

 English
English文献解读|Sci Adv(14.957):盘状结构域受体 2 调节异常的间充质谱系细胞命运和基质组织
✦ +
+
论文ID
原名:Discoidin domain receptor 2 regulates aberrant mesenchymal lineage cell fate and matrix organization
译名:盘状结构域受体 2 调节异常的间充质谱系细胞命运和基质组织
期刊:Science Advances
影响因子:14.957
发表时间:2022.12.21
DOI号:10.1126/sciadv.abq6152
背 景
间充质谱系 (MLin) 细胞从其局部环境中接收机械信号,并沿各种谱系分化,包括成骨细胞、软骨细胞和脂肪细胞。已知MLin细胞与细胞外基质 (ECM) 之间的相互作用会影响细胞增殖、迁移和细胞命运决定。异位骨化(HO)是一种发生在创伤性损伤后的病理情况,其中MLin细胞与ECM的相互作用出现了问题。HO 的特征是 MLin 细胞的异常骨软骨分化,导致在骨骼外软组织(包括肌腱或肌肉)中形成异位软骨和骨骼。在以往的研究中已发现由于受伤部位富含胶原蛋白的 ECM 结构发生变化,固定的患肢会抑制与创伤相关的肢体 HO。然而,在肌肉骨骼创伤的条件下,MLin 细胞与周围 ECM 相互作用的机制以及这如何改变 MLin 细胞命运尚不清楚。
实验设计

结 果
01

Mli细胞的表征
研究者团队建立了HO /腱切断术 (BT) 损伤的小鼠模型 (图1A), 在三个时间点(未受伤、受伤后 1 周和受伤后 3 周)从肌腱损伤部位采集细胞用于单细胞 RNA (scRNA) 测序分析(图1A-B)。通过差异基因表达分析在收获的细胞群中表征各种细胞群, 发现MLin 细胞的特征性表达Pdgfrα 和Prrx1(图1C),同时软骨形成分化标志物Acan和Sox9以及早期成骨标志物Runx2的表达增加并伴随着纤维状胶原蛋白基因Col1a1的显著增加(图1D-E)。二次谐波生成 (SHG) 成像显示,在BT后损伤部位1周内,胶原高度无序(反映为纤维各向异性水平较低),3周时纤维逐渐对齐排列,尽管相对于未损伤对照组仍明显无序(图 1F)。这些发现突出了 MLin 细胞沿着骨软骨谱系的进展,伴随着肌腱损伤后胶原蛋白沉积和组织的增加。
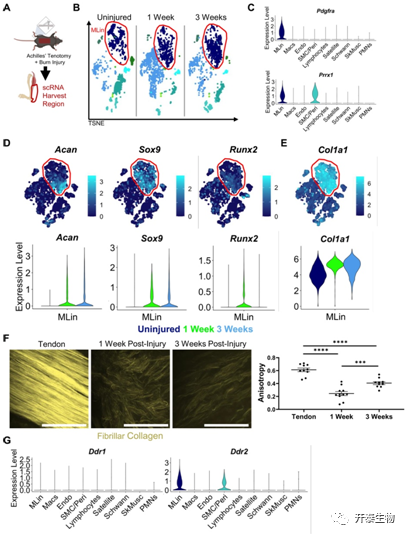
图1. MLin 细胞在受伤后表达纤维状胶原蛋白和胶原蛋白结合蛋白 DDR2
(A)小鼠模型建立和单细胞收集。(B) scRNA 测序 TSNE 图。(C) Pdgfra和Prrx1的基因相对表达的小提琴图。(D) 软骨形成基因(Acan和Sox9)、成骨基因Runx2和 (E) Col1a1的相对表达的 TSNE 和小提琴图。(F)胶原蛋白ECM各向异性定量图。(G) DDR1和DDR2基因相对表达的小提琴图。
之后,研究者团队通过 scRNA 测序,发现盘状结构域受体 DDR1蛋白(Ddr1)在受伤之前或之后表现出最低限度的表达,而DDR2蛋白(Ddr2)在 MLin 和 SMC/Peri 群体中均高度表达,这表明了它在 HO 中发挥正向作用的可能性(图1G)。DDR2 信号转导评分显示,MLin细胞在所有细胞群中得分最高,这表明DDR2信号在该细胞群中优先激活(图2A)。接下来使用转座酶可及染色质 (snATAC) 的单核测定来检测Ddr2和附近基因的转录起始和编码区内的开放染色质区域 ( Ccdc190 ,Hsd17b1和Uap1 ) 受伤后1周在 MLin 细胞中。在Ddr2转录起始位点(TSS) 之前和之后的 MLin 细胞中检测到更开放的染色质结构以及更容易发生转录过程(图2B)。免疫荧光成像显示,HO小鼠和人类组织样本都显示了Ddr2 表达上调和磷酸化/配体激活的 DDR2 (pDDR2)显著增加(图2C-F),表明了DDR2在小鼠和人体组织的肌肉骨骼创伤后具有关键的调控作用。
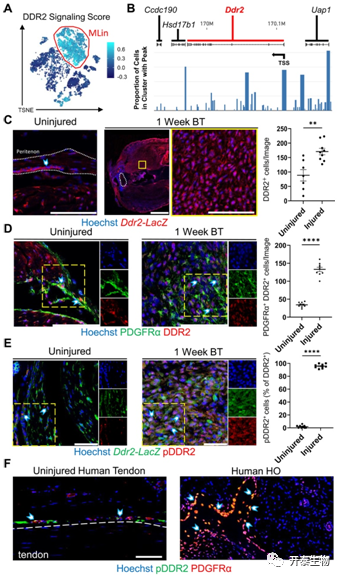
图2. 损伤后 MLin 细胞中的Ddr2表达上调和 DDR2 激活增加
(A)TSNE可视化显示DDR2 信号评分。(B) Ddr2和周围基因轨迹的 snATAC 测序可视化。(C) BT 后 1 周Ddr2-LacZ报告小鼠未受伤肌腱和损伤部位的代表性共聚焦图像 (63×)。(D) 野生型 C57B6 小鼠 BT 后 1 周未受伤肌腱和损伤部位的代表性共聚焦图像 (63×)。Ddr2-LacZ小鼠BT 后 1 周未受伤肌腱和损伤部位的代表性共聚焦图像 (63×)。(F) 用 Hoechst、抗 PDGFRα 和抗磷酸化 DDR2 (pDDR2) 染色的未受伤人肌腱和成熟人 HO 组织的代表性共聚焦图像 (20×)。
02
DDR2 调节损伤后 HO 的形成
接下来研究者团队采用了具有 Ddr2 整体缺失的小鼠 (Ddr2 slie/slie)且进行了BT损伤处理,并与同窝对照 (LC) 进行比较。他们首先使用免疫荧光成像观察到在Ddr2 slie/slie小鼠受伤后 表达 PDGFRα 的 MLin 细胞的平均百分比下降了51.88%(图3A),且软骨形成减少(图3B)。又利用显微计算机断层扫描 (μCT) 成像发现Ddr2slie/slie小鼠受伤的后肢在9周后,成骨作用显著下调且HO形成显著减少(图3C-3E)。在受伤后立即每天用伊马替尼(DDR2抑制剂)治疗16天的 C57B6 小鼠中,损伤后近端和远端残余肌腱以及跟骨 HO 形成显著减少(图3F)。这些结果表明,DDR2 缺失或抑制会阻止成骨,并且可减少体内HO 形成。
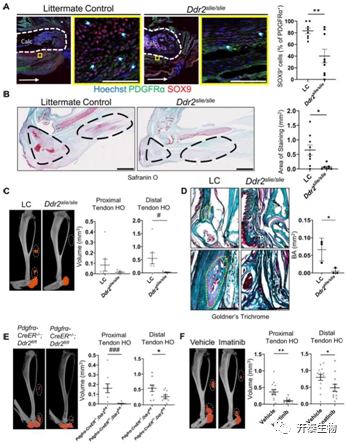
图3. DDR2 缺失或抑制会阻止成骨
(A) BT 后 1 周 LC 和Ddr2slie/slie小鼠的代表性共聚焦图像 (63×)。(B) BT 后 3 周LC 和Ddr2 slie/slie小鼠受伤后肢的番红 O 染色和阳性染色定量。(C) LC和Ddr2slie/slie小鼠(每组5 ~ 7只小鼠)损伤后肢μCT成像,定量BT后9周残余跟腱近端和远端HO骨体积。(D) BT后9周LC 和Ddr2 slie/slie部分肌腱骨区域的 Goldner 毛状体染色。(E) BT后9周 Pdgfrα-CreER−/−、Ddr2fl/fl 和Pdgfrα-CreER+/−、Ddr2fl/fl 小鼠损伤后肢的 μCT 成像。(F) 在 BT 后 9 周,用载体或伊马替尼治疗的野生型小鼠受伤后肢的 μCT 成像。
03
DDR2影响ECM排列
研究者团队接下来将目标聚焦到了ECM上,固定 1 或 3 周后小鼠的胶原原纤维与从移动小鼠中收获的胶原原纤维相比,各向异性更小且更杂乱无章(图4A),但是一周的固定也足以几乎完全减轻 HO生成(图4B),与固定小鼠相比,移动小鼠中的 MLin 细胞表达显著更高量的Col1a1和Col3a1,而Ddr2表达没有检测到差异(图4C-D)。Ddr2 slie/slie小鼠在受伤后 9 周显示 ECM 整齐排列减少(图4E),注意到Ddr2 slie/slie小鼠细胞中粘着斑(FAs) 方向偏差的显著增加(图 4F),表明在没有 DDR2 信号传导的情况下 FAs 更加混乱。这些结果表明DDR2 缺失导致受伤后的 ECM 胶原成分紊乱且排列无序。
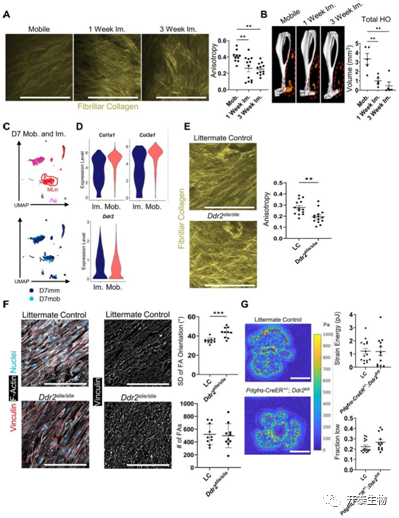
图4. DDR2 缺失导致受伤后 ECM 紊乱
(A) 小鼠损伤部位纤维胶原的 SHG 成像。(B) BT 后 9 周野生型小鼠移动或固定受伤后肢的 μCT 成像。(C) UMAP 图和(D)小提琴图显示从损伤部位采集的细胞scRNA测序中Col1a1、Col3a1和Ddr2的相对表达。(E)BT 后 1 周在 LC 和Ddr2 slie/slie小鼠中损伤部位纤维胶原的 SHG 成像。(F) MLin 细胞的代表性共聚焦图像 (40×)。(G)牵引力显微热图。
04
DDR2调节机械信号转导
研究者团队最后将目标聚焦到了Yes1 相关转录调节因子以及含有转录调节因1 (YAP/TAZ)通路,他们从损伤部位鉴定出了11个独特的簇,其中包括了Prrx1、Pdgfr α 和Ddr2,且显示出增加的Ddr2预测基因活性(图5A-D)。Ddr2的高表达伴随着更高水平的Pdgfrα 和Ddr2基因活性 (图5E-G)。他们发现这Yap1 (YAP) 和Wwtr1 (TAZ)这两个基因在Ddr2高 MLin 细胞中表达更高的活性,明确了 YAP/TAZ 信号潜在重要性(图5H)。最后他们又评估YAP/TAZ 轴的下游目标—Ctgf,并证实了其在Ddr2高表达 MLin簇中具有的更高活性(图5I)。总之,这些数据证明在高表达 Ddr2 的 MLin 簇中,YAP/TAZ 信号通路在损伤后上调。
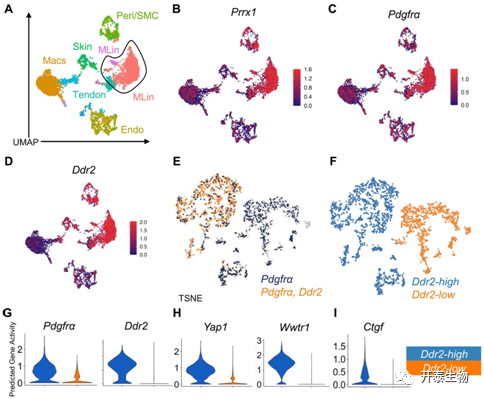
图5. Ddr2调控FAK/YAP/TAZ 轴的snATAC 分析
(A) UMAP 图显示从 BT 后第 0 天、第 7 天和第 42 天的损伤部位采集的细胞的 snATAC 分析中识别出的细胞簇。(B-D) UMAP 图显示了Prrx1、Pdgfr α 和Ddr2的预测基因活性。(E) TSNE 图可视化。(F) TSNE 图显示标记的Ddr2 簇。(G-I) 小提琴图显示预测基因活性Ddr2 -high 和 -low 簇中的Pdgfr α、Ddr2、Yap1、Wwtr1和Ctgf。
+ + + + + + + + + + +
结 论
研究者团队通过一系列转录组学和蛋白质组学分析,确定了DDR2是 HO 形成中的关键 MLin 细胞调节因子。可通过组成型或条件型敲除Ddr2以及药物靶向抑制DDR2信号通路,来减少小鼠体内 HO 的形成。从机制上讲,DDR2 会改变FAK方向和随后的基质组织,调节FAK 和YAP/TAZ介导的 MLin 细胞信号传导,Ddr2缺失会降低FAK/YAP/TAZ 依赖性通路中的 MLin 细胞的成骨分化、HO 形成和胶原排列。因此,ECM-DDR2 相互作用对于驱动 HO 至关重要,并且可以作为治疗该疾病过程的先前未知的治疗靶点。
+ + + + +



