

 English
English文献解读|Nature(48.5):哺乳动物衰老和死亡的普遍转录组学特征
✦ +
+
论文ID
原名:Universal transcriptomic hallmarks of mammalian ageing and mortality
译名:哺乳动物衰老和死亡的普遍转录组学特征
期刊:Nature
影响因子:48.5
发表时间:2026.05.27
DOI号:10.1038/s41586-026-10542-3
背 景
衰老的特征是系统性损伤累积、功能衰退,以及通常呈指数级增长的死亡率。这些表现可以通过遗传、饮食和药物干预进行调节,包括Hutchinson-Gilford早衰症和Klotho基因敲除(Klotho-KO)、热量限制、雷帕霉素和侏儒模型。自2004年以来,干预测试计划(ITP)已在遗传异质性UM-HET3小鼠中评估了50多种化合物,并确定了10多种延长寿命的治疗方法,包括雷帕霉素、阿卡波糖、17-α-雌二醇、卡格列净。尽管已经描述了衰老的哺乳动物分子特征和寿命调节模型,但一直缺乏对衰老、缩短寿命模型和长寿干预措施中共同存在的与死亡相关的机制的统一分析。衰老和干预措施会影响健康和死亡率,但这种调节的潜在分子机制仍不清楚。
实验设计
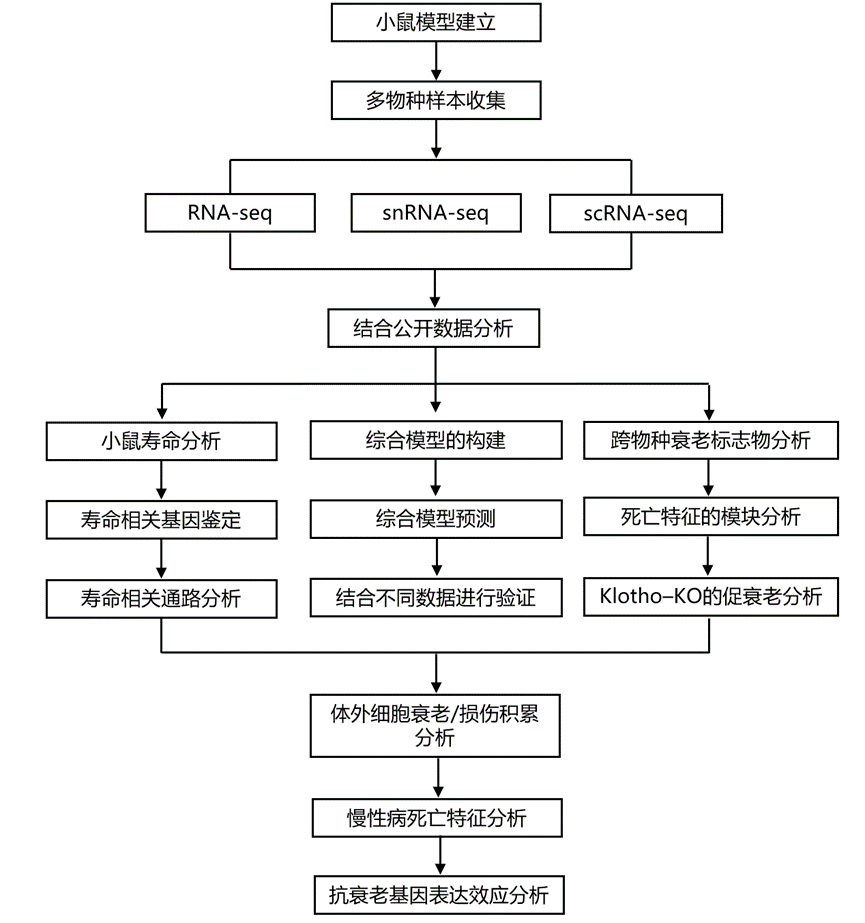
结 果
01

啮齿动物衰老和寿命的生物标志物
为了鉴定寿命和死亡率相关的基因表达特征,研究团队对170只老年(22月龄)遗传异质性UM-HET3雄性和雌性小鼠的肝脏进行了转录组分析(RNA-seq)。这些小鼠接受了20种药物诱导的ITP干预或对照饮食,同时还纳入了12只年轻(4至6月龄)的对照组小鼠(图1a)。干预组包括对生存无影响、有害和有益的化合物,其中包括能显著延长雄性和/或雌性小鼠寿命的干预措施(雷帕霉素、卡格列净、卡托普利、17-α-雌二醇以及雷帕霉素联合阿卡波糖)。这种设计使得能够在统一的死亡率框架下,对衰老和寿命调控相关的分子变化进行联合分析。
为了估算每只动物的预期全因死亡率,他们对每个队列、性别、地点和干预措施的 ITP 生存数据拟合了 Gompertz 模型(图1b),并将按性别和干预措施汇总的队列水平拟合结果整合为 Gompertz 元模型。对于每只测序小鼠,他们计算了预期对数风险率(图1c),即给定年龄和实验组别的队列特异性拟合结果及其对应的元模型预测结果的平均值(方法)。模型模拟的寿命与生存曲线的经验估计值非常吻合(图1d),这支持使用 Gompertz 拟合来捕捉干预措施对寿命的影响。他们还分析了标准化年龄,即实际年龄除以预期最大寿命,从而得出两个综合指标——预期死亡率和标准化年龄——将实际年龄与干预引起的寿命调节结合起来。
为了将研究结果推广到ITP之外,他们构建了一个啮齿动物基因表达元数据集,该数据集包含来自26种组织的3575个小鼠和大鼠样本,涵盖了广泛的年龄范围(4-943天)和多个品系。此外,他们还纳入了964个来自接受各种药物、遗传、饮食和环境干预的动物样本,这些干预对寿命的影响各不相同,有的延长寿命,有的缩短寿命,有的则没有影响(图1e)。最终,他们获得了来自96个数据源和79种干预措施的4539个啮齿动物转录组数据。
利用已发表的生存数据,他们再次按品系、性别和干预措施拟合了队列特异性和汇总的 Gompertz 模型,以计算每个样本的预期对数风险率、标准化年龄和最大寿命(图1f)。模型模拟的第 90 百分位寿命与经验估计值一致。预期死亡率随年龄增长而增加,并受遗传背景、早衰模型、高脂饮食和延长寿命治疗的影响。
对于跨研究的多组织特征,他们通过将每个样本的表达量中心化到同一数据集、组织和性别中随机选择的匹配对照组的中位数表达量来计算相对表达量。线性混合效应模型揭示了9059-9167个与实际年龄、标准化年龄和预期死亡率相关的基因,以及3098-4439个与年龄校正后的最大寿命和死亡相关性状相关的基因(图1g-h),表明寿命和死亡率在分子水平上受到系统性调控。
与寿命呈负相关的显著基因包括Igf1(一种在物种内和物种间均存在的经典长寿调节因子)和Ddost(参与晚期糖基化终产物加工,并与癌症生存率降低相关)。而促长寿基因包括Fmo3(一种mTOR和炎症抑制剂)和Nmrk1(参与NAD+生物合成)。Igf1的表达水平随年龄增长而下降,而Fmo3的表达水平则升高,表明衰老相关的变化既包含有害成分,也包含补偿成分。
与 ITP 的结果一致,基因和通路水平上,生理年龄特征与年龄校正后的死亡率特征重叠,而寿命特征则相反。干扰素、白细胞介素和 p53 信号通路、补体和凝血以及其他炎症和应激反应程序与寿命呈负相关,与衰老和死亡率呈正相关,而氧化磷酸化和线粒体翻译则呈现相反的模式(图1i)。脂肪酸和外源性物质代谢也与死亡率呈负相关,支持其在寿命调控和累积损伤清除中的作用。总之,这些分析在一个大型多组织元数据集上定义了啮齿动物衰老、死亡率和寿命的保守转录组特征。
为了进一步提高性能,减少批次效应,他们构建了“相对”时钟,方法是将每个转录组样本以同一数据集和组织中随机选择的匹配对照参考组为中心,并预测相应的年龄差异(图1j)。这提高了啮齿动物多组织时钟的准确性,并且对参考组的选择具有鲁棒性,其性能与表观遗传时钟相当。为了估计转录组年龄(tAge)的不确定性(这对于小样本量尤其有用),他们还训练了贝叶斯岭回归(BR)版本的时钟,其准确性也与之相似。
组织特异性生物钟(肾脏、肝脏、大脑和骨骼肌)以及小鼠特异性多组织模型与啮齿动物生物钟的性能相当(图1k),表明整合组织并不会显著降低性能。其他机器学习模型[支持向量机 (SVM)、轻量级梯度提升机 (LightGBM)、随机森林和k近邻 (KNN)]的性能均未优于弹性网络模型。
为了评估模型在不同组织和数据集上的泛化能力,他们进行了留一组织(LOTO)和留一数据集(LODO)检验。在LOTO分析中,相对多组织时间时钟能够捕捉所有保留组织中的年龄动态(图1l),表明系统性衰老相关特征在不同器官间是共享的。在LODO分析中,相对时钟在独立数据集上的表现优于绝对时钟,这支持了数据集内中心化能够有效减轻批次效应。因此,他们在整个研究中都重点关注相对时钟。
在实际年龄、标准化年龄和死亡率模型中,时钟系数均呈正相关。实际年龄和死亡率时钟共有的主要正相关特征包括Gpnmb、Cst7和Cdkn1a(图1n),它们分别编码GPNMB(一种炎症相关糖蛋白)、CST7(一种溶酶体组织蛋白酶抑制剂,与神经退行性疾病有关)和CDKN1A/p21(一种p53通路细胞周期抑制剂)。相反,死亡率系数最高的负相关基因包括Nrep、Col1a1和Col3a1,这些基因与分化和伤口愈合相关,可能反映了衰老和短寿动物再生能力的丧失。与时钟系数一致,Cst7和Cdkn1a的表达随着年龄增长和年龄调整后的死亡率增加而增加,而Nrep、Col1a1和Col3a1的表达随着年龄增长和年龄调整后的死亡率增加而减少。
接下来,他们通过将时钟模型的预测结果与独立数据集中预期最大寿命的年龄和干预相关变化进行关联,来检验时钟模型是否能够同时捕捉到生理年龄和寿命调节效应。作为基准,他们训练了一个“寿命时钟”,该时钟能够直接根据表达变化预测最大寿命的差异。将生理年龄作为特征添加后,其性能并未显著提升,这表明年龄信息已充分编码在表达谱中。生理时钟的预测结果与年龄高度相关,但与干预引起的寿命变化相关性较低,而寿命时钟则呈现相反的模式(图1o)。相比之下,归一化年龄时钟和死亡率时钟能够同时追踪年龄和寿命效应,并且在不同组织中表现一致。
当干预措施分为缩短寿命模型和延长寿命模型时,基于时间时钟的模型能够检测到有害干预措施(与年龄匹配的对照组相比,tAge 值更高),但未能有效捕捉到延长寿命的干预措施,这与既往报道一致,即延长寿命的干预措施通常不会显著逆转与年龄相关的转录组特征。相比之下,标准化年龄时钟,尤其是死亡率时钟,能够区分短寿命模型和长寿命模型,预测有害干预措施的 tAge 值更高,而延长寿命的干预措施的 tAge 值更低(图1p)。值得注意的是,死亡率时钟比专门的寿命时钟更能区分寿命调节模型,这表明整合衰老和干预措施的信息可以提高分子健康生物标志物的整体性能。
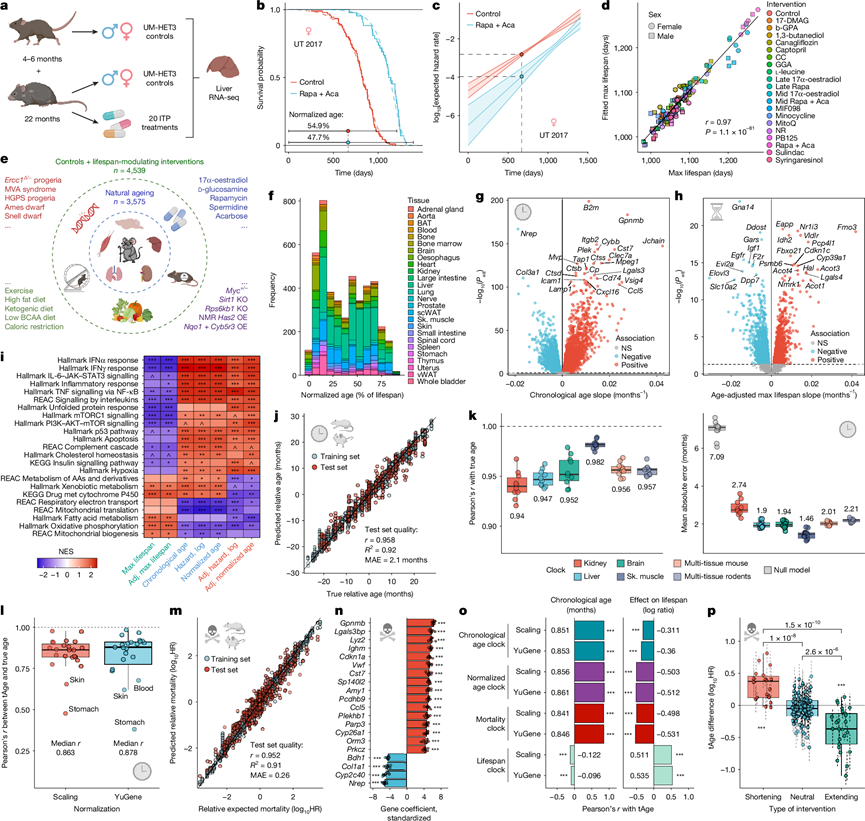
图1. 啮齿动物多组织转录组时钟捕捉与衰老和死亡相关的分子变化。
(a) ITP 测序样本概览。(b) 对照组 ITP 小鼠以及接受雷帕霉素 (Rapa) 和阿卡波糖 (Aca) 治疗的小鼠的生存情况。(c)预期全因死亡率的 Gompertz 拟合曲线。(d) ITP 队列中,基于原始生存数据(x轴)或 Gompertz 拟合曲线(y轴)的最大寿命。(e) 小鼠和大鼠组织基因表达样本汇总用于荟萃分析的示意图。(f) 啮齿动物样本的标准化年龄和组织。标准化年龄是指给定年龄、品系、性别和干预措施下预期最大寿命的比例。(g) 对照动物的年龄基因表达特征。(h) 根据年龄校正的预期最大寿命的转录组特征。(i) 对衰老特征、最大寿命、标准化年龄和预期死亡率进行 GSEA 分析。(j) 弹性网络啮齿动物多组织转录组时钟在训练集和测试集上相对时间年龄的性能。(k) 器官特异性和多组织相对时间年龄时钟在十个随机选择的测试集上的准确性。(l) 弹性网络多组织时间年龄时钟的留一组织交叉验证性能。(m) 基于YuGene的弹性网络啮齿动物多组织转录组时钟在训练集和测试集上预测相对预期死亡率的性能。(n) 基于YuGene的弹性网络死亡率时钟中绝对系数最大的前20个基因。(o) 弹性网络时钟在独立数据集上预测实际年龄(左)和干预寿命效应(右)的准确性。(p) 平均死亡率。在缩短寿命、中性和延长寿命的干预措施中,分别针对每个数据集和性别,使用弹性网啮齿动物多组织死亡时钟评估处理组和对照组样本之间的年龄差异。
02
保守的跨物种衰老生物标志物
为了检验衰老和死亡的多组织特征是否在啮齿类动物和灵长类动物中保守,他们扩展了元数据集,纳入了来自30多种组织的2623个食蟹猴样本和来自脑、骨骼肌、皮肤和血液的4003个人类样本。对于每个样本,他们计算了其实际年龄(根据物种最大寿命进行调整)、标准化年龄以及使用已发表的生存数据计算的预期死亡率。
利用该元数据集,他们训练了统一的多物种、多组织(“通用”)相对生物钟,用于预测时间年龄、标准化年龄和预期死亡率。这些模型在所有四个物种中均表现良好(图2a),在leave-one-fold-out (LOFO)分析中,物种内Pearson系数r≥0.94 (时间年龄)和r≥0.91 (死亡率)。在所有测试样本中,时间年龄与实际年龄的相关系数r =0.952,与泛哺乳动物DNA甲基化生物钟相当。在leave-one-species-out (LOSO)分析中,时间钟捕捉到了未见物种的衰老轨迹,表明衰老的核心转录组特征在哺乳动物中具有普遍性。
跨物种比较揭示了保守的基因和通路水平衰老特征。啮齿类动物和灵长类动物中一致上调的基因包括Gpnmb、Vsig4、Cdkn1a和Eda2r,而保守的下调标记包括Nrep、Col1a1和Col3a1(图2b)。物种特异性衰老斜率呈正相关,无论是在整体上还是在单个组织内(图2c)。跨物种一致的功能变化包括干扰素、TNF 和 p53 信号通路、缺氧和补体/凝血的上调,以及 DNA 修复、线粒体翻译、氧化磷酸化和胶原蛋白形成的下调。总之,这些结果支持哺乳动物衰老的转录组结构在不同组织和物种中具有保守性。
为了检验器官层面的衰老和死亡特征是否在单个细胞类型中得到重现,他们分析了单细胞转录组分析(scRNA-seq)数据集,该数据集涵盖了不同年龄段C57Bl/6J小鼠的多种组织。为了提高样本覆盖率,他们采用了元细胞方法,将来自同一组织和同一动物的随机选择的细胞计数合并(图2d)。首先通过汇总每个组织和每只动物的所有可用细胞,并应用小鼠多组织时间时钟来验证该策略。该时钟能够高精度地捕捉到不同器官的年龄动态。改变元细胞大小揭示了覆盖率和时钟性能之间的饱和关系:在每个元细胞约 100 个细胞时,组织间的中位数r = 0.90,而在每个元细胞 25 个细胞时,中位数r = 0.84(图2e-f)。
随后,他们分别对每种细胞类型应用了元细胞分析和多组织时钟。值得注意的是,49种细胞类型中有48种(98%)和46种(94%)分别显示小鼠实际年龄与基于时间时钟和死亡时钟预测的tAge之间存在显著的正相关性(图2g-h)。这一结果得到了独立验证,他们使用荧光激活细胞分选(FACS)技术对来自年轻(2月龄)和年老(22-24月龄)C57BL/6雌性小鼠的细胞类型进行了转录组分析,结果显示,两种时钟均检测到所有细胞类型(每组至少3个样本)的tAge存在显著差异。值得注意的是,干细胞群(间充质干细胞和造血干细胞)也表现出显著的促死亡表达转变,表明衰老相关的变化不仅限于分化谱系。
因此,基因水平对衰老过程中tAge增加的贡献在不同细胞类型中呈正相关。主要的共同促死亡变化包括衰老、炎症和凋亡相关基因的上调,例如Cdkn1a、Lgals3、Casp1、S100a8和S100a4,以及Sparc的下调,Sparc编码细胞外基质(ECM)合成、分化和伤口愈合的调节因子(图2i)。总之,这些数据表明,尽管组织和细胞类型具有不同的特性和功能,但衰老伴随着跨组织和细胞类型的保守的损伤累积转录组学特征。

图2. 衰老和死亡的转录组生物标志物在哺乳动物物种和细胞类型中是保守的。
(a) 基于YuGene的弹性网络多物种多组织相对预期死亡率时钟的LOFO性能。(b) 使用混合效应模型检验了在不同组织和物种中与年龄始终相关的40个基因。(c) 跨物种基因表达年龄斜率的成对Spearman相关性。(d) 基于元细胞的scRNA-seq分析tAge。(e) BR小鼠多组织时间时钟对年龄预测的准确性与Tabula Muris Senis组织中元细胞大小的关系。(f) BR时间时钟对年龄预测的准确性与每个元细胞的平均基因覆盖率的关系。(g) 表示使用 BR 小鼠时间时钟(左)和死亡率时钟(右)估算的各细胞类型中实际年龄与 tAge 之间的 Pearson 相关性。(h) 使用 BR 时间时钟(左)和死亡率时钟(右)估算的代表性细胞类型的 tAge 动态变化。(i) 衰老过程中导致不同细胞类型中与死亡相关的转录组变化的 前 40 个基因。
03
模块时钟追踪死亡率特征
为了识别衰老和长寿过程中可作为通路特异性定量生物标志物的共调控转录组成分,他们对以各数据集、组织和性别为中心的啮齿动物相对表达元数据集应用了加权基因共表达网络分析(WGCNA)。结果揭示了28个稳健的模块(图3a),每个模块包含30-630个基因,这些基因在衰老过程中以及不同干预措施下均表现出协同表达变化。分别在雄性和雌性中独立获得的模块显著重叠,表明其在不同性别间具有稳健性。过滤后,各模块显示出显著不同的通路富集,重叠极少,其中包括富集于炎症/先天免疫反应、线粒体翻译/电子传递和细胞外基质组织/上皮间质转化(EMT)的模块。
为了评估各模块的生物学一致性,他们在校正实际年龄后,检验了各模块第一主成分之间的偏相关性。功能相关的模块聚集在一起——例如,免疫模块(炎症、干扰素和 T 细胞信号传导)与代谢相关模块明显区分开来。与基因和通路水平的结果一致(图1i),免疫模块与年龄和死亡率呈正相关,与预期最大寿命呈负相关,而线粒体翻译、氧化磷酸化和脂质代谢模块则呈现相反的模式(图3b)。大多数模块与衰老和寿命呈相反的相关性;然而,热应激反应、翻译和 ECM/EMT 模块与这两个指标均呈同向相关性,提示存在适应性或补偿性反应。一些模块(例如内质网中的蛋白质加工和 mTOR 信号传导)与寿命有关,但与年龄无关,这表明寿命调节部分独立于生理衰老。
为了基于这些转录组网络成分构建一个精简的生物标志物组合,他们针对每个模块训练了多组织生物钟,用于预测生物体的实际年龄和预期死亡率(图3c)。基于所有模块基因(2141个基因)训练的全局模型与全转录组生物钟的性能相当,表明这些模块共同捕获了大部分与死亡率相关的变异。各个模块的生物钟对其目标变量的预测准确度中等。剔除性能较差的模块后,他们保留了23个功能注释的模块特异性生物钟,用于预测生物体的实际年龄和死亡率(图3c)。在测试样本中,所有死亡率时钟均与实际年龄呈正相关,与预期最大寿命呈负相关(图3c),这与死亡率指标的综合性质一致。模块间的关系在 tAge 水平上得以保留——例如,与免疫相关的时钟预测结果聚集在一起。
接下来,他们利用已建立的急性炎症应激体内模型验证了模块特异性时钟,并将其应用于注射脂多糖 (LPS) 或磷酸盐缓冲液 (PBS) 的 5 至 6 月龄 C57BL/6J 小鼠的脑基因表达数据。根据综合时间时钟和死亡率时钟(图3d)以及大多数模块时钟(图3e),LPS 均能增加雌雄小鼠的 tAge,其中免疫模块(炎症、干扰素信号通路和适应性免疫反应)的促死亡变化最为显著,这与 LPS 诱导的神经炎症一致。差异表达基因的富集分析证实了炎症通路的显著激活(图3e)。
相反,与同龄对照组相比,热量限制小鼠的肝脏样本在许多死亡模块时钟中均表现出tAge降低(图3e),其中代谢相关模块[血红素代谢/活性氧(ROS)通路、脂质代谢/PPAR信号通路和胆固醇代谢/mTOR信号通路]的影响最为显著。与此一致的是,热量限制诱导了参与细胞呼吸、氧化磷酸化以及脂质、血红素、脂肪酸和氨基酸代谢的基因的广泛上调,这与其代谢重塑和延长寿命的作用相一致。综上所述,这些结果表明分子年龄可以通过不同的生物通路进行调控,并且模块特异性时钟提供了一个可解释的框架,用于量化有害或促进健康的干预措施在通路水平上的影响。
为了进一步验证本研究的生物标志物,并在已建立的早衰症模型中阐明与死亡相关的机制,他们对6只Klotho基因敲除小鼠和6只年龄匹配的C57BL/6J对照小鼠的肾脏和骨骼肌进行了RNA-seq。Klotho是一种促长寿因子,可调节IGF1-mTOR、NRF2和TGFβ信号通路;其缺陷会导致早衰症,使小鼠的最大寿命限制在4-5个月,而过表达则可延长寿命。
正如预期,Klotho在对照组肾脏中高表达,而在Klotho敲除小鼠的肾脏和骨骼肌中几乎检测不到。为了避免Klotho表达对tAge预测的直接影响,他们重新训练了啮齿动物多组织生物钟,排除了Klotho,并将其应用于对照组和Klotho敲除小鼠的样本。时间生物钟和死亡率生物钟均检测到Klotho敲除小鼠肾脏和骨骼肌中tAge升高(图3f),且在肾脏中的影响更为显著,因为该基因通常在肾脏中表达。在导致tAge升高的主要基因中,Cdkn1a在早衰小鼠的肾脏和骨骼肌中均上调(图3g),这与Klotho在抑制细胞衰老中的作用相一致。总体而言,基因水平对 tAge 升高的贡献在肾脏和肌肉之间呈正相关。
多个模块特异性死亡时钟也显示Klotho基因敲除小鼠的tAge加速,尤其是在能量代谢和NRF2信号通路模块中(图3h)。基于呼吸/线粒体翻译、氧化磷酸化/三羧酸循环和胆固醇代谢/mTOR信号通路相关基因训练的时钟显示,两种组织中的tAge均显著增加,而NRF2信号通路/蛋白酶体模块时钟则分别显示肾脏和肌肉中tAge显著和接近显著的加速。
在Klotho敲除小鼠肾脏中,呼吸/线粒体翻译和胆固醇代谢/mTOR信号通路模块的基因是导致复合促死亡转变的最强贡献者之一。与此同时,参与呼吸、线粒体翻译、氧化磷酸化和脂肪酸代谢的基因普遍下调,这与老年和短寿动物的模式相似(图1i),并支持代谢功能受损是Klotho缺陷的关键后果。相比之下,炎症和干扰素模块在Klotho敲除小鼠的两种组织中均下调,且不会导致tAge加速。尽管免疫相关模块的生物钟总体上最为准确(图3c),但它们仍然基本没有反应,这表明该早衰模型主要通过代谢紊乱而非炎症激活来加速分子衰老。还观察到组织特异性效应:ECM 组织/EMT 和肌肉收缩/细胞骨架模块分别选择性地加速肾脏和肌肉中的 tAge(图3h)。
为了检验Klotho-KO是否会在不同细胞类型中诱导系统性促衰老效应,他们对8周龄对照小鼠和同龄Klotho -KO小鼠的肾脏和脑组织进行了单细胞核转录组分析(snRNA-seq)。均匀流形近似和投影(UMAP)解析了肾脏和脑组织的主要细胞类型(图3i-j),并且Klotho -KO小鼠肾脏中所有细胞类型的Klotho表达均显著降低,这与整体数据一致。在分别对每种细胞类型和基因型的元细胞进行聚合,并应用排除Klotho基因的啮齿动物多组织时钟进行训练后,死亡率时钟检测到除亨利氏袢外的所有肾脏细胞类型中tAge均显著加速(图3i),而时间时钟显示出类似但并不完全显著的效应。近端肾小管细胞中与死亡相关的tAge基因贡献与肾脏整体水平呈正相关,突显了snRNA-seq和RNA-seq结果的一致性。主要的共同促死亡肾脏生物标志物包括上调的Cdkn1a和Cp(编码铜蓝蛋白糖蛋白,该糖蛋白参与铁代谢,并在多种疾病模型中表达升高),以及下调的Mogat1、Cyp51和Fmo5。
在脑组织中,对照组的基线Klotho表达水平较低,其敲除相关的下调仅在神经元和少突胶质细胞中显著,除少突胶质细胞前体细胞外,其他大多数细胞类型也呈现类似的趋势。死亡率和生物钟均检测到神经元、星形胶质细胞和少突胶质细胞的tAge显著加速,内皮细胞的tAge加速接近显著水平(图3j)。相比之下,小胶质细胞的死亡率tAge略有下降,这与Klotho-KO小鼠脑组织 中未观察到炎症激活相一致(图3h)。这些结果共同表明,寿命调节干预措施可以对不同细胞类型和功能成分产生截然不同的健康相关影响,凸显了衰老和死亡特征的异质性。

图3. 共调控基因表达模块定义了衰老和长寿的基本分子特征。
(a) 基因共表达网络(谱嵌入),其中选定的彩色模块由WGCNA识别。(b) 每个模块的第一主成分与实际年龄(左)、预期死亡率(中)或年龄校正后的预期最大寿命(ML;右)之间的Spearman相关性。(c) 基于单个模块、所有模块组合或整个转录组基因训练的弹性网络啮齿动物多组织时钟的性能。(d) 注射 PBS 或 LPS 的 5 至 6 月龄雄性(左)和雌性(右)C57BL/6J 小鼠脑组织的死亡率 tAge。(e) 由热量限制 (CR) 在肝脏和 LPS 在脑组织中引起的标准化死亡率 tAge 差异,使用模块特异性死亡率时钟进行估计。(f) Klotho-KO和对照C57BL/6J小鼠(每组n = 6只小鼠)肾脏(上)和肌肉(下)的死亡年龄。(g) 在Klotho-KO肾脏(左)和肌肉(右)中,基于不包括Klotho的弹性网啮齿动物死亡率时钟,导致Klotho- ko肾脏(左)和肌肉(右)中促或抗死亡转录组变化的前25个基因。(h) 不同模块特异性死亡时钟下,Klotho-KO小鼠与对照组小鼠的标准化死亡率差异;不同模块特异性死亡时钟下,Klotho-KO小鼠与对照组小鼠的标准化死亡率差异;显著性通过方差分析(ANOVA)进行检验。(i-j) UMAP(左图)以及对照组和Klotho -KO小鼠肾脏和脑中不同细胞类型的元细胞死亡率tAge差异(右图)。
04
细胞损伤诱导死亡生物标志物
为了检验体内衰老和死亡特征是否能在细胞损伤累积模型中重现,他们将多组织转录组时钟应用于体外衰老和应激诱导数据集。在培养的人类原代成纤维细胞中,时间时钟和死亡时钟均显示,不同供体细胞的tAge随时间呈显著线性增加(图4a),这与细胞倍增速度的减缓相一致。在对照WI-38细胞中,复制性衰老也伴随着类似的tAge升高(图4b)。在两种模型中,tAge与时间的相关性均高于与倍增次数或传代次数的相关性,表明tAge信号先于明显的表型衰退。在培养过程中,原代成纤维细胞和WI-38细胞中与死亡相关的基因表达变化具有一致性,其中CDKN1A是主要贡献基因之一(图4c)。值得注意的是,WI-38细胞中hTERT的过表达抑制了细胞衰老,并消除了tAge的增加(图4b),表明转录组时钟主要反映的是损伤相关的变化,而非时间本身。与此一致的是,由人原代成纤维细胞诱导产生的多能干细胞(iPSC)的tAge值相对于供体细胞均有所降低(图4d),这与之前的分子研究结果一致。
接下来,他们测试了急性损伤干预是否会在体外加速tAge的发生。在用代谢应激剂寡霉素或2-脱氧葡萄糖(2-DG)处理的人类原代成纤维细胞中,两种生物钟均检测到tAge较供体匹配的对照组有所增加(图4e)。暴露于20 Gy γ射线照射的小鼠和裸鼹鼠胚胎及皮肤成纤维细胞在两种物种和细胞类型中也均显示出tAge升高(图4f和扩展数据图7h),这支持了转录组生物钟反映生物系统中累积损伤负荷的观点。相比之下,γ射线照射或hTERT过表达均未改变人类细胞中的DNA甲基化年龄,这表明DNA甲基化和转录组生物钟追踪衰老的互补方面,表观遗传标记反映长期的随机变化,而基因表达则捕捉更动态的功能状态。
γ射线照射后,tAge加速现象在多种啮齿动物死亡模块时钟中均有体现,包括染色质修饰、干扰素信号传导、呼吸/线粒体翻译、细胞骨架/糖酵解和胆固醇代谢/mTOR信号传导(图4g),并伴有干扰素、白细胞介素和TNF信号传导、炎症和细胞凋亡相关基因的上调。
为了表征导致人类细胞模型衰老相关变化的通路,他们定义了四种哺乳动物共有的多物种共表达模块。啮齿类和灵长类动物的模块存在显著重叠,对合并的元数据集进行WGCNA分析,揭示了15个富集于特定通路的多物种模块,这些通路包括炎症、干扰素信号通路、细胞外基质/上皮间质转化(ECM/EMT)和mRNA剪接,每个模块的内部相关性均高于随机基因集。随后,他们构建了14个互不重叠的模块特异性通用时间时钟和死亡率时钟,其平均LOFO性能r > 0.3(图4h)。与啮齿类动物的对应模块类似,通用死亡率模块时钟能够追踪实际年龄和预期最大寿命。
将通用时钟应用于重编程数据,结果显示iPSC中大多数时间时钟和死亡时钟的tAge值均降低(图4i),其中ECM/EMT模块的再生信号最强,这与重编程过程中的去分化和间质-上皮转化一致。EMT相关基因在iPSC中也相应下调(图4i)。相反,代谢应激因子寡霉素和2-DG主要在代谢模块中增加死亡tAge值,包括脂肪酸代谢、氧化磷酸化/血红素代谢、细胞骨架/糖酵解和线粒体翻译(图4j),这进一步支持了模块时钟能够精确定位受损伤调节干预影响最大的通路。
比较不同模型中基因水平对死亡率和年龄变化的影响发现,复制培养和γ射线照射产生相似的模式,而重编程和hTERT过表达形成相反的聚类,寡霉素和2-DG则介于两者之间。一些基因在细胞损伤和再生模型中表现出一致的行为(图4k),包括Cdkn1a、应激反应基因Apod以及Eda2r(一种炎症调节因子,也是转录组和蛋白质组水平上可靠的衰老生物标志物)。因此,这些基因代表了衰老和应激诱导的损伤在不同生物组织层次积累的候选通用生物标志物。
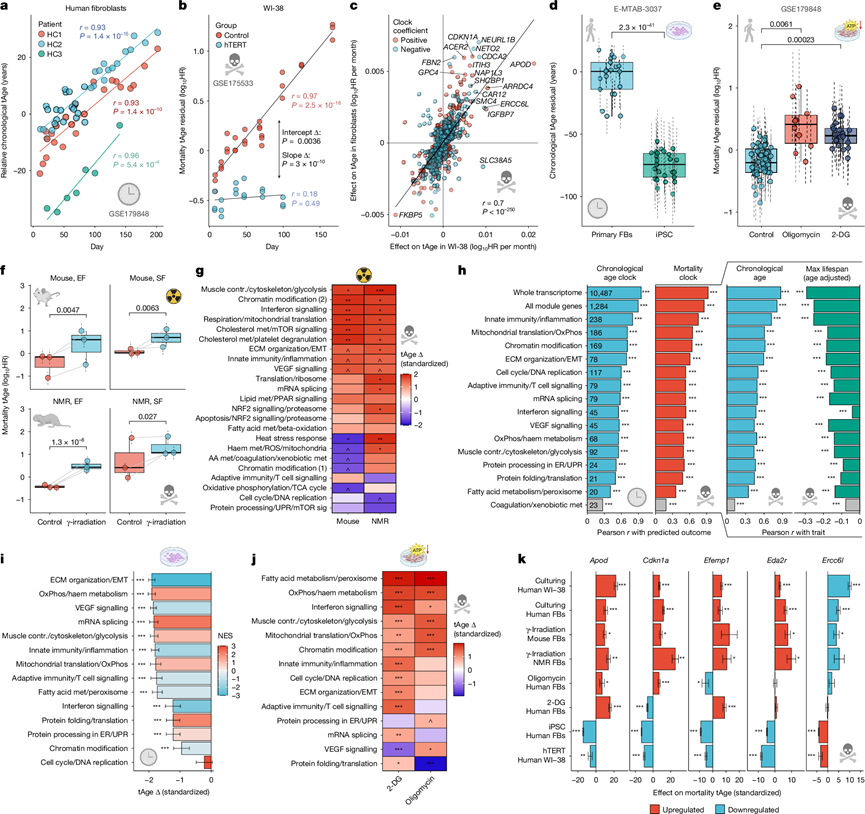
图4. 基于组织的转录组时钟可检测体外细胞衰老和损伤积累。
(a) 利用弹性网络通用时间时钟估计人原代成纤维细胞培养过程中的时间年龄(tAge)动态变化。(b) 利用弹性网络通用死亡时钟估计对照组(n = 30)和hTERT转导组(n = 18)WI-38细胞培养过程中的死亡率年龄(tAge)动态变化。(c) 基于弹性网络通用死亡时钟,比较培养的原代成纤维细胞和对照组WI-38细胞中基因水平死亡率相关表达变化的一致性。(d) 使用BR通用时间时钟估计的原代成纤维细胞(FB)(n = 24)和匹配的诱导多能干细胞(n = 25)的时间tAge残差。(e) 使用BR通用死亡率时钟估计的对照组(n = 46)、寡霉素处理组(n = 10)和2-DG处理组(n = 20)人原代成纤维细胞(培养≥40天)的死亡率tAge残差。(f) 小鼠和裸鼹鼠皮肤成纤维细胞 (SF) 和胚胎成纤维细胞 (EF) 在对照组和 γ 射线照射条件下的死亡率 tAge。(g) 使用模块特异性啮齿动物死亡率时钟估计的照射组和对照组小鼠(左)和裸鼹鼠(右)成纤维细胞之间的标准化死亡率 tAge 差异。(h) 基于单个模块、所有模块组合或整个转录组基因训练的弹性网络多物种多组织时钟的准确性。左图:时间时钟(蓝色)和死亡率时钟(红色)的LOFO性能(Pearson相关系数r)。右图:死亡率tAge与实际年龄(蓝色)或年龄校正后的预期最大寿命(绿色)之间的Pearson相关系数r。(i) 由人类成纤维细胞重编程诱导的标准化时间tAge变化(n = 49),使用模块特异性通用时间时钟进行评估。(j) 寡霉素或2-DG处理后,使用模块特异性通用死亡时钟评估的标准化死亡率tAge变化。(k) 根据弹性网络通用死亡时钟,对细胞模型中死亡率tAge变化贡献最大的5个基因。
05
慢性病死亡率特征
为了探究慢性病理是否会诱导与衰老和死亡相关的转录组变化,他们分析了九个啮齿动物数据集,涵盖了阿尔茨海默病、慢性肾脏病(CKD)、糖尿病肾病、缺血性卒中、非酒精性脂肪性肝炎和肝细胞癌(HCC)模型,每个模型均设有年龄匹配的对照组。啮齿动物多组织时间时钟和死亡时钟显示,除HCC外,所有疾病模型和器官的tAge均显著加速(图5a)。在缺血性卒中模型中,心脏的死亡tAge在第1天开始增加,并在第3天进一步增加;脑组织的死亡tAge在第3天时在年轻和老年动物中均表现出更显著的增加。小胶质细胞的tAge在第14天时在所有时钟中均表现出一致的增加,且同侧半球的tAge高于对侧半球。这些结果突显了疾病相关分子衰老动力学的时空和器官特异性异质性。
相比之下,HCC肝脏的死亡率tAge低于年龄匹配的对照组(图5a)。与此信号相关的基因富集于细胞周期、去分化和ECM组织(Plekhb1、Col1a1、Col3a1、Col4a1、Ube2c和Cdk1)。相应地,EMT/ECM和细胞周期模块对tAge的降低贡献最大(图5b),而EMT/ECM模块时钟的tAge降低幅度较小。同时,炎症和氨基酸/异生物质代谢模块促进了tAge的增加(图5b),并伴有炎症、干扰素和p53信号通路的激活,以及呼吸、氧化磷酸化和脂肪酸代谢的抑制。许多模块特异性的细胞死亡时钟在HCC中也显示出tAge的增加。总之,这些数据表明,肿瘤结合了类似年轻细胞的特征(如增殖、去分化和细胞外基质重塑)以及促衰老的标志(包括炎症和代谢紊乱)。
基因水平对tAge增加的贡献在不同疾病模型中基本一致(图5c)。Gpnmb、Cdkn1a和Lgals3在不同疾病中表现出最一致的促死亡作用,在至少五个模型中显著上调(图5d)。这些基因与免疫调节和慢性病理有关,它们也促进了LPS诱导的小鼠脑内tAge加速,这与LPS相关特征和疾病相关特征之间的正相关性一致(图5c)。
啮齿动物模块特异性死亡时钟显示,在大多数疾病模型中,大多数细胞子系统的 tAge 均有所增加(图5e),其中干扰素和炎症模块的影响最大且最一致,这与它们在各种疾病中的广泛上调以及慢性炎症在年龄相关病理学中的核心作用相一致。只有松果体中的阿尔茨海默病和心脏中的缺血性中风缺乏炎症模块的加速,并且单独聚类(图5c)。
与啮齿动物数据一致,通用死亡时钟在校正年龄和性别后,检测到克罗恩病、溃疡性结肠炎、慢性肾病、心力衰竭和阿尔茨海默病患者组织中tAge显著加速。为了进一步检验死亡时钟是否能前瞻性地预测死亡时间,他们分析了弗雷明汉心脏研究(FHS)的血液数据。校正年龄和性别后,全因死亡率与通用复合死亡时钟、多模块死亡时钟以及几个模块特异性死亡时钟(染色质修饰、炎症/先天免疫和氧化磷酸化/血红素代谢)预测的tAge显著相关(图5f)。几种多组织转录组时钟的性能与第二代人类血液DNA甲基化时钟(DunedinPACE、YingDamAg和PhenoAge)相当,并且优于现有的人类转录组时间时钟(Peters和RNAAgeCalc),无论是在完整队列还是在共享的患者子集中均是如此(图5f)。值得注意的是,实际年龄的准确性与死亡时间的预测在很大程度上是分离的——例如,Horvath2时钟在年龄预测方面最为准确,但与死亡率无显著相关性,而DunedinPACE和染色质修饰转录组时钟则显示出较弱的年龄相关性,但与死亡率的相关性很强。
虽然采用不同的训练模式,但在调整年龄和性别后,转录组和DNA甲基化时钟预测结果在患者间呈正相关(图5g)。在转录组模型中,染色质修饰时钟与表观遗传时钟的相关性最高,表明这些模式之间存在相互作用,这与DNA甲基化和染色质状态之间的机制关系一致。
最后,他们检测了衰老、死亡和疾病的常见转录组学生物标志物,包括GPNMB、CDKN1A、LGALS3、S100A6和ITGB2,是否也与人类血液中的死亡时间和心血管结局相关。经年龄和性别调整的 Cox 比例风险模型显示,这些标志物与死亡率和新发心血管疾病 (CVD)、卒中和充血性心力衰竭 (CHF) 总体呈正相关,其中LGALS3和S100A6对大多数结局具有显著性意义 。在英国生物银行超过5万名参与者的血浆蛋白质组学数据中,GPNMB、CDKN1A和LGALS3蛋白水平与全因死亡率、多种疾病(包括心脏骤停、心力衰竭、肝硬化、糖尿病、肾衰竭、抑郁症和动脉粥样硬化)以及危险因素(包括肥胖、高血压和睡眠障碍)呈正相关(图5h)。这些发现共同表明,Gpnmb、Cdkn1a和Lgals3是跨组织、跨物种和跨分子层面的保守的死亡率和慢性疾病生物标志物。

图5. 哺乳动物衰老相关疾病表现出与死亡相关的共同转录组特征。
(a) 使用BR啮齿动物多组织死亡时钟估计对照组(灰色)和年龄匹配的慢性疾病组(彩色)动物的死亡年龄(tAge)。(b) 使用复合弹性网络啮齿动物死亡时钟估计2岁B6C3F1小鼠(每组n =6只小鼠)的对照组和HCC组肝脏中各模块对死亡年龄(tAge)差异的贡献。(c) 基于弹性网络啮齿动物死亡时钟,计算不同疾病模型中基因水平与死亡率相关的表达变化的成对Pearson相关性。(d) 根据弹性网络啮齿动物死亡时钟,计算慢性疾病中导致死亡率tAge增加的前5个基因。(e) 慢性疾病对标准化死亡率tAge的汇总效应,基于模块特异性啮齿动物死亡时钟。(f) 转录组(绿色)和DNA甲基化(DNAm;蓝色)时钟与FHS人类血液数据中死亡时间的关联。(g) DNA甲基化(DNAm;列)与弹性网络通用死亡率转录组时钟(行)之间的偏Spearman相关性。(h) 血浆GPNMB、CDKN1A和LGALS3与英国生物银行(UKB)中人类死亡时间、疾病结局和风险因素的关联。
06
异体共生对健康的促进作用
为了鉴定体内分子年龄减缓的转录组特征,他们分析了接受异时共生(HPB)的小鼠的肝脏基因表达。在HPB中,老年小鼠与年轻伴侣共享血液循环,并表现出多种衰老相关表型的逆转,包括干细胞功能增强、髓鞘再生、神经发生和血管生成增加,以及心脏肥大减少。此前,他们发现HPB可降低20月龄小鼠与3月龄伴侣小鼠血液和肝脏中的表观遗传年龄(eAge),且这种效应在分离后仍然存在,并伴有寿命延长;而年轻伴侣小鼠则表现出短暂的eAge升高,并在恢复后恢复正常。
他们分析了接受异时或同时共生3个月后,立即处死(“附着”)或分离2个月后处死(“分离”)的年轻和年老小鼠的肝脏转录组。在年老的异时共生小鼠中,啮齿动物多组织死亡时钟检测到附着时间点tAge显著降低,而时间时钟则显示出不显著的趋势(图6a)。分离2个月后,两种时钟均检测到tAge显著降低。年轻的异时共生小鼠在HPB后立即表现出比同时对照组更高的tAge,但这种差异在恢复后减小,与eAge的变化趋势相似。
在老年小鼠(合并附着组和脱离组)中,HPB 相关 tAge 降低的最强贡献者是Nrep,它在暴露于年轻血液后上调(图6b),这与增强的再生潜能一致。抗死亡变化还包括Cdkn1a和Vcam1的下调,抑制它们可以减轻神经炎症并改善老年小鼠的学习和记忆。
在老年小鼠中,HPB诱导的tAge减缓反映在大多数模块特异性生物钟中(图6c),表明其具有跨多个子系统的系统性健康促进作用。与此一致的是,老年异时共生小鼠表现出炎症、干扰素、p53和凋亡通路下调,以及氧化磷酸化和脂质代谢上调,这与衰老和死亡相关的通路特征相反(图1i)。在幼鼠中,HPB的转录组效应较弱且复杂,兼具促死亡和抗死亡的特征。
为了研究哺乳动物发育过程中tAge的动态变化,他们分析了小鼠从受精卵到出生的胚胎发生基因表达谱。先前的研究报道,在胚胎发育第4.5天(E4.5)至E10.5期间存在一个DNA甲基化恢复期,随后eAge单调增加,这与“零点”模型相符。利用小鼠多组织转录组时钟(包括生理年龄和死亡率),他们观察到了类似的U型tAge轨迹(图6d),tAge最低值出现在E10附近,95%置信区间为E8.5至E15,这支持了基因表达水平上的系统性恢复期。
E10前后基因对tAge的贡献呈负相关,表明在胚胎复苏期和E10后阶段,主要由一些共同的基因发挥着相反的作用。关键贡献基因包括Cdkn1a以及炎症调节基因S100a8、S100a9和Lgals3,这些基因在E10之前表达下调,之后表达上调。早期胚胎发生过程中,基因组范围内的mRNA表达也出现下调,随后在E10之后出现广泛的上调。然而,仅使用各阶段上调或下调的基因重新计算tAge,结果仍然显示E10之前tAge显著下降,之后上升,表明U型tAge动态变化并非完全由全局转录组重塑驱动。
在胚胎发育第10天(E10)之前表达下调,随后表达上调的基因富集于免疫应答和脂质代谢相关通路;而表达模式相反的基因则富集于细胞周期和mRNA剪接相关通路。与此一致的是,炎症应答、干扰素信号通路、p53通路和脂肪酸代谢在胚胎发育第10天之前普遍受到抑制,随后激活。基于模块贡献和模块特异性生物钟,炎症和干扰素模块也是胚胎复苏信号的主要贡献者之一(图6e)。大多数模块生物钟(例如呼吸/线粒体翻译、脂质代谢和热应激应答)的tAge动态变化呈U型,但部分模块生物钟(例如细胞周期)在早期发育过程中tAge升高,表明存在混合通路水平的信号。
与死亡相关的基因表达变化导致的tAge减缓在包括老年HPB、热量限制和早期胚胎发生在内的多种衰老减缓模型中呈正相关(图6f),其中HPB和热量限制模型之间的相似性尤为显著。共同的返老还童相关特征包括Cdkn1a以及炎症因子Ccl5、S100a9和Lgals3的下调(图6g)。总体而言,Cdkn1a和Lgals3是哺乳动物衰老、返老还童和死亡过程中最稳定的分子标志物之一,适用于各种疾病和寿命调节干预措施。
为了检验胚胎复原是否在不同细胞谱系中保守,他们分析了E6.5至E8.5胚胎的scRNA-seq数据,并使用最优运输方法重建了每种E8.5细胞类型的祖先轨迹。推断的谱系与已知的胚胎起源相符。在整个胚胎水平(将每个样本的所有细胞合并为元细胞)上,基于时间序列和死亡率的小鼠多组织时钟均检测到从E6.5到E8.5的tAge显著降低(图6h),这与整体数据一致。谱系加权元细胞分析显示,在几乎所有检测的细胞类型中,tAge 值在这一时间段内均显著下降(图6i-j),涵盖所有三个胚层,包括中胚层(例如内皮细胞、近轴中胚层和咽部中胚层)、内胚层(例如肠道)和外胚层(例如神经嵴和脊髓)。相比之下,胚外内胚层的 tAge 值略有上升,这与胚外组织在类似阶段 eAge 值升高的情况一致,表明早期再生可能仅限于胚内谱系。某些谱系(例如心肌细胞)的 tAge 值呈 U 形曲线,从大约 E8 开始增加(图6j-k),表明 tAge 的起始点可能因组织和检测方式而异。

图6. HPB 和早期胚胎发生表现出抗衰老基因表达效应。
(a) 采用BR啮齿动物多组织死亡时钟模型估算3月龄(左)和20月龄(右)C57BL/6J小鼠肝脏在3个月同时共生或HPB治疗后的死亡年龄(tAge)。(b) 基于弹性网络啮齿动物死亡时钟模型(logFC×时钟系数),分析老年异时共生体( n =12)中导致促死亡和抗死亡转录组变化的前25个基因。(c) 使用模块特异性啮齿动物死亡时钟估计老年肝脏中由HPB诱导的标准化死亡率tAge差异。(d) 使用BR小鼠多组织时间时钟估计C57BL/6J小鼠胚胎发生过程中的时间tAge。(e) 发育日与基于模块特异性啮齿动物时钟估计的 tAge(左)或死亡率 tAge(右)之间的 Pearson 相关性。(f) 基于弹性网络啮齿动物死亡率时钟,计算不同年龄减速模型中基因水平死亡率相关表达变化的成对 Pearson 相关性。(g) 根据弹性网络啮齿动物死亡率时钟。(h) 使用BR小鼠多组织时序钟估算的C57BL/6小鼠胚胎间叶细胞(E6.5-E8.5)的年龄。(i) 基于线性混合效应模型,计算E6.5-E8.5期间不同细胞谱系的时序tAge变化斜率。(j) 计算E6.5-E8.5期间特定细胞谱系的时序tAge轨迹。(k) 根据基因表达数据估算和比较tAge的TACO应用界面截图。
+ + + + + + + + + + +
结 论
本研究整合了来自四种哺乳动物(小鼠、大鼠、猕猴和人类)25种以上组织的11000多个转录组,旨在开发准确、可解释的啮齿动物和多物种生物标志物,用于预测实际年龄和预期死亡率,并预测寿命调节干预措施、死亡时间、慢性疾病和返老还童的效果。与衰老相关的变化在不同物种和细胞类型中均保守存在,揭示了哺乳动物衰老和死亡的普遍转录组特征,包括CDKN1A和LGALS3,它们的蛋白水平也与英国生物银行(UK Biobank)中的死亡率和多种疾病相关。在体内和体外损伤累积模型中,均观察到与死亡相关的特征,包括炎症、复制性衰老、代谢抑制和γ射线照射,而细胞永生化、重编程、异时共生和早期胚胎发生可减弱或逆转这些特征。网络分析揭示了衰老和死亡相关标志的模块化结构,涵盖炎症、干扰素信号传导、线粒体功能、染色质修饰和细胞外基质组织。为了分析单个细胞组分的衰老,本研究构建了模块特异性时钟,揭示了干预措施的通路特异性效应:慢性疾病主要加速炎症模块的衰老,而热量限制和Klotho缺乏则靶向线粒体和代谢模块。转录组和DNA甲基化时钟显示,人类血液中衰老加速的相关性日益增强,其中染色质相关模块时钟的相关性最为显著,凸显了分子衰老模式之间的机制联系。本研究揭示了细胞死亡调控的保守特征和模块化结构,为量化和靶向调控不同物种和组织中细胞子系统的衰老提供了框架。
+ + + + +



