

 English
English文献解读|Nat Metab(20.8):大鼠皮下白色脂肪组织的性别二态性和运动训练的多组学反应
✦ +
+
论文ID
原名:Sexual dimorphism and the multi-omic response to exercise training in rat subcutaneous white adipose tissue
译名:大鼠皮下白色脂肪组织的性别二态性和运动训练的多组学反应
期刊:Nature Metabolism
影响因子:20.8
发表时间:2024.05.01
DOI号:10.1038/s42255-023-00959-9
背 景
皮下白色脂肪组织(scWAT)是一种动态储存和分泌器官,可调节全身稳态,但耐力运动训练(ExT)和性对其分子景观的影响尚未完全确定。
实验设计
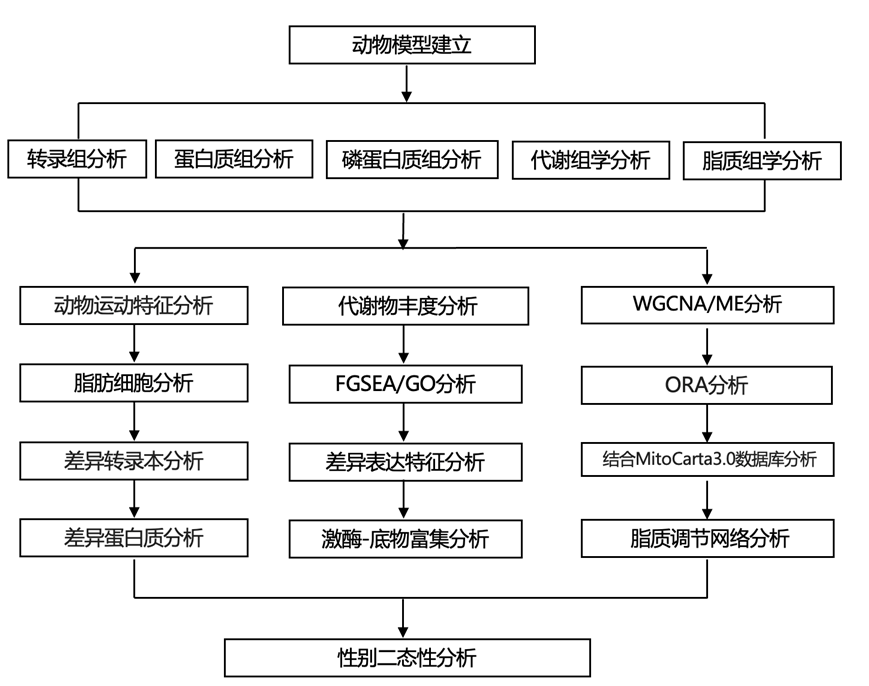
结 果
01

耐力训练的性别二态性表型反应
在本项研究中,从 6 个月大开始,雄性和雌性 F344 大鼠接受渐进式 ExT 计划,包括每周连续五天在跑步机上跑步,持续 1、2、4 或 8 周,目标强度为最大氧耗率的 70-75%消耗(VO2 max)(图1a)。不运动对照 (SED) 大鼠与训练 8 周的组大鼠进行年龄和性别匹配。在用于多组学分析的动物中,在8周的ExT后,两种性别的VO2max(相对于瘦体重或总体重)增加相似,而在8周的静止后,SED大鼠的VO2max相对于总体重显著降低(图1b)。8周后,雄性大鼠的体重和全身脂肪下降(图1c)。在雌性大鼠中,脂肪量在训练4周后下降,但在8周时恢复到训练前的水平(图1c)。在同一时间段内,SED大鼠的雄性和雌性脂肪量都增加了(图1c)。因此,8周的ExT减轻了雌性大鼠脂肪量的增加,减少了雄性大鼠脂肪量。考虑到身体成分的变化,他们在ExT后检测了scWAT脂肪细胞的大小(图1d)。对近56000个总脂肪细胞的分析表明,训练后,两种性别的大小分布存在差异(图1e)。值得注意的是,4周训练后,雌性大鼠的scWAT中较小的脂肪细胞(直径20 μm)显著增加,而较大的脂肪细胞(直径≥45 μm)相应减少,这与雌性大鼠的脂肪量在4周时间点的减少一致(图1d)。进一步与8周训练后的脂肪量变化一致,脂肪细胞大小分布模式与SED对照组相似,这可能反映了雌性大鼠在长时间训练后保留腰腹脂肪储存的代偿反应(图1e)。相反,与SED相比,8周训练的雄性大鼠的scWAT显示小脂肪细胞的比例显著较高,而大脂肪细胞的比例显著较低(图1e),表明训练诱导的scWAT脂肪细胞大小减少。因此,尽管训练诱导的心肺功能改善在两种性别中相似,但在整体身体成分和scWAT特异性脂肪细胞形态中均出现了性别二态性ExT反应。在选择进行多组学分析的大鼠中,与SED相比,1周和2周训练的雄性大鼠血浆甘油含量较高,1周和2周训练的雌性大鼠血浆甘油含量较低(图1f)。雄性大鼠的非酯化脂肪酸(NEFA)在训练的早期阶段有升高的趋势,但在第8周时下降,这表明在早期训练反应中,scWAT的三酰甘油(TAG)发生动员,但随着训练时间的延长,总脂肪储备耗尽,这种动员不能持续。与这一结果相一致的是,与雌性大鼠相比,雄性大鼠的瘦素水平更高,并且在与减脂呈正相关的训练中显示出更稳健的下降(图1g)。在4周训练的雄性中,胰高血糖素降低,血糖升高,血浆胰岛素水平没有变化(图1h-i)。相反,在雄性大鼠中,这些变量都不受ExT的影响。总体而言,研究结果强调了在对训练的系统性和scWAT特异性生理适应方面的性别差异,其中一些指标可作为雄性脂肪动员增强和雌性脂肪储存中脂质保留的指标。在血糖、胰高血糖素和胰岛素水平方面,性别差异也很明显,雌性的血浆胰岛素水平总体较低(图1h-i)。
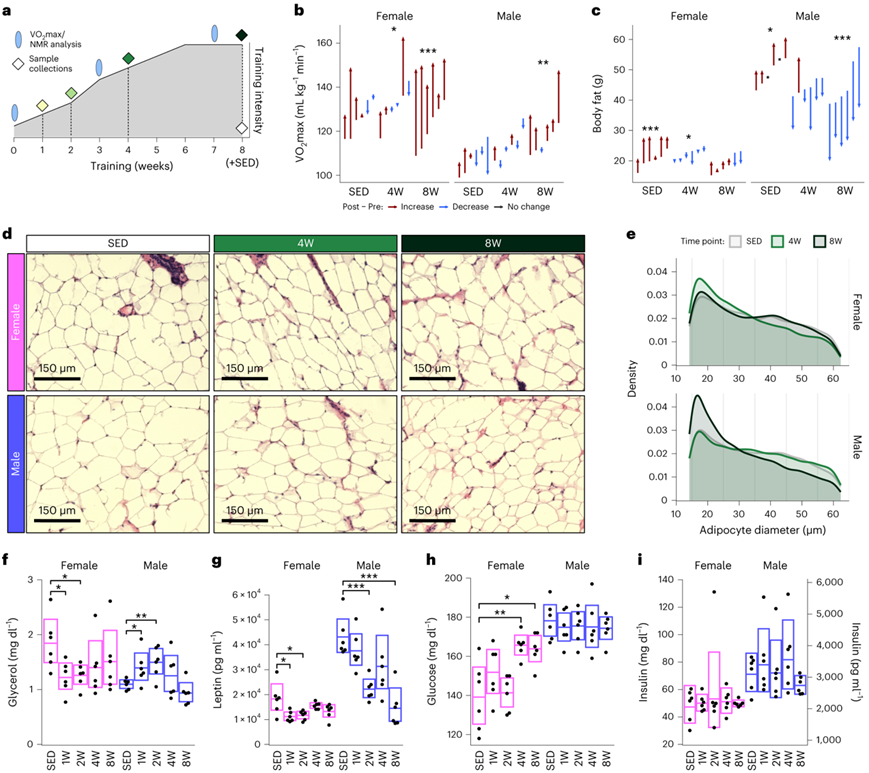
图1. 耐力训练的性别二态性表型反应。
(a) 训练方案图。(b) 在SED、4W和8W时间点记录选择用于多组学分析的动物的训练前和训练后的相对VO2最大值。 (c) SED、4W和8W训练的动物训练前和训练后的总脂肪量。(d) 来自不运动、4W 和 8W 雌性和雄性大鼠的 scWAT 的代表性组织学图像。(e) 组织学切片的脂肪细胞直径分布。(f-i) 大鼠的甘油、瘦素、葡萄糖和胰岛素水平。
02
不运动大鼠 scWAT 显示分子性别二态性
考虑到 ExT 的性别差异表型反应,他们对选择进行多组学分析的大鼠进行了更彻底的表征。在所有组中的分子丰度中都观察到了显著的性别差异;具体来说,10336 个转录本、4226 个蛋白质、6028 个磷酸位点和 615 个代谢物在 雄性和雌性大鼠之间显示出统计显著差异(图2a),占每个数据集中所有量化特征的 20-60%。SED雄性大鼠scWAT中升高的最高转录本包括与脂质代谢过程(Car2和Acss2)和不饱和脂肪酸(FA)合成(Fads2和Fads1)相关的转录本(图2a)。在SED雌性大鼠中表现出较高表达的转录本包括参与生长因子/Akt信号传导(Igf1r、Dapk1和Kit)、脂肪形成(Adipoq)和葡萄糖摄取(Slc2a4)的分子(图2a)。在蛋白质组水平上,雄性中最重要的差异蛋白与脂质代谢(Acot1、Hadh和Acbd7)、胆固醇转运(Npc2)和生酮(Hmgcl)相关(图2a)。 SED 雌性大鼠的 scWAT 显示出与核和细胞结构完整性相关的蛋白质(Lmnb1、Col14a1 和 Svil)丰度较高(图2a)。与转录组数据一致,雌性scWAT 中 Adipoq 和 Slc2a4 (Glut4) 蛋白丰度较高。相对于雌性大鼠,雄性大鼠抗氧化剂 Sod1 在mRNA 和蛋白质水平上均升高(图2a)。
在这些差异分析的基础上,他们进行了快速基因集富集分析(FGSEA),以检测基因本体(GO)(BP,生物过程;MF,分子功能;CC,细胞成分)的富集。他们还评估了 SED 雄性和雌性scWAT 的转录组和蛋白质组谱之间的通路水平一致性(图2b)。与氧化代谢、线粒体复合物组装、支链氨基酸(BCAA)分解代谢、蛋白酶体激活和核糖体亚基相关的通路在转录组和蛋白质组水平上均在雄性中持续富集(图2c)。在雌性中,包括板状足和肌动蛋白丝束在内的通路在两个组中均富集(图2c)。在探索具有性别差异的蛋白质组学网络时,SED雄性表现出与有氧代谢、多囊泡运输(包括囊泡泛素化依赖的分解代谢和分选)和翻译调节相关的通路的富集(图2d)。在蛋白质组水平上,与mRNA核调控、mRNA剪接和mRNA降解以及非编码RNA调控过程(5.8S RNA成熟和小核糖体RNA代谢)相关的网络表达上调;在转录组水平上,与免疫和发育相关的网络表达上调,如免疫效应过程、细胞因子产生和轴突的正向调节、血管和骨骼肌发育相关的网络表达上调(图2d)。
RefMet代谢产物化学亚类的FGSEA也揭示了性别双态性,在SED雄性中富集了氨基酸和酰基辅酶A。雌性大鼠scWAT显示了TAG的富集,主要由几个长链(>40个碳)TAG化合物驱动。其中一些代谢组学差异与雄性转录组和蛋白质组水平的线粒体和氨基酸代谢通路富集一致。他们在转录组,蛋白质组和代谢组水平上观察到了SED雄性和雌性F344大鼠的scWAT的显著差异。
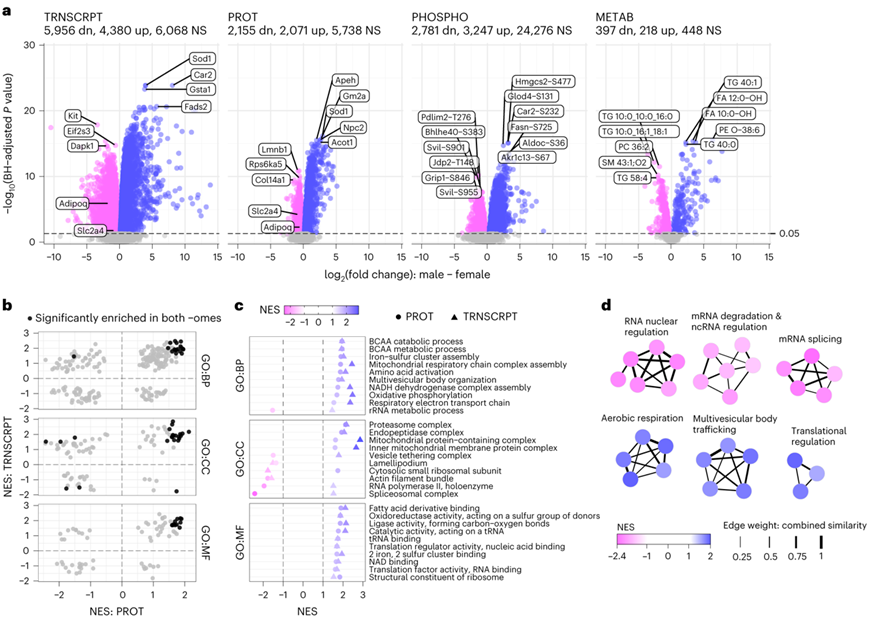
图2. 大鼠 scWAT 分子景观中的性别二态性。
(a) 火山图显示了比较雄性和雌性 SED 的差异分析结果中转录本 (TRNSCRPT)、蛋白质 (PROT)、磷酸位点 (PHOSPHO) 和代谢物 (METAB) 变化的幅度和显著性。(b) 在蛋白质组学和转录组学中通过 GO 分离进行检测的基因集的标准化富集分数 (NES) 的散点图。(c) GO分析。(d) 显著富集的通路的聚类。
03
性别特异性多组学 scWAT 对 ExT 的适应
接下来,他们分析了样本水平的数据集,以确定每个训练时间点的性别差异表达特征(例如,1周训练的雌性大鼠vs SED雌性大鼠)(图3a-b)。总体而言,雌性大鼠scWAT在所有四个ExT时间点具有相似数量的差异组学特征,而雄性大鼠的差异表达特征的数量随着训练时间的延长而增加(图3a-b)。
在转录组学分析中,在训练 1 周和 8 周的雌性(分别为 167 个和 193 个转录本)和训练 2 周的雄性(529 个转录本)中观察到最大数量的差异转录本(图3a-b)。六个转录本在雌性的所有训练时间点都显示出差异表达。值得注意的是,Grb14(胰岛素信号传导的负调节因子)随 ExT 下调,而Olah[一种中链 (MC) FA 硫酯酶]和Hmgcs2(一种具有脂肪生成作用的生酮酶)在雌性大鼠的所有训练时间点均有所增加。在雄性大鼠中,唯一在所有训练时间点表现出差异表达的转录本是缺氧诱导的碳酸酐酶 (Ca12),该酶的表达下调。FGSEA 进一步强调了 ExT 进行性转录组反应中的性别差异(图3c)。ExT 1周和2周后,雄性scWAT中与免疫受体活性以及细胞因子、生长因子、细胞外基质、整合素和胶原蛋白结合相关的通路正富集(图3c)。这些通路的富集是由参与组织应激(Hif1a、Tlr4、C4b和Nlrp3)、组织重塑(Itgb1、Timp3、Lifr、Pten和Fgfr1)和血管生成(Kdr、Flt1、Pdgfb和Vegfa)的基因驱动的(图3c)。2 周后,GO-MF 细胞因子结合和免疫受体活性途径的富集是由与辅助 T 17 (TH17 ) 细胞激活(Cd4、Il17ra、Tgfbr2、Il6st和Ctsl )相关的转录本驱动的。雌性表现出与转录/翻译调节相关的通路的早期富集,通常在 1 周时达到峰值,并在整个训练过程中保持富集(图3c)。与氧化磷酸化(OXPHOS)活性相关的通路在1周时两性都有所下降,但在2周和4周时雄性和雌性大鼠中恢复(图3c)。在8周训练的大鼠中,与适应性免疫系统、免疫受体活性、嗜天青颗粒和空泡腔相关的通路在雄性大鼠中富集,而与转录和翻译调节过程、有氧呼吸和嗜天青颗粒相关的通路在雌性大鼠中富集(图3c)。
在蛋白组学分析中,雄性大鼠的 scWAT 蛋白质组发生了剧烈变化,随着训练时间的延长而增加(8 周时有 654 个差异表达蛋白),而雌性大鼠的反应要小得多,大多数差异发生在 2 周后(221 个差异蛋白)(图3a-b)。在雄性中,18种蛋白质在所有时间点都有差异丰度,其中大多数与线粒体功能(Maip、Atp6v0d1和Immt)、线粒体溶质转运(Slc25a11、Slc25a3和Slc25a15)和囊泡转运(Tmed2和Sec61a1)有关。 Lrpprc 是一种调节线粒体基因转录以促进脂质氧化的蛋白质,其在雄性训练过程中有所增加。Slc25a15 是一种促进精氨酸合成的线粒体鸟氨酸载体,在雄性 scWAT 的训练过程中也有所升高。精氨酸部分通过 Ampk 激活促进脂肪分解。FGSEA 显示,从 ExT 2 周开始,雄性 scWAT 中与 OXPHOS 和线粒体特异性核糖体生物发生相关的通路急剧增加(图3d)。在整个训练过程中,雄性还表现出与囊泡运输、核糖体活性和生物发生相关的通路的富集(图3d)。雌性在所有四个时间点都表现出五种蛋白质的丰度差异。4或8周的训练增加了雌性抗炎和胰岛素增敏Orm1的表达,提示训练在没有减脂的情况下改善瘦雌性scWAT胰岛素信号传导的候选机制。与雄性大鼠相比,雌性大鼠scWAT在第1周和第2周时线粒体相关通路呈负性富集,在第8周时线粒体相关通路略有富集。在8周时,两种性别均显示出与转录和翻译相关的通路的富集,雄性大鼠的蛋白质组学反应比转录组学反应更强(图3c-d)。
在磷酸化蛋白组学分析中,数百个蛋白磷酸化位点受到ExT的差异调节,其两性二态的时间模式与蛋白质组学的ExT反应相似。总的来说,雄性动物在4周和8周的外泌体治疗后表现出更强的磷酸化蛋白质组学反应(图3a-b)。雌性大鼠在所有4个训练时间点均可调节14种磷蛋白,包括与炎症反应(Pde4 a - b和Lrrfip1-S85)和Camk2b-T398(脂肪细胞胰岛素信号的负调控因子)相关蛋白的磷酸化水平降低。雄性小鼠在整个ExT中表现出一致的15种蛋白磷酸化的调节,包括调节囊泡转运的蛋白(Htt-S621, Klc1-S521/S524和Uhrf1bp1l-S418)和自噬的蛋白(Bnip3-T66和Ulf1-S458)的磷酸化增加。为了根据已知底物磷酸化的变化来推断激酶活性,他们利用来自PhosphoSitePlus (PSP)的激酶-底物关系数据进行了激酶-底物富集分析(KSEA)(图3e)。该分析的结果显示,在2周训练的雄性小鼠中,MAP3K8、LRRK2和MYLK激酶的活性降低(图3e),而在4周和8周训练的雄性小鼠中,脂解诱导激酶PRKAA1 (AMPK1ɑ1)的活性增加。雄性DYRK2活性在8周时降低(图3e),这主要是由CARHSP1磷酸化(S30、S32和S41)的变化驱动。在雄性中,AKT1活性在4周时增加,到8周时随着MAPK1和mTOR活性的增加而减弱。
鉴于mTORC1在合成代谢信号传导(包括在脂肪组织中)中的明确作用,加上在雌性大鼠scWAT中观察到与胰岛素信号传导抑制相关的蛋白和磷蛋白(例如Grb14和Camk2b)的下调,他们试图确定表明mTOR活性富集的磷蛋白的特征。胰岛素依赖性RAGC-T394磷酸化、RPS6-S236、EIF4EBP1(S65和T70)、AKT1S1 (PRAS40) (S183)和MAF1(S68和S75)的磷酸化支持mTOR活性的增加(图3f)。在4周训练的雌性大鼠中,驱动AKT1富集的磷酸位点包括分解代谢FOXO3的AKT依赖性失活位点(S253)和FA合成蛋白ATP柠檬酸裂合酶(ACLY)激活位点附近的两个位点(S425和T447)。在脂肪细胞中,ACLY促进Glut4的表达和脂肪新生,这种方式在雌性大鼠中更为明显。结合多组学分析结果,这些数据表明合成代谢胰岛素/AKT/mTORC1的富集是促进雌性scWAT胰岛素敏感性和脂质储存/再循环的潜在机制。
他们采用了一整套代谢组学技术来通过 ExT 表征 scWAT 代谢组/脂质组的变化。这包括非靶向方法,以及专注于化学相关分析物组(例如氨基酸、酰基肉碱和核苷酸)的定量靶向方法。总体而言,scWAT代谢组在两性中表现出最小的性别二态性ExT反应,差异调节最多的代谢物出现在两性的第 2 周和第 8 周(第 2 周,雌性和雄性大鼠分别有 135 和 161 个差异代谢物;第 8 周,则分别有139 和 211 个差异代谢物)(图3a-b)。4周训练时间点显示了最显著的性别差异,在雄性大鼠中有154种差异调节的代谢物,而在雌性大鼠中只有20种。在两种性别中,RefMet代谢亚类的FGSEA显示,在训练1周的大鼠中,磷脂酰胆碱(PC)、醚磷脂酰胆碱(O-PC)和酰基肉碱物种呈正富集,而在8周时,TAG呈负富集(图3g)。雌性的PC和O-PC磷脂物种的早期富集时间延长,鞘磷脂(SM)和不饱和脂肪酸在1周和2周时呈负富集。在所有4个ExT时间点,雄性小鼠在ExT和PC的1、4和8周均显示O-PC、二酰基甘油(DAG)和溶血磷脂酰胆碱(LPC)呈正富集(图3g)。另外,雄性小鼠在第4周时表现出磷脂酰乙醇胺(PE)的增加,到第8周时下降(图3g)。值得注意的是,在 SED 雄性大鼠 scWAT 中升高的酰基辅酶 A 在训练 4 周的雄性中下降(图3g,图S4a),而氨基酸在 8 周时急剧增加,进一步支持脂质利用率的增加以及雄性大鼠中 ExT 向氨基酸代谢的转变。这一结果也与8周时雄性中与scWAT脂肪酰基CoA丰度对应的单核苷酸和二核苷酸磷酸激增,以及核苷酸三磷酸减少和AMP:ATP比值增加一致(图S4c)。相反,雌性在第1、2和8周表现出氨基酸的正富集(图3g,图S4b)。

图3. 性别特异性多组学 scWAT 对 ExT 的适应。
(a-b) 转录本、蛋白质、磷酸位点和代谢物的 UpSet 图。(c-d) 在转录组学或蛋白质组学FGSEA 结果的八个比较中的任何一个中最显著富集。(e) 从KSEA的磷酸化蛋白质组学差异分析结果中推断出,每个训练组与性别匹配的对照组的指示激酶活性。(f) 磷酸化位点(粉红色)在8w训练的雌性中驱动mTOR富集的示意图。 (g) 根据代谢组学FGSEA结果,RefMet化学亚类在8个比较中至少一个显著富集。
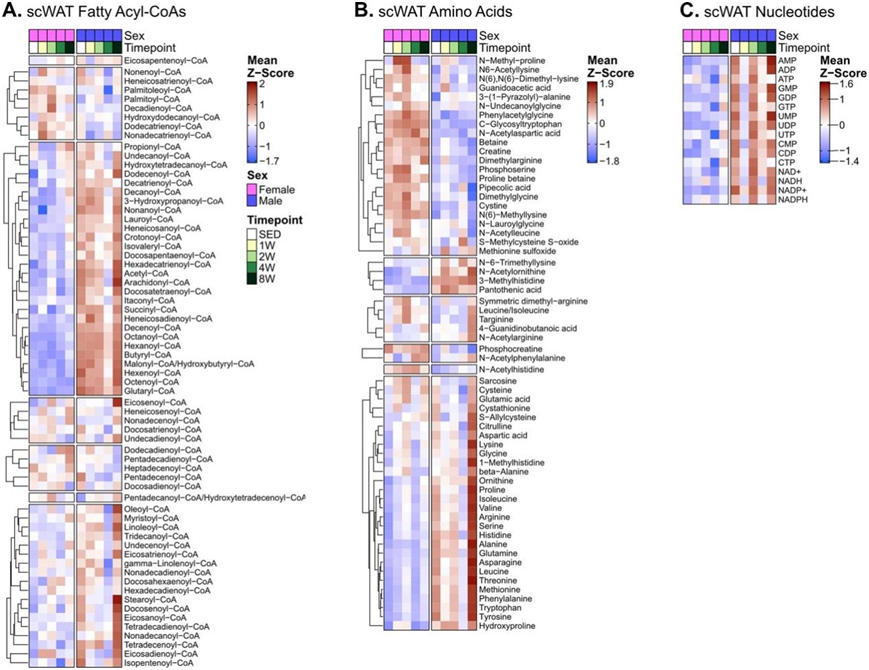
图S4. 代谢组学可视化。
(a–c) scWAT 代谢组热图显示酰基辅酶A、氨基酸和核苷酸。
04
对 ExT 的综合表型组学反应
为了整合组学数据集,他们进行了加权基因共表达网络分析(WGCNA),并生成代谢组学/脂质组学、蛋白质组学和转录组学数据集的模块(高度相关的特征组)。然后,他们评估了模块表观特征(ME)(每个模块的主特征向量)与选定的临床血浆分析物和其他表型标记物的相关性(图4a),以及代谢组学和其他组学ME之间的相关性(图4b) 。然后,进行过表征分析(ORA)来测试每个模块中通路或RefMet化学子类的定位。
在整个训练过程中,最大的代谢组学/脂质组学模块 M1 的 ME 较高。M1由长链(>48 个碳)TAG 过度代表(图4c)。除了M1,其他在雌性中具有较高ME的模块是M4(由SM和O-PE过度代表)和M5(由PC和O-PC物种过度代表)。M1, M4和M5模块ME与血糖呈负相关,M1和M4 ME与scWAT脂联素和脂肪量变化呈正相关(图4a)。这提示了这些模块与葡萄糖调节和脂质储存之间的关系。第二大模块M2由酰基CoA物种过度代表(图4c);其代谢能在雄性中较高,并与scWAT脂联素和体脂变化(减脂)呈负相关(图4a)。M3主要含有38-40个碳PC(图4c),雄性的ME在8周时增加。M3和含氨基酸的M6模块的ME与每个视野的scWAT脂肪细胞计数(缩小的脂肪细胞)呈正相关,与体脂变化呈负相关(图4a-c),提示与脂肪细胞的脂质动员和利用相关。
T1 是最大的转录组学模块,其 ME 在雄性中最高,并且随着雌性大鼠训练的进行而略有增加。T1 中与囊泡输出和回收过程相关的通路过多,包括自噬体和蛋白酶(图4d)。T1 还包含与线粒体复合体组装和 FA 分解代谢过程相关的通路;其 ME 与 TAG 定位 M1 代谢组学模块的 ME 呈负相关(图4b)。雄性大鼠中的 T7 ME 也较高,并且该模块由有氧呼吸过度代表(图4d)。该ME与TAG定位M1模块呈负相关,与含有酰基辅酶A的M2 ME的ME呈正相关(图4b)。T2-T4 的 ME 在雌性中较高,并且与 scWAT 脂联素蛋白丰度相关(图4a)。T2和T3由与发育过程相关的通路过度代表(图4d),包括脂肪形成祖细胞的标记(Cd34、Dpp4、Icam1和Pdgfrb),表明雌性scWAT中脂肪形成潜力升高或不同的前脂肪细胞表型。T5 ME 也与 TAG 定位 M1 的 ME 相关,并且在雌性中保持相对稳定,在雄性中先增加然后在 8 周时降低,这导致 8 周时雌性相对于雄性的 ME 丰度升高(图4b)。
最大的蛋白质组学模块 P1 的 ME 在雄性中要高得多,并且在两性中随着训练而增加。P1 以与线粒体有氧呼吸/OXPHOS 相关的通路为主(图4e)。P1 ME与脂肪和体重、含有TAG的M1 ME和scWAT脂联素水平呈负相关(图4a-b),再次证明脂质氧化与scWAT TAG丰度之间的负相关。值得注意的是,P1 还含有与过氧化物酶体束缚和生物发生相关的蛋白质(Pex11a、Pex13 和 Pex19)、蛋白酶体蛋白和先天免疫系统激活,表明免疫信号传导对促进 α 和 β 氧化的作用在 scWAT 和 ExT 中。第二大模块 P2-P4 和 P6 的 ME 在雌性中更为丰富;他们的ME与体脂和scWAT脂联素水平的变化相关(P2-P3),与血浆葡萄糖、胰岛素和瘦素(P2-P4)呈负相关(图4a)。 P6模块包含与脂质合成相关的蛋白质(Gpat3、Dgat2、Acsl1、Elovl1、Agpat2-4和Mboat7)和胰岛素调节的葡萄糖转运蛋白GLUT4(Slc2a4),它们与含有TAG的M1 ME呈正相关,与含有TAG的M1 ME呈负相关。总之,这些结果揭示了 scWAT 中介导对 ExT 的性二态性反应的分子中心。

图4. ExT 的综合表型组学反应。
(a) ME 和临床测量之间 Spearman 相关性的热图。 (b) 代谢组学 ME 和其他组的 ME 之间 Spearman 相关性的热图。(c) 每个代谢组学 WGCNA 模块中所有过度代表的 RefMet 化学子类。(d-e) 每个转录组学或蛋白质组学WGCNA 模块中出现次数最多的 GO-BP。
05
脂质调节网络显示 ExT 的性别二态性
利用MitoCarta3.0数据库中的FGSEA可以更深入地揭示了性别对scWAT线粒体代谢的影响。与FGSEA结果一致,与SED雌性相比,SED雄性显示出MitoCarta通路的强富集(图2c,图5a),包括参与FA氧化、碳水化合物和支链氨基酸代谢的蛋白质丰度增加。通过训练,雄性显示出分解代谢通路(如支链氨基酸和碳水化合物代谢和FA氧化)的稳健富集(图5b)。与此相反,雌性则显示出更丰富的线粒体核糖体和翻译通路,显示出Fe-S聚类生物合成和解毒方面的独特增加(图5b)。尽管两种性别均有线粒体代谢途径的变化,但通过线粒体DNA含量、映射到线粒体基因的RNA测序(RNA-seq)读数和心磷脂CL(72:8)丰度评估的线粒体含量均不受训练的影响(图5c)。这表明scWAT的线粒体丰度并未因ExT而改变。
对调节 scWAT 脂质回收(再酯化和脂解)的蛋白质的具体分析揭示了长链酰基辅酶 A 合成酶 (Acsl) 亚型的性别二态性,长链酰基辅酶 A 合成酶 (Acsl) 亚型是调节脂肪酰基辅酶 A 代谢分配的酶家族。雌性主要WAT亚型Acsl1的表达水平高于雄性(图5d)。3-磷酸甘油酰基转移酶(Gpat)亚型(甘油脂合成的限速酶)的丰度也表现出性别二态性,在所有时间点,与雄性相比,雌性中的Gpat3较高,而雄性中的Gpat4较高(图5d)。甘油二酯酰基转移酶 (Dgat) 和磷酸烯醇丙酮酸羧激酶 (Pck) 也是关键的再酯化酶。与脂肪量保存一致,雌性在所有时间点都表现出较高水平的 Dgat1 和 Pck2(图5d)。脂质合成蛋白 Acaca (Acc1) 在 SED 雄性 scWAT 中升高,并且在雌性中随着训练 8 周而升高(图5d)。
最后,他们检测了 scWAT TAG 丰度和脂质代谢的其他代谢组标记物的变化。与脂肪量、血浆瘦素和脂肪细胞大小分布的减少一致,ExT 8周后,雄性的中位总 scWAT TAG 丰度减少了约 70%(图5e)。其特征是,训练 4 周和 8 周的雌性中短链 (SC) 和 MC TAG 丰度增加,而长链 (LC) TAG 种类(> 40 个碳)在 8 周时减少。在雄性 ExT 8 周时,SC 种类的丰度与 SED 没有不同,LC TAG 物种减少,但是程度低于雌性(图5f)。雌性在所有训练时间点都有较高水平的几乎所有种类的MC和LC(>8碳)酰基肉碱,而雄性有较高水平的SC酰基肉碱,代表FA完全氧化的产物,除了C2(乙酰基)(图5f)。雌性在第 1 周时还表现出 MC 和 LC 酰基肉碱的强劲增加,表明 FA 氧化减少,与 OXPHOS 蛋白水平降低一致。这些特征与雄性中 FA 氧化的激活消耗 LC 酰基肉碱并产生 SC 酰基肉碱以及雌性中 LC 物种的保存一致,可能用于回收到 TAG 进行储存。总之,scWAT 脂质代谢物的变化表明雄性的脂质利用和脂肪储存的消耗与雌性的脂质回收以保存脂肪量。
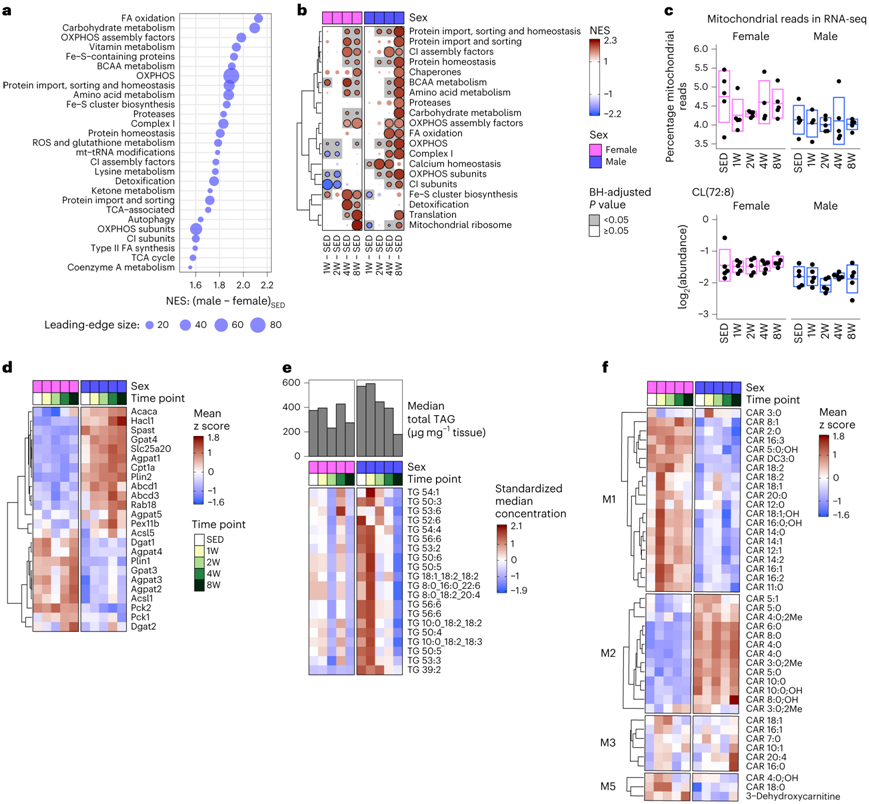
图5. 综合组学揭示了 scWAT 线粒体代谢和 ExT 脂质回收中的性别二态性。
(a) 来自蛋白质组学SED雄性与SED雌性 FGSEA 结果。(b) 在蛋白质组学 FGSEA 结果中的八个经过训练的对照与不运动对照组的比较中,最显著富集的 MitoCarta 的热图。 (c) 线粒体基因读数百分比和心磷脂 CL(72:8) 的水平。(d) 参与脂质代谢的蛋白质。(e) 雄性和雌性大鼠的scWAT样品中每个时间点的中位总TAG浓度的条形图。(f)根据代谢组学 WGCNA 模块分组的酰基肉碱种类的热图。
+ + + + + + + + + + +
结 论
本项研究利用综合多组学方法,并利用体力活动分子传感器联盟 (MoTrPAC) 生成的数据,在SEC大鼠的 scWAT 以及该组织对 ExT 的动态反应中显示出深刻的性别二态性。具体来说,SEC雌性的 scWAT 显示出与胰岛素信号和脂肪生成相关的组学特征,而SEC雄性大鼠的 scWAT 在与有氧代谢相关特征的富集。这些性别特异性组学特征通过 ExT 得以保留或放大。总体而言,这项研究强调了性别对脂肪组织生物学的强大影响,并为研究 scWAT 对 ExT 的反应提供了新的理论依据。
+ + + + +



