

 English
English文献解读|Cell Rep Med(14.3):独特的多细胞免疫抑制中心为左侧和右侧结肠癌提供不同的干预策略
✦ +
+
论文ID
原名:Distinctive multicellular immunosuppressive hubs confer different intervention strategies for left- and right-sided colon cancers
译名:独特的多细胞免疫抑制中心为左侧和右侧结肠癌提供不同的干预策略
期刊:Cell Reports Medicine
影响因子:14.3
发表时间:2024.05.07
DOI号:10.1016/j.xcrm.2024.101589
背 景
结肠癌是一种高度异质性的疾病,是全球癌症相关死亡的主要原因。根据原发部位,它进一步分为左侧结肠癌和右侧结肠癌,每种结肠癌都有不同的临床和分子特征。与左侧结肠癌 (LCC) 相比,右侧结肠癌 (RCC) 更常与微卫星不稳定性高 (MSI-H) 以及KRAS和BRAF突变相关。多项研究表明,肾细胞癌的总体生存率较低。然而,这些特征与 LCC 和 RCC 的临床结果和治疗反应之间的关联仍不清楚。
实验设计
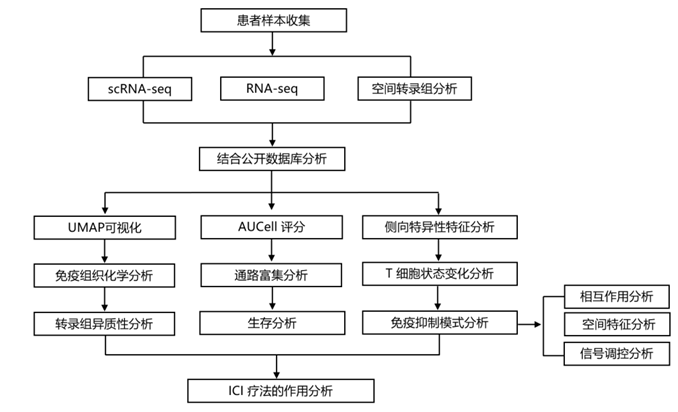
结 果
01

LCC 和 RCC 之间的单细胞表达图谱
本项研究包括发现队列和验证队列,使用无监督方法进行聚类分析并获得细胞亚群。根据亚群的转录组特征,鉴定出九种主要细胞类型(图 1 A-B)。根据肿瘤所在侧对每位结肠癌患者的全局细胞类型注释进行分类。尽管比例不同,但所有这些细胞类型在患者之间以及 LCC 和 RCC 之间都是相同的(图 1 C)。与 LCC 相比,RCC 中的髓细胞和 T 淋巴细胞的相对丰度增加,而 B 淋巴细胞和浆细胞的相对丰度降低(图 1 D)。进一步进行了七重免疫组织化学 (IHC) 染色,以概述左侧和右侧结肠癌的多细胞生态系统(图 1 E)。这些结果表明 RCC 代表了与 LCC 不同的多细胞生态系统。
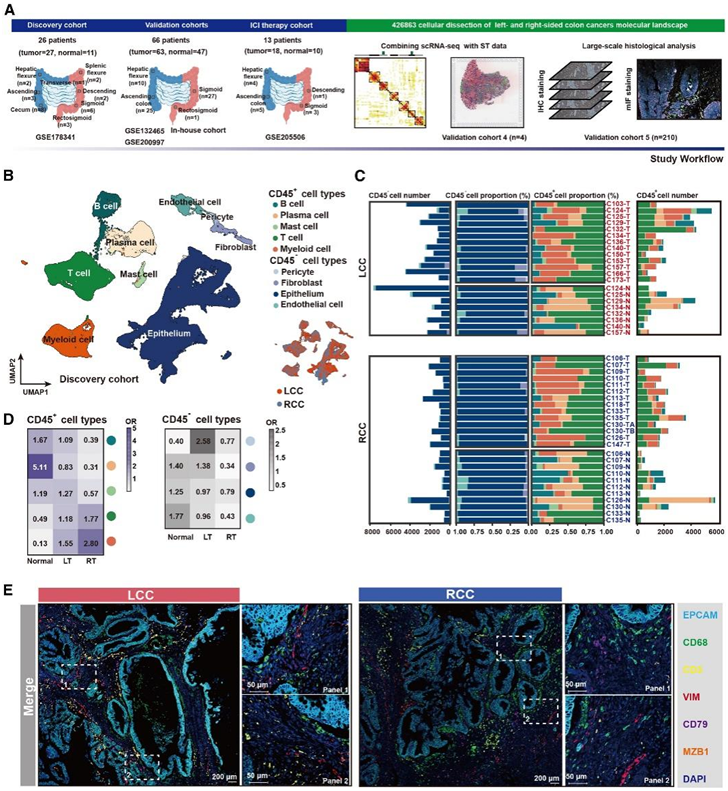
图1. lcc和rcc单细胞转录组图谱。
(A)本研究样本采集及分析过程。(B)UMAP可视化。(C)单个样本正常粘膜和结肠肿瘤组织中主要细胞类型的比例,按左结肠癌(LCC)和右结肠癌(RCC)分组。(D)热图显示各组织中主要细胞类型的优势比(OR)。(E)左右两侧肿瘤组织中的多重免疫组化(mIHC)染色分析。
02
LCC 和 RCC 之间恶性上皮的转录组异质性
以免疫细胞和基质细胞为参考推断大规模拷贝数变异(CNV)而识别的恶性上皮具有染色体缺失和拷贝数增加,与之前的报道一致(图 2 A)。总体而言,他们从 27 个包含至少 100 个恶性上皮的肿瘤样本中鉴定出 50450 个恶性上皮,并保留以进行进一步分析(图 2 B)。恶性上皮的聚集揭示出患者特异性和侧特异性聚类(图 2 C-D)。为了表征恶性上皮的肿瘤异质性,他们基于非负矩阵分解 (NMF) 分析了每个肿瘤中共表达基因的聚类,并获得了总共 96 个肿瘤内程序。肿瘤内程序可以反映肿瘤的共同特征,进一步的层次聚类分析确定了肿瘤中七个突出的元程序(图 2 E),每个程序包含一组独特的基因。基于元程序的基因集,计算了基因评分,这最终能够定义主要的细胞状态。值得注意的是,增殖干性 (PS) 和免疫分泌 (IS) 元程序在左侧和右侧肿瘤的恶性上皮中显示出相反的表达模式(图 2 F)。
PS 元程序的特点是表达与增殖相关的基因(例如UBE2C、MYBL2和PRKDC)和与干细胞相关的基因(例如ALDH1A1和DCLK1)。IS 元程序增加了主要组织相容性复合体 (MHC) II 类分子(例如CD74、HLA-DRB1和HLA-DPA1)和分泌蛋白分子(例如REG4、AGR2和AGR3)的表达。ALDH1A1、UBE2C和MYBL2是 PS 表达元程序中相对权重最高的三个基因,主要在左侧结肠上皮中表达。在左侧肿瘤 (LT) 中,表达这三个基因的上皮比例与 scRNA-seq 数据中的 PS 元程序活性相关。
总体而言,可以通过特征基因的集体表达来预测左侧和右侧结肠肿瘤的侧面特异性元程序活性。验证队列 5 中的特定标志物的 IHC 证实了恶性上皮中侧面特异性元程序的表达(图 2 G-H)。此外,PS 和 IS 元程序的 IHC 染色评分(H 分数)(以特征基因的集体表达为代表)分别与 LCC 和 RCC 的晚期阶段相关(图 2 I)。

图2. 恶性上皮内表达元程序的鉴定。
(A) 热图显示代表性患者(LCC 为 C125,RCC 为 C109)个体细胞大规模 CNV 的染色体图。(B) 84975 个上皮细胞的 UMAP 图,按恶性和非恶性细胞着色。(C) 恶性上皮细胞聚类的 UMAP 图,按患者(左)或癌症类型(右)着色。(D) 堆叠直方图显示每个恶性上皮聚类的患者比例(上)或癌症类型(下),按患者或癌症类型着色。(E)热图描绘了所有患者共有的表达元程序。(F) 恶性上皮中的 PS 和 IS 元程序在左侧和右侧结肠癌中的活性有显著差异。(G) 验证队列 5 中 PS(上)和 IS下元程序的特征基因产生的蛋白质的代表性免疫组织化学 (IHC) 染色。(H) 左侧肿瘤(LT)和 右侧肿瘤(RT)之间 PS 和 IS 特征基因表达的 H 分数。(I) 早期和晚期结肠肿瘤组织中 PS 和 IS 染色评分的分布。
03
表达元程序与致癌特性和预后的关联取决于原发肿瘤的位置
接下来,他们考虑了单侧性特异性元程序的致癌特性和预后意义。通过建立 PS 元程序活动的最佳阈值 (AUC > 0.15),将左侧肿瘤(LT)中的恶性上皮分为 PShigh组和 PSlow组(图 3 A)。基因集富集分析 (GSEA) 显示,糖酵解和增殖相关通路在 PShigh恶性上皮中显著富集(图 3 B)。通过分析增殖标志物 Ki-67 来验证恶性上皮的增殖状态(图 3 C)。乳酸脱氢酶 A (LDHA) 表达通过增加乳酸和质子的产生来驱动有氧糖酵解;因此,它的表达是增加有氧糖酵解的分子指标(图 3 C)。在验证队列 5 中,IHC 染色显示 LT 恶性上皮中的 Ki-67 和 LDHA 蛋白表达水平明显高于右侧肿瘤(RT)(图 3 D)。在 LT 中,PS 元程序的 IHC 染色评分与 Ki67 和 LDHA 的蛋白表达呈正相关。LDHA 通过将丙酮酸转化为乳酸来执行糖酵解信号的最后一步。通过分析 scRNA-seq 数据,他们证实了恶性上皮中 PS 元程序表达与 LT 中的糖酵解信号之间存在正相关(图 3 E)。此外,观察到 PS 元程序表达与增加的 MCT4 表达之间存在关联(图 3 E)。肿瘤细胞中乳酸转运蛋白的过度表达可减缓应激加速生成,而MCT4是糖酵解细胞中表达的有效乳酸输出蛋白。mIHC 染色进一步验证了 LT 中 PShigh恶性上皮的富集,同时伴有 MCT4 表达的升高(图 3 F)。
根据AUC > 0.15的阈值将RT恶性上皮分为IShigh组和ISlow组(图3G)。缺氧和免疫相关通路在IShigh恶性上皮中显著富集(图3H)。在验证队列5中,IHC染色结果显示,与LT相比,RT中恶性上皮的缺氧增加,如较高的CAIX染色评分所示(图3I-J)。CAIX蛋白表达水平也与RT恶性上皮IS元程序评分呈正相关。通过对scRNA-seq数据的分析。他们进一步证实了恶性上皮中IS 元程序的表达与RT期缺氧信号有很强的相关性,同时伴有CAIX表达的升高(图3K)。在本研究队列中,观察到吸烟的肾癌患者的恶性上皮表现出更高的IS活性和缺氧评分。此外,在TME中异常增强的转化生长因子β (TGF-β)和干扰素γ (IFN-γ)阻碍免疫监视和促进癌症发展(图3H)。他们发现IS元程序富集评分与免疫特征评分(ESTIMATE)之间存在很强的相关性。
接下来,他们评估了两个单细胞定义的侧特异性表达元程序与通过大量基因表达确定的共识分子亚型 (CMS) 组的关系。使用 IS 和 PS 元程序中的基因集计算癌症基因组图谱 (TCGA) 结肠腺癌 (COAD) 队列和基因表达综合 (GEO) 中独立结肠癌队列中每个肿瘤的元基因表达评分。PS 元程序富集评分较高的 LCC 患者 5 年总生存率 (OS) 较差,但 RCC 患者并非如此(图 3 L)。相反,RCC 患者的 IS 元程序富集评分较高与较差的 5 年 OS 相关,但 LCC 患者的 IS 元程序富集评分较高与较差的 5 年 OS 相关(图 3 M)。在 LCC 中,PShigh亚组与 CMS2 密切相关(图 3 L),其特征是典型的 WNT 和 Myc 激活。另一方面,肾细胞癌中的IShigh亚群与CMS1亚群有很大的重叠(图3M),CMS1亚群在TME中以强大的免疫细胞浸润为特征,并包括微卫星不稳定(MSI)/错配修复(MMR)。侧特异性元程序亚型系统进一步将 LCC 中的 CMS2 或 RCC 中的 CMS1 分层为不同的亚组。与其他亚组相比,PShigh/CMS2 和 IShigh/CMS1 的预后最差(图 3 L-M)。
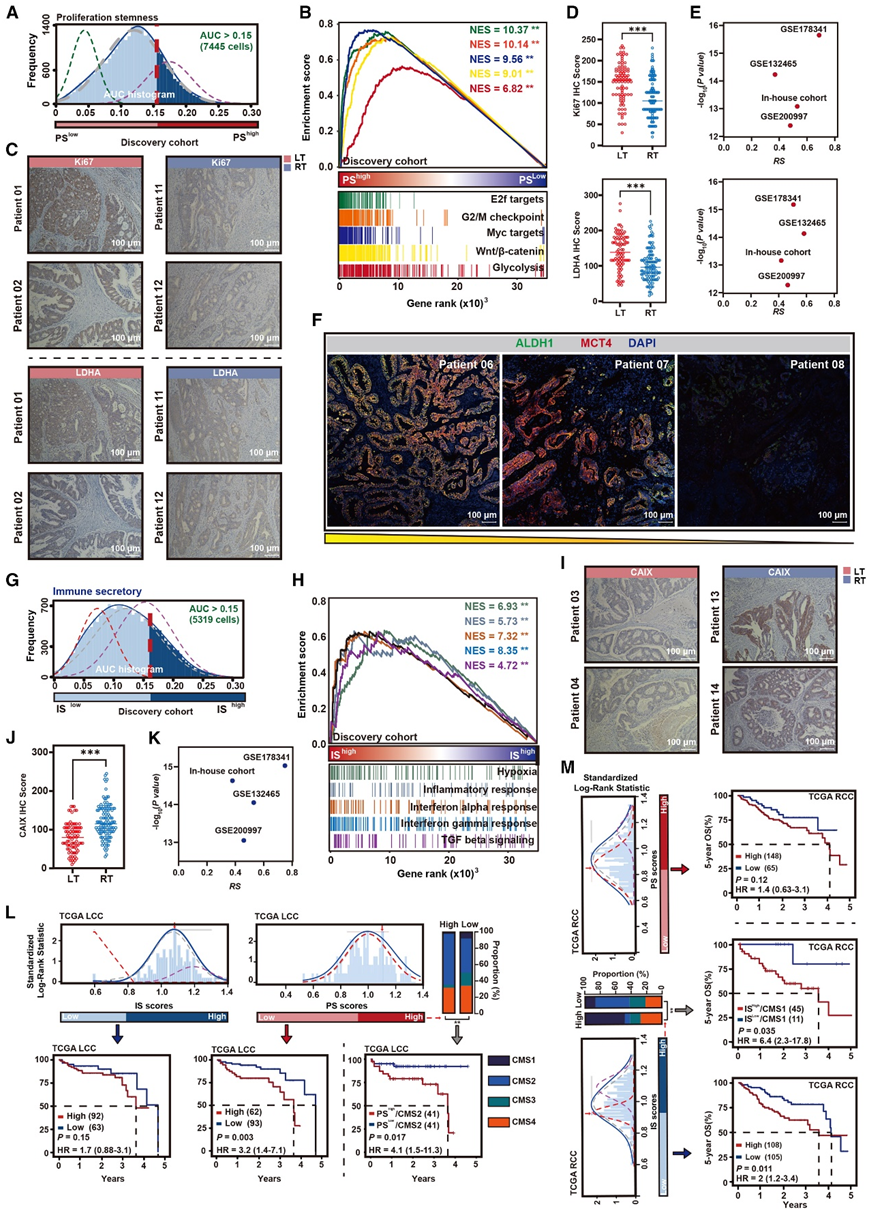
图3. 恶性上皮细胞侧向特异性表达元程序。
(A) 尖峰图显示左侧恶性结肠上皮的 PS AUCell 评分分布。(B) GSEA 中 PShigh与 PSlow左侧恶性结肠上皮的前 5 种富集通路。(C) 验证队列 5 中的 Ki-67和 LDHA的 IHC 染色。(D) 左侧和右侧结肠肿瘤组织中 Ki67 和 LDHA 染色评分的分布。(E) 左侧恶性结肠上皮的 PS 评分与有氧糖酵解评分和 MCT4表达之间的 Spearman 相关性。(F) LT 的代表性 mIHC 染色。(G) 峰值图显示右侧恶性结肠上皮中 IS AUCell 评分分布。(H) GSEA 中 IShigh与 ISlow右侧恶性结肠上皮的前 5 种富集通路。(I) LT 和 RT 中的 CAIX 的 IHC 染色(验证队列 5)。(J) 左侧和右侧结肠肿瘤组织中 CAIX 染色评分的分布(验证队列 5)。(K) 右侧恶性结肠上皮的 IS 评分和缺氧评分之间的 Spearman 相关性。(L) 尖峰图显示 LCC 患者中不同 PS 和 IS的标准化对数秩统计量。(M) 尖峰图显示具有bulk RNA-seq数据 (TCGA-COAD) 的 RCC 患者中不同 PS 和 IS的标准化对数秩统计量。
04
侧特异性TME重塑骨髓细胞的组成
骨髓细胞在肿瘤组织中的扩增表明它们在形成TME中起着积极的作用。他们捕获了18个具有异质基因表达模式的髓系亚群,包括三类:树突状细胞(DC) (C1-C6)、单核细胞(C7-C10)和巨噬细胞(C11-C18)(图4A-B)。每侧骨髓亚群的分布模式不同(图4C-D),显示了侧特异性特征。分泌型磷蛋白1 (SPP1)巨噬细胞(Mph-SPP1)是结肠肿瘤中最丰富的亚群,其中大部分来源于RT。RT中Mph-SPP1细胞上调的基因显著富集于缺氧、血管生成和EMT通路的正调控(图4E)。这些发现表明特定信号通路的激活与 Mph-SPP1 细胞的分化及其各侧特异性的肿瘤促进作用有关。在 TCGA 和 GSE39582 结肠癌队列中,Mph-SPP1 细胞浸润较高的患者的 5 年 OS 率较短。基于 mIHC 染色,与 LT 相比,SPP1+巨噬细胞在 RT 中特异性富集(图4F-G)。RT中SPP1+巨噬细胞的富集与较低的OS率和晚期肿瘤分期显著相关(图4I-H)。

图4. 髓系细胞的特征。
(A) 髓系细胞的 UMAP 图。(B) 气泡热图显示髓样聚类中的标记基因。(C)按样本类型(正常、LT 或 RT)着色的 UMAP 图。(D) 热图显示每个组织中出现的髓样聚类的 OR。(E) 与 RT 中的其他巨噬细胞亚群相比,Mph-SPP1 细胞中的基因组 GSEA 富集。(F) LT 和 RT 中SPP1 +巨噬细胞的代表性 mIHC 染色(验证队列 5)。(G) 散点图显示基于 mIHC 染色结果的左侧和右侧结肠肿瘤中SPP1 +巨噬细胞的密度。(H) Kaplan-Meier 曲线显示不同 SPP1 +巨噬细胞浸润的 RCC 患者 OS。(I)早期和晚期肾细胞癌患者肿瘤切片中SPP1 +巨噬细胞的密度。
05
LCC和RCC之间T淋巴细胞状态的变化
在邻近的非肿瘤组织和肿瘤组织中,T淋巴细胞构成了最大的免疫细胞聚类(图1C)。T淋巴细胞无监督聚类分析共鉴定出23个侧别分布趋势不同的细胞亚群(图5A-B)。自然杀伤(NK)细胞显示出三个亚群,均富集RT(图5A-C)。NK-CD160细胞表现出以CD69、NR4A1和CXCR6高表达为特征的组织驻留记忆表型转录特征。NK-XCL1 细胞的转录特征与 NK-CD160 细胞相似。不同之处在于,NK-XCL1 细胞表现出更高的幼稚相关基因(TCF7、CCR7和SELL)表达水平和更低的细胞毒性相关基因(GZMA、GZMB、GZMK和IFNG)表达水平,表明杀死肿瘤细胞的能力存在差异。NK-CD16 细胞过度表达细胞毒性基因(GNLY、GZMB和PRF1)和抑制相关基因HAVCR2,表明它们可能出现功能障碍。
在 CD4 + T 细胞区中,他们鉴定出代表不同 T 细胞功能状态的典型 CD4+ T 细胞亚群,包括基于典型标记基因表达的幼稚(CD4-CCR7;CCR7+LEF1+)、效应记忆(CD4-GZMK;GZMK+ CCL5+)、TH1 样(CD4-CXCL13;CXCL13 + IFNG+)、TH17 样(CD4-IL23R;IL23R+ RORC+)、滤泡 T 辅助(CD4-CXCR5;CXCR5 + BCL6+)、调节性(CD4-FOXP3;FOXP3+ IKZF2 +)和增殖调节性(CD4-CTLA4;CTLA4+MKI67+)T 细胞状态。与RT相比,LT中CD4+ T细胞比例较高(图5C)。调节性T细胞(Treg)(CD4-FOXP3和CD4-CTLA4)高表达的IL2RA、FOXP3和CTLA4在CD4+ T细胞亚群中所占比例最高,且在肝移植中显著升高(图5B-C)。CD4-FOXP3细胞和CD4-CTLA4细胞具有相似的转录特征,但不同之处在于CD4-CTLA4细胞强表达增殖相关基因,这意味着它们在应答肿瘤抗原时发生克隆扩增。
接下来,他们研究了动态细胞转移,以进一步确认两种肿瘤类型中CD8+ T细胞的不同免疫状态。在CD8+ T细胞亚群中,他们首先鉴定了代表不同功能状态的典型CD8+ T细胞亚群,包括幼稚T细胞(CD8-LEF1;CCR7+LEF1+)、中央记忆(CD8-GPR183;SELL+GZMA+)、效应存储器(CD8-GZMK;GZMK+CXCR4+)、近期激活的效应细胞记忆或效应细胞(CD8-CXC3R1;KLRG1+CXC3R1+)、耗尽(CD8-LAYN;HAVCR2+PDCD1+)和组织驻留记忆(CD8-CD6;CD6+XCL1+) T细胞和上皮内淋巴细胞(CD8-CD160;CD160+NR4A1+)(图5A)。伪时间分析显示,CD8-LEF1细胞位于轨迹路径的起点,而CD8-CD160细胞和CD8-LAYN细胞位于轨迹路径的末端(图5D)。在这一转变过程中,细胞毒性特征评分降低,而耗尽特征评分升高(图5E)。早期CD8+ T细胞主要分布在RT中,少数细胞在细胞状态转换路径的末端,而LT中的CD8+ T细胞主要分布在细胞毒性和耗竭状态的末端(图5F)。此外,CD8- LEF1细胞位于CD8+ T细胞分化的早期阶段,在RT中显著富集(图5C)。他们评估了RT来源的CD8-CD161 (CD8-LEF1) T细胞中细胞毒性相关基因的表达水平,并验证它们显示出较低的细胞毒性能力(图5G)。GSEA显示,RT中的CD8-CD161以OX40下调和T细胞信号通路枯竭为特征,表明这些细胞处于激活程度较低但未耗尽的状态(图5H)。通过在验证队列5中进行mIHC染色,CD161+ CD8+ T细胞在RT中比在LT中特异性富集(图5I-J)。发现RT中CD161 CD8 T细胞的富集与较低的OS率显著相关(图5K),并且与肿瘤晚期相关(图5L)。
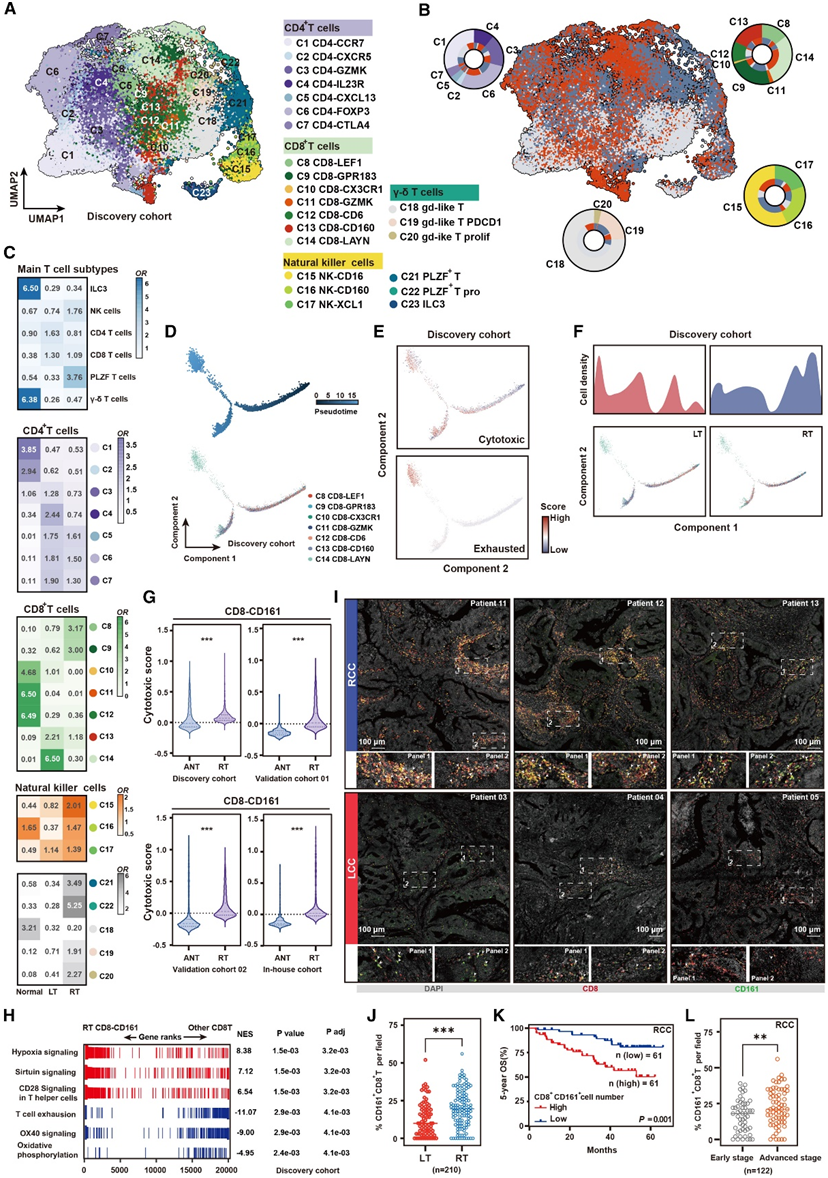
图5. T 细胞状态的表征。
(A)T 细胞的 UMAP 图。(B)按样本类型(正常、LT 或 RT)着色的 UMAP 图。(C) 热图显示每个组织中出现的 T 细胞亚群的 OR。(D)CD8 + T 细胞的伪时间分析。(E) 2D 伪时间图显示 LT 和 RT 样本中 CD8 + T 细胞的细胞毒性或耗竭信号的动态。(F) LT(左图)和 RT(右图)样本中伪时间排序的 CD8 + T 细胞的 2D 图。(G) 小提琴图显示来自邻近正常组织和RT样本的 CD8-CD161 细胞的细胞毒性评分。(H)与 RT 中的其他 T 亚群相比,CD8-CD161 细胞中的基因组的 GSEA 富集。(I)LT 和 RT 中 CD161 + CD8 + T 细胞的代表性 mIHC 染色(验证队列 5)。(J)根据 mIHC 染色结果显示左侧和右侧结肠肿瘤中CD161 + CD8 + T 细胞的密度的散点图。(K) 按 CD161 + CD8 + T 细胞浸润分层的 RCC 患者的 Kaplan-Meier OS 曲线。(L)早期和晚期 RCC 患者肿瘤切片中CD161 + CD8 + T 细胞的密度。
06
LCC 和 RCC 的 TME 中存在不同的免疫抑制模式
利用 CellChat,他们分析了来自 scRNA-seq 数据集的信号,以可视化由配体-受体相互作用介导的细胞间通讯。总体而言,具有更高侧特异性元程序活性的恶性上皮与免疫细胞的通讯强度增加(图6A-B),并介导来自其他亚群的最多传出和传入事件,可能代表 TME 内的信号中心和分泌枢纽。左侧和右侧肿瘤相应细胞群的细胞特征评分分别突出了位于同一点并包裹在恶性上皮细胞周围的聚类(图6C)。在RT中,他们观察到IShigh恶性上皮在靠近乏氧肿瘤核心的地方富集。进一步检测发现,这些区域中有许多与Mph-SPP1细胞和CD8-CD161 T细胞共定位(图6C),这表明这些细胞群之间可能存在活跃的细胞通讯。在肝移植中,PShigh恶性上皮细胞富集于糖酵解程度高的细胞区域,并与Mph-PLTP细胞、活化的Treg细胞和CD8-LAYN细胞共定位(图6C)。
在肝移植中,PShigh恶性上皮细胞与Mph-PLTP细胞之间的通讯强度较PSlow恶性上皮细胞与Mph-PLTP细胞之间的通讯强度明显增强(图6A)。Mph-PLTP细胞表现出脂肪酸氧化和氧化磷酸化信号增强,这是M2巨噬细胞的主要代谢特征。免疫荧光实验证实,LT中周围的PShigh恶性上皮显示了严重的Mph-PLTP细胞浸润(图6D)。PShigh恶性上皮细胞通过CSF1-CSF1R对调节Mph-PLTP细胞极化,并通过IL-13和TGF-β信号通路维持其抗炎表型(图6E)。NicheNet分析显示,Mph-PLTP细胞具有较高的EGF、WNT3A、VEGFA和IGF2配体活性和相对较高的基因表达。这些配体与PShigh恶性上皮细胞上相应受体结合所调控的靶基因是PS表达元程序的重要组成部分(图6E-F)。
在RT中,分别发现IShigh恶性上皮与Mph-SPP1细胞或CD8-CD161细胞之间的相互作用增加(图6B)。由于这些细胞群与缺氧诱导通路密切相关,他们提出在肿瘤的缺氧区域存在一个局部的IShigh恶性上皮网络,连接Mph-SPP1细胞和CD8-CD161细胞。这一网络可能协同增强免疫抑制。在RT中,空间转录组分析(ST-seq)和mIHC染色显示这些细胞群在乏氧肿瘤组织中非常接近,提示它们之间可能存在相互作用(图6G-H)。在细胞因子受体对中,IShigh恶性上皮通过CCL15-CCR1轴募集Mph-SPP1细胞。此外,IShigh恶性上皮通过程序性死亡配体1 (PD-L1)-CD80与Mph-SPP1细胞相互作用(图6I)。mIHC染色进一步证明CD80+ SPP1+巨噬细胞存在于PD-L1+高恶性上皮附近,证明它们之间的相互作用(图6J)。
CD8-CD161细胞处于记忆状态,细胞毒性弱,具有免疫抑制表型。CellChat分析再次证实,IShigh恶性上皮通过CXCL16-CXCR6对增强了CD8-CD161细胞的募集,并通过PD-L1轴与它们密切相互作用(图6I)。此外,Mph-SPP1细胞通过SPP1-CD44轴降低CD8-CD161细胞的细胞毒性能力(图6I)。
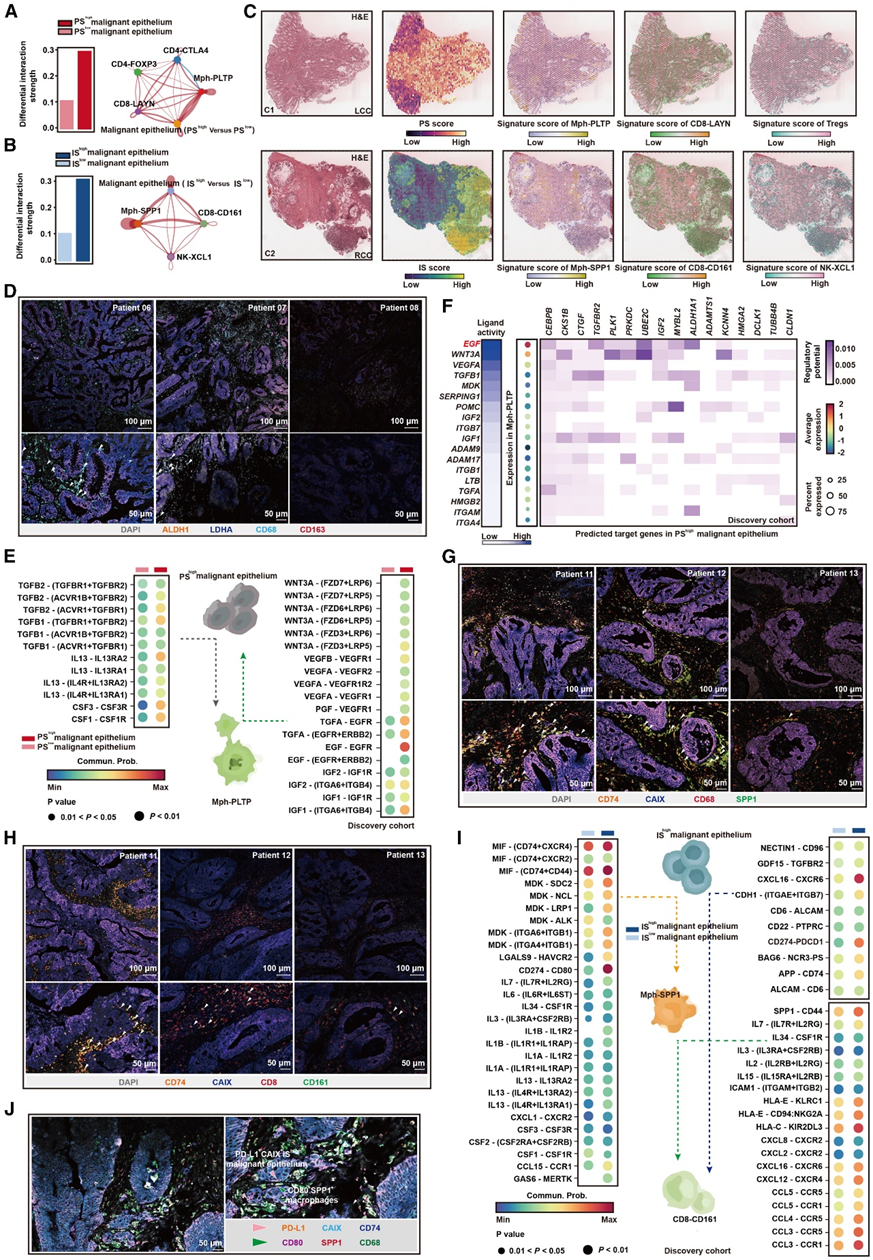
图6. 左侧和右侧结肠癌微环境中细胞间相互作用模式不同。
(A) 条形图分别显示 PShigh和 PSlow恶性上皮与 LT 中的免疫细胞之间的通信强度。(B) 条形图分别显示 IShigh和 ISlow恶性上皮与 RT 中的免疫细胞之间的通信强度。(C) 细胞空间特征图。(D) LT 中ALDH1 +恶性上皮和 CD163 +巨噬细胞的代表性 mIHC 染色。(E) 与 PSlow恶性上皮相比,LT 中PShigh恶性上皮与 Mph-PLTP 细胞之间的信号配体-受体对上调。(F) 根据 NicheNet 推断出 Mph-PLTP 细胞调节 PShigh恶性上皮的配体。(G-H) mIHC 染色的代表性图像。(I) RT 中 IShigh恶性上皮与 Mph-SPP1 细胞和 CD8-CD161 细胞之间的信号配体-受体对上调,而 ISlow恶性上皮则相反。(J) 代表性 mIHC 染色。
07
ICI处理可能对RCC更有效
免疫检查点抑制剂(ICI)治疗(PD-1/PD-L1阻断)可能是肾癌更合理的选择。接下来,他们分析了之前研究的scRNA-seq数据,该研究旨在揭示结直肠癌患者对ICI治疗耐药和敏感的基础。从本研究中提取结肠癌患者的scRNA-seq数据和治疗相关信息,组成ICI治疗队列(图7A)。对所有样本进行的聚类分析显示了代表8种细胞类型的聚类(图7B)。他们比较了实现病理完全缓解(pCR)的患者暴露于ICI前后的细胞类型分布和功能变化。在治疗前的标本中,来自RT的恶性上皮高度表达IS元程序(图7C)。在pCR反应中,ICI治疗诱导了高恶性上皮比例的显著降低,这一点通过mIHC染色得到了证实(图7C)。
通过比较巨噬细胞的+ICI/pCR组和- ICI组,他们发现上调的基因与内吞作用、吞噬作用、抗原受体介导的信号通路和T细胞活化相关(图7D)。ICI治疗后+ICI/pCR组RCC患者的Mph-SPP1细胞比例明显下降(图7E)。在ICI对总体CD8+ T细胞转录组的影响方面,细胞毒性、耗竭和增殖基因特征评分均与ICI治疗相关;与-ICI组相比,细胞毒性特征在+ICI/pCR组中富集,而耗尽和增殖的特征在+ICI/非pCR组中富集(图7G)。有趣的是,ICI不仅增加了CD8-CD161细胞的比例(图7H),而且增加了CD8-CD161细胞的抗原呈递过程和特化IFN-γ应答基因(HLA-DQA1、HLA-DRA和CD74)的表达(图7I)。此外,ICI治疗后,CD8-CD161细胞增加了CD8-GZMK细胞的特征,减少了CD8-LAYN细胞的特征,这与轨迹分析的结果一致(图7J)。这些发现表明,ICI可能促进CD8-CD161细胞向CD8-GZMK细胞的转化,阻断CD8-CD161细胞向CD8-LAYN细胞的转化,从而增强RT的抗肿瘤免疫。
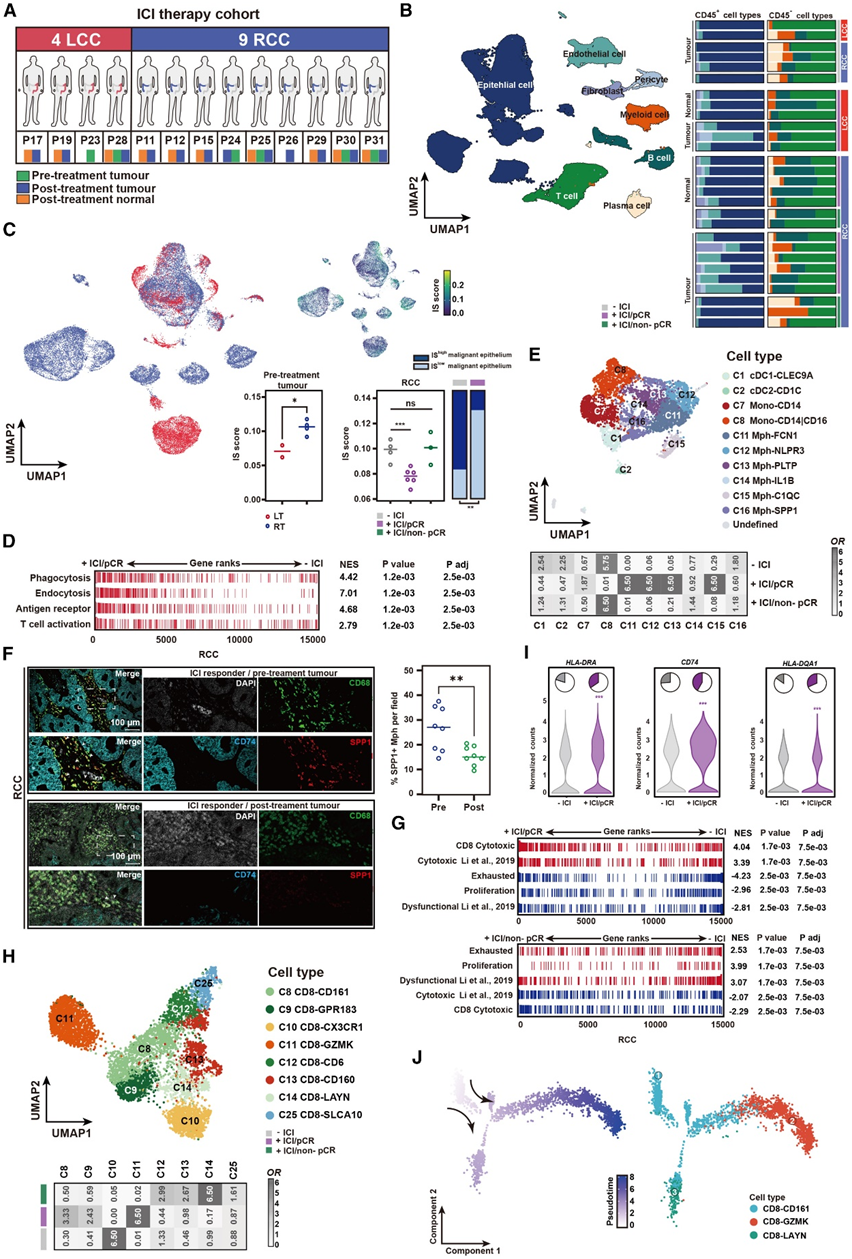
图7. 恶性上皮中 IS 元程序的消退与免疫检查点抑制剂治疗相关。
(A)结肠癌患者的治疗相关信息。(B)所有样本中广泛细胞类型的 UMAP 图(左)。细胞聚类频率显示为每个样本中总细胞的比例(右)。(C) UMAP 图按颜色显示恶性上皮起源,以及恶性上皮中 IS 评分的表达。(D) 所有巨噬细胞中基因集的 GSEA 富集比较。(E) 髓系细胞的 UMAP 图和热图显示每组中出现的细胞亚群的 OR。(F) mIHC 染色的代表性图像。(G) 所有 CD8 + T 细胞中基因集的 GSEA 富集。(H) CD8 + T 细胞的 UMAP 图和热图显示每组中出现的细胞亚群的 OR。(I)scRNA-seq 数据集中HLA-DRA、CD74和HLA-DQA1的表达。(J) RT 样本中CD8+ T 细胞聚类的伪时间排序分析。
+ + + + + + + + + + +
结 论
本项研究结合来自 105 名结肠癌患者的scRNA-seq以及空间转录组学和大规模组织学分析,描绘了两个特定于一侧的表达元程序来捕获左侧和右侧恶性上皮之间常见的转录异质性模式。增殖干性元程序在与 Mph-PLTP 细胞、活化的调节性 T 细胞 (Treg) 和耗竭的 CD8-LAYN 细胞共定位的左侧恶性上皮中明显富集,构成葡萄糖代谢重编程生态位。IS元程序在右侧恶性上皮中表现出特异性富集,尤其是在吸烟的右侧结肠癌患者中。 IShigh恶性上皮在空间上定位于缺氧区域,通过减弱 Mph-SPP1 细胞抗原呈递和招募先天性细胞毒性降低的 CD8-CD161 细胞来促进免疫逃避。
+ + + + +



