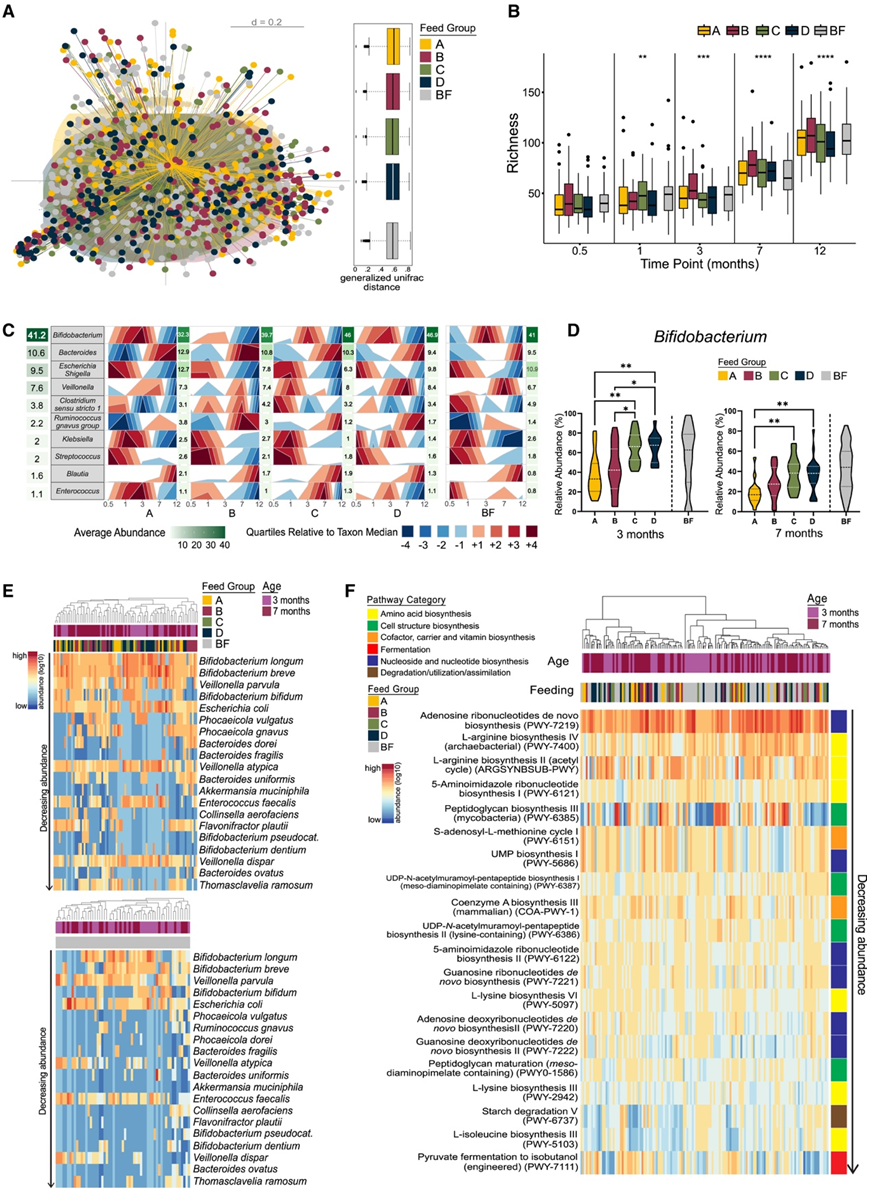
图3. 双歧杆菌和GOS干预对婴儿微生物群组装的影响。
(A) 婴儿β多样性(MDS 图)。(B) 不同喂养组随时间变化的丰富度。(C) 在 >80% 的样本中发现的前 10 个相对丰度 (%) 类群(属)。(D)双歧杆菌属相对丰度的频率分布。 (E) 单个样本的前 20 个物种的相对丰度。(F) 与按丰度递减排序的宏基因组测序样本相关的前 20 个已识别途径。
03
不同喂养组之间的代谢物明显不同,并表现出昼夜节律性
接下来,他们分析了 3 个月大时的代谢环境,因为尚未引入固体食物,而婴儿在这个时间点摄入的配方奶量最多。主成分分析 (PCA) 发现母乳喂养和配方奶喂养的婴儿的代谢物聚类明显分离,而配方奶喂养的婴儿代谢物仍然无法区分(图 4 A)。有趣的是,纯配方奶粉喂养的婴儿代谢物聚类在远离母乳喂养的婴儿的地方,而混合喂养(接受母乳和配方奶粉)与母乳喂养的婴儿代谢物聚类部分重叠(图4A),这表明母乳或婴儿配方奶粉的摄入在各自的粪便代谢特征中发挥着重要作用。
他们对103份母乳样本和4种不同配方奶的人乳和配方奶的代谢物谱进行了额外的分析(PCA)。在这里,证明了配方奶粉样本明显远离所有母乳喂养的样本。在婴儿的母乳和粪便样本中发现了母乳低聚糖,例如岩藻糖基乳糖 (FL)(80% 的 3 个月粪便样本中 FL 呈阳性)。代谢物丰度的聚类分析用于根据喂养组对模式进行分层(图 4 B)。正如 49 个最丰富特征的热图中所示,母乳喂养和配方奶喂养的婴儿存在差异代谢物聚类,母乳喂养的聚类中仅偶尔出现配方奶喂养婴儿的样本中(图 4 B)。对各个代谢物的更详细分析表明,磷酸胆碱以及一些特殊的糖衍生物(如糖酸、N-乙酰化四糖或 N-乙酰化三糖)在母乳喂养组中增加,而肉碱在所有配方奶喂养组中增加。吲哚衍生物,例如细菌来源的吲哚乳酸,在配方奶喂养的 B、C 和 D 组中也有所增加(图 4 C)。采用网络相关分析来确定细菌种类和粪便代谢物的关联(图4D)。前十个显著细菌种类与代谢物呈正相关或负相关,表明类群之间的功能冗余(图 4 E)。为了评估这种情况是否也发生在一岁的婴儿身上,他们分析了 3 个月大时代谢物丰度的白天依赖性波动。热图中显示了所有粪便样本中粪便代谢物的峰丰度,证明存在昼夜节律(图 4 F)。网络相关分析进一步通过零半径操作分类单元 (zOTU) 关联细菌,如长双歧杆菌和韦永氏球菌与节律性代谢物联系起来(图4G)。
这些发现支持这样的假设:昼夜节律调节在生命的早期阶段就开始,可能对婴儿微生物组的功能做出贡献。
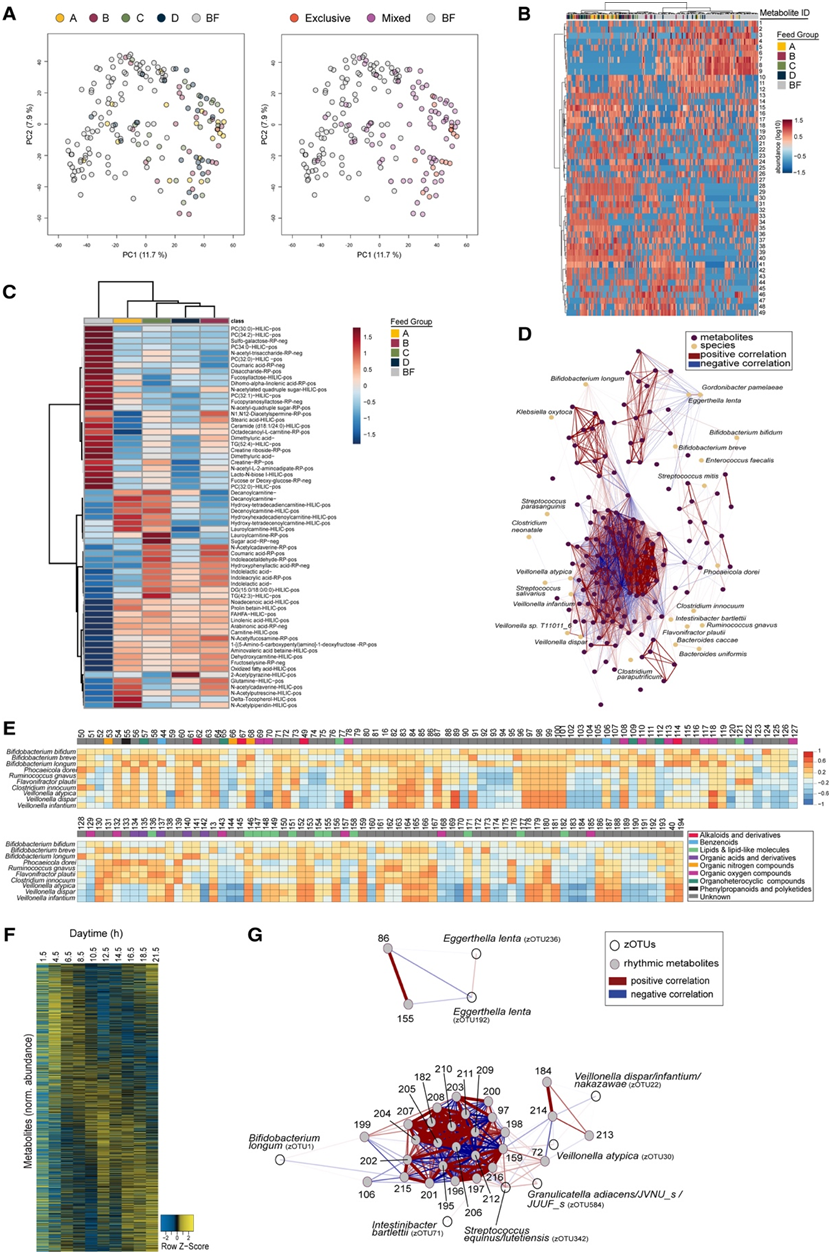
图4. 代谢物清楚地区分喂养组并显示昼夜节律变化。
(A)婴儿代谢物谱(3 个月)的主成分分析。(B) 样本内归一化热图,描绘了婴儿粪便样本(3 个月)中发现的前 49 种代谢物的相对丰度。(C) 每个喂养组显示的注释粪便代谢物的平均丰度(3 个月)。(D) 使用宏基因组测序鉴定的粪便代谢物和微生物的相关网络。(E) 显著相关物种(宏基因组测序)和粪便代谢物的热图。 (F) 一天中不同时间段粪便代谢物标准化丰度的热图。(G) zOTU 和节律代谢物的网络相关分析。
04
婴儿微生物群昼夜节律的发育受到年龄和配方奶粉补充的影响
根据白天依赖的代谢物丰度差异及其与不同细菌分类群的联系(图4G),在白天和夜间从婴儿收集的粪便样本的细菌丰富度和α多样性中发现了显著的昼夜节律(图5A)。昼夜节律也存在于单个 zOTU 水平上,如 24 小时内标准化峰值丰度所示(图 5 B)。有趣的是,检测到高丰度的双歧杆菌属具有显著的节律性(图 5 C)。随着婴儿的成熟,昼夜 zOTU 的数量也随之增加,同时婴儿肠道微生物群的多样性也不断增加,节律性 zOTU 的数量在 12 个月时达到最高(图 5 D)。有趣的是,添加双歧杆菌和GOS的配方奶粉组中节律性细菌的数量最高,甚至超过了母乳喂养婴儿中节律性zOTU的数量(图5E)。节律性zOTU的数量最多的属为韦氏菌属(7个zOTU)、拟杆菌属(6个zOTU)、双歧杆菌属(6个zOTU)、链球菌属(6个zOTU)和梭状芽孢杆菌属(5个zOTU)。在7个月大时,配方奶粉组D(604)的样本中,显著节律性分配途径的总量占主导地位,其次是母乳喂养组,在162个途径中显示出节律性(图5F)。
为了研究生命早期观察到的微生物节律是宿主驱动的还是依赖于微生物内在时钟机制,他们在恒化器模型中培养了来自选定的纯母乳喂养和配方奶喂养(A 组)婴儿的 3 个月粪便样本。为了建立稳定的生长条件,设置了初始适应阶段(分批培养),然后在恒化器中连续培养 7 天,以确保宿主独立条件(图 5 G)。含有母乳喂养供体粪便样本的恒化器之间的 Beta 多样性相似,但在喂养条件之间存在显著差异。从连续培养的第 7 天开始,在 48 小时的过程中每 4 小时取样一次(每个恒化器 13 个样本)(图 5 G)。对微生物群组成的昼夜节律分析揭示了在优势门、科和属水平上说明的所有恒化器中真正的内源昼夜节律。zOTU(图5I)和选定的代谢途径分析突出了明显的节律变化。根据婴儿粪便样本中的结果(图5C-E),发现双歧杆菌属和肠杆菌科的zOTU在母乳喂养的样本中过多存在,而Clostridium sensu stricto 1在配方奶粉A的恒化器样本中过少存在(图5I)。这些结果表明,复杂微生物群中的特定分类群在连续培养7天后会发生昼夜节律性,这表明婴儿细菌具有不依赖于宿主的内在节律性。

图5. 婴儿微生物群日间相关节律的发育受到配方奶补充剂的影响。(A) 已知排便时间的样本的 α 多样性的昼夜分布。(B) 在婴儿样本中检测到的 777 个 zOTU 的相对丰度热图。(C)所有婴儿样本中双歧杆菌属的昼夜分布。(D) 0.5、3、7 和 12 个月时的节律性 zOTU,节律性 zOTU 总数。(E) 五个喂养组中的节律性 zOTU,图底部显示节律性 zOTU 总数。(F) 按喂养组分开的婴儿样本(3 个月和 7 个月)中节律和心律失常通路的量化和意义。(G) 含有从母乳喂养(BF)和安慰剂配方奶喂养婴儿粪便培养物的趋化因子1-4的研究设计和采样时间点示意图。 (H) zOTU 的系统发育聚类,显示为分支图。(I) 连续培养 7 天后来自恒化器培养物的 zOTU 的昼夜节律图谱。
+ + + + + + + + + + +
结 论
这项随机对照干预研究(德国临床试验 DRKS00012313)对婴儿出生第一年的粪便进行了纵向采样,揭示了配方奶喂养和母乳喂养婴儿之间相似的粪便细菌群落。与含有长双歧杆菌和短双歧杆菌或安慰剂的婴儿配方奶粉相比,含有 GOS 的婴儿配方奶粉中双歧杆菌的含量持续较高。代谢组学和微生物组学分析揭示了 24 小时波动和昼夜节律网络。细菌多样性、特定分类群和功能途径的节律性随着年龄的增长而增加,并且在母乳喂养和补充 GOS 后最强。在恒化器模型中,优势类群的昼夜节律在体外得到进一步维持。因此,微生物群的节律性在生命早期就形成并受到饮食的影响。
+ + + + +



