

 English
English文献解读|Nat Commun(15.7):小鼠海马突触分辨率的整合转录组学和蛋白质组学图谱
✦ +
+
论文ID
原名:An integrated transcriptomic and proteomic map of the mouse hippocampus at synaptic resolution
译名:小鼠海马突触分辨率的整合转录组学和蛋白质组学图谱
期刊:Nature Communications
影响因子:15.7
发表时间:2025.08.26
DOI号:10.1038/s41467-025-63119-5.
背 景
对大脑区域、神经元群和突触类型的转录组和蛋白质组分析揭示了大脑在细胞和亚细胞分子多样性方面的巨大潜力。然而,目前缺乏同时全面研究神经元转录本和蛋白质多样性的研究。mRNA和蛋白质的定量分析之所以必要,任何一种分子的丰度都不能可靠地由另一种分子的丰度预测。动态的mRNA-蛋白质关系受到mRNA稳定性、定位、运输、翻译和降解等因素的影响,最终导致二者水平的差异。例如,在果蝇胚胎发育过程中,特定区域富集的mRNA会影响局部蛋白质组,但这并不反映mRNA的总体丰度。在非洲爪蟾视网膜锥体发育中也观察到了类似的上下文依赖性关系,在神经元可塑性过程中树突mRNA翻译的时空调控中也观察到了类似的现象,其中局部mRNA的翻译会响应环境信号,蛋白质丰度也会相应变化。这些因素强调了开展综合分析以理解细胞和亚细胞特征分子蓝图的必要性。然而,此类研究仍然十分匮乏。
实验设计
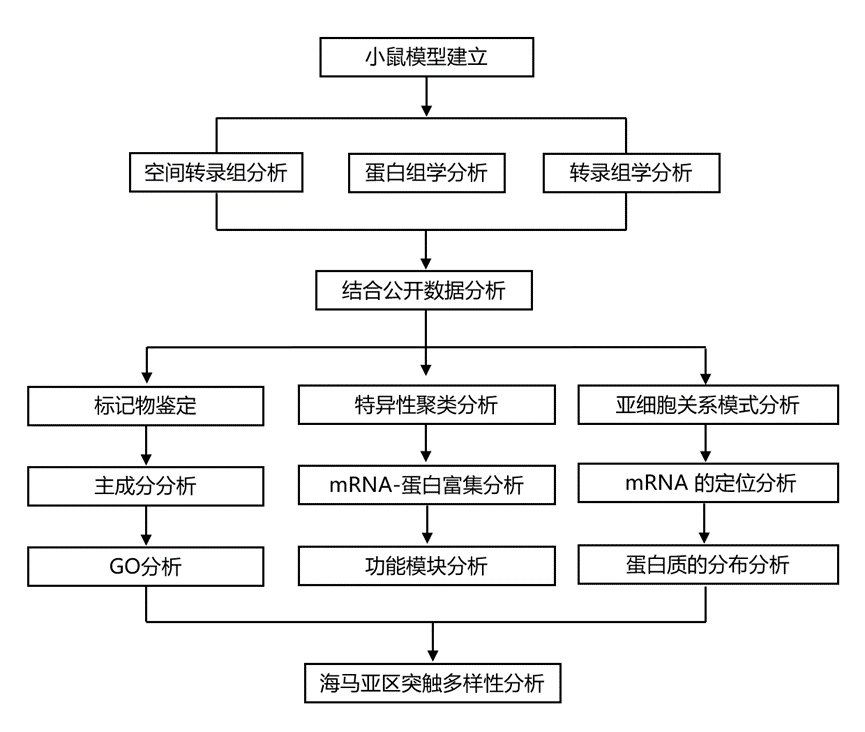
结 果
01

用于小鼠海马体深度空间转录组和蛋白质组分析的流程
研究团队开发了一种用于对急性分离的小鼠海马组织进行深度空间转录组和蛋白质组分析的流程,从而克服了需要组织固定的传统方法所固有的技术难题。他们在解剖显微镜下进行手工解剖,并将之前用于大鼠海马CA1、CA3和DG15的解剖方案应用于小鼠海马(由于CA2和CA3在解剖显微镜下视觉上无法区分,因此将它们一起解剖)。利用离散层的视觉标记,他们进一步解剖了CA1的四个层[基底树突层(SO)、由细胞体组成的锥体层(SP)、以及分别包含近端和远端顶树突的辐射层(SR)和腔隙分子层(SLM)]。对于每个样本,他们将两只小鼠的组织混合,以获得足够的材料用于后续的转录组和蛋白质组分析,从而减少技术误差和实验偏差(图 1a)。通过检测每个区域已知分子标记的富集情况来验证解剖的准确性;这包括CA1区的Fibcd1和Homer2,CA3区的Bok和Nectin3,以及DG区的Prox1和Calb2。CA2区的标记物也在CA3样本中得到了进一步验证。CA1层中富集的标记物对应于其已知的中间神经元、胶质细胞和突触细胞群的组合:SO区的Trhde和Chrm2,SP区的Cck和Pvalb,SR区的Map2和Lrrtm1,以及SLM区的Ndnf和Adgrl2。这些数据表明,该工作流程能够识别从精确显微解剖的海马体中分子的空间分布。
利用开发的组织显微切割和制备流程(图 1a),他们对CA1、CA3和齿状回(DG)亚区的转录组和蛋白质组进行了分析。该流程实现了深度覆盖,定量分析了各亚区超过10000个蛋白质,并检测到超过17000个mRNA转录本(图1c-d)。虽然所有转录本和绝大多数蛋白质(约93%)在所有亚区均能检测到,但他们仍使用无监督主成分分析(PCA)探究了各亚区是否存在不同的特征。结果表明,各亚区高度聚类,表明它们在分子水平上存在差异(图 1d)。除了上述已知的亚区标记富集外,他们还检测到各亚区之间数千个mRNA和蛋白质的差异表达(图1e-g),所有这些数据均可在可搜索平台https://syndive.org.上找到。他们进一步通过原位杂交和免疫荧光染色验证了每个亚区域的关键候选者。
为探究各海马亚区富集的分子群体,他们通过GO分析验证这些转录本与蛋白质是否承担不同的功能角色。在CA1区,mRNA与蛋白质组学数据均显示出突触后功能与突触整合相关条目的富集,具体表现为Shank3、Homer2等支架蛋白,以及GABA受体亚基Gabra3、谷氨酸受体亚基Grin3a、5-羟色胺受体亚基Htr1b和乙酰胆碱受体亚基Chrna5等神经递质受体的显著富集(图1f-h)。CA3区则凸显出轴突运输与髓鞘相关过程,并富集调控神经元兴奋性的钾通道家族成员。在DG区,核内过程与基因调控成为最显著的富集通路,其分子基础包括转录因子及Wnt信号通路相关分子。蛋白质组数据中,转录调控因子Mecp2的富集尤为引人注目——该蛋白的突变与Rett综合征及MECP2重复综合征相关,已知会破坏齿状回的神经发生过程。
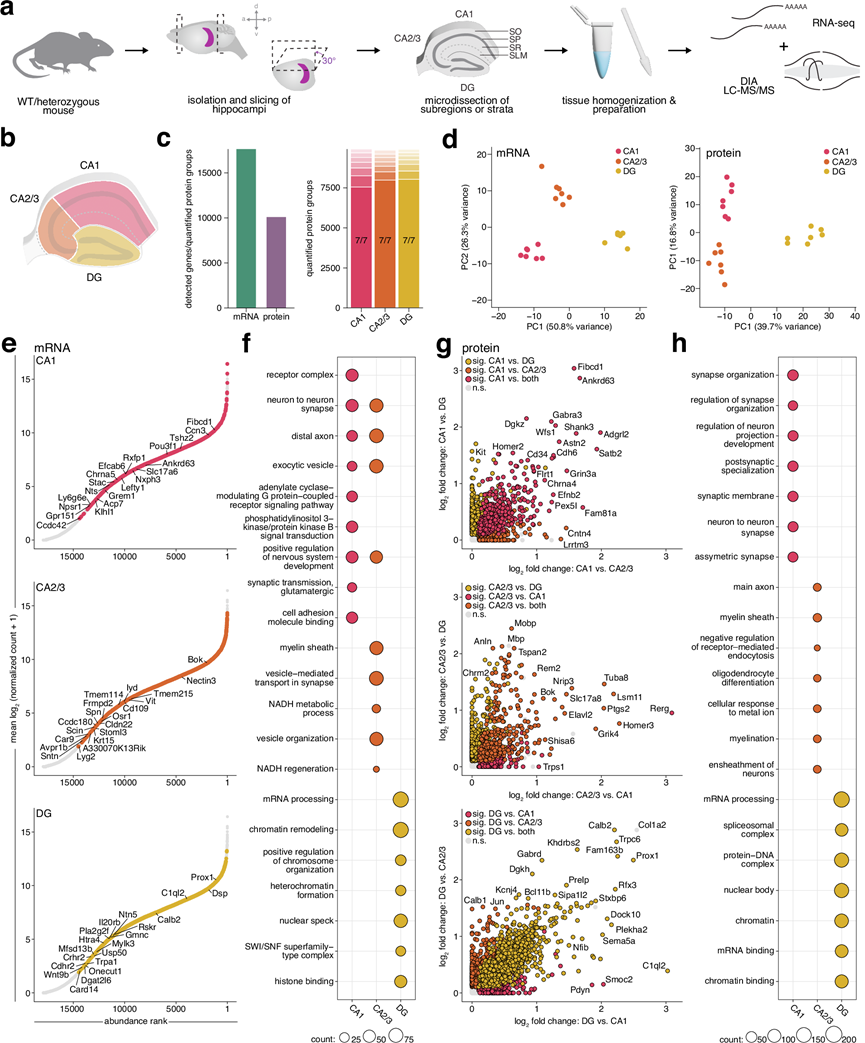
图1. 小鼠海马深度空间转录组学和蛋白质组学分析流程,以及亚区域转录组和蛋白质组的比较分析。
(a) 组织分析的实验流程。(b) 示意图显示为准备组织而进行显微切割的 CA1、CA2/3 和 DG 亚区域。(c) 左图:所有亚区域中检测到的 mRNA 转录本总数和定量蛋白质组。右图:每个亚区域定量蛋白质组的有效值条形图。(d) 转录组(左)和蛋白质组(右)的主成分分析 (PCA) 显示了按亚区域聚类的情况。(e) mRNA的等级丰度图表示给定亚区域中转录本的相对丰度。(f) 基于与其他两个亚区域相比每个亚区域中显著富集的转录本。(g) 散点图显示每个亚区域中蛋白质相对于其他两个亚区域的富集。(h) 基于每个亚区域中与其他两个亚区域相比显著富集的蛋白质的过度表达分析。
02
层特异性转录组和蛋白质组揭示了区室化的分子特征和功能作用
接下来,他们探究了CA1锥体神经元各层区室的这些富集模式可能存在的差异。鉴于其层特异性结构,他们研究了mRNA和蛋白质如何促进SP、SO、SR和SLM及其相关中间神经元群的功能特化(图 2a)。通过分析超过17000个mRNA转录本和约10000个蛋白质(图 2b),他们鉴定了与CA1各层特定功能相对应的独特表达模式(图 2),并验证了关键候选基因。这种分层的分子组织结构,如PCA聚类所示(图 2c),揭示了数千个mRNA和蛋白质的显著富集(图 2d-f)。GO分析进一步表明,这些分子的空间调控既能形成局部微环境,又能使神经元区室分化(图 2e-g)。
在SO中,检测到的Trhde、Erbb4和Chrm2的显著富集标志着重要的中间神经元群体。其他富集的mRNA编码参与运输和转运的蛋白,如驱动蛋白和P4-ATP酶,其相关通路涉及微管及轴突运输(图 2e)。他们还发现了高度富集的髓鞘相关蛋白,这些蛋白共同参与髓鞘的形成与维持。这与大量有髓鞘的锥体细胞轴突穿过邻近的齿状回以及位于内层/齿状回交界处的中间神经元相一致,这些中间神经元有助于抑制性回路的形成,从而调节海马网络的活动。
在SP中,观察到的Cck转录本和小清蛋白蛋白质的富集标志着篮状细胞群体的存在,该类细胞与锥体细胞形成胞周连接。虽然该层以胞体为主,但GO分析显示与神经元及突触膜组织相关的mRNA显著富集,特别是受体和离子通道的mRNA,表明这些分子在胞体合成后将运输至作用位点(图2d-e)。其中包括与神经系统疾病相关的分子。由于锥体细胞层容纳了锥体细胞的细胞核,该层富集突触相关mRNA符合生物学逻辑。相反,该层富集的蛋白质主要与细胞核内过程相关(图2f-g)。该层的转录组与蛋白质组功能差异最为显著,这很可能源于其作为细胞核所在层位,负责转录绝大部分神经元mRNA的特性。
SR是CA1区神经毡的主要组成部分,主要由顶端树突构成,这些树突接收来自CA3区轴突的Schaffer侧支输入。对mRNA和蛋白质谱的通路分析突显了与突触功能和修饰相关的过程(图 2e-g)。富集的mRNA包括影响磷酸化的激酶(Stk38、Pdpk1、Prkcb、Braf、Mapk9、Ttbk2、Pdk3),这些激酶与“肽基丝氨酸磷酸化”相关,肽基丝氨酸磷酸化是一种调节突触功能和可塑性的翻译后修饰。例如,PSD-95在Ser295位点的磷酸化增强了其在突触处的稳定性和积累,从而促进突触强度和AMPA受体的募集。在突触前末梢,α-突触核蛋白Ser129位点的磷酸化通过促进其与突触前蛋白(如Vamp2和突触蛋白)的相互作用,在增强其突触前靶向性方面发挥着关键作用。转录组分析还揭示了泛素-蛋白酶体系统(UPS)家族成员(Usp9x、Usp40、Usp34、Usp32、Usp24、Usp48、Usp31、Mindy2)的富集,强调了蛋白质的周转和调控,以及其在突触和突触可塑性中的作用。如阿尔茨海默病中所见,UPS的失调会破坏肌浆网突触功能,而调节UPS活性可以恢复长时程增强作用。蛋白质组分析进一步突显了自噬相关过程,其中富集了诸如Atg5之类的蛋白质,这些蛋白质调节突触小泡的周转和自噬以支持可塑性。总的来说,SR转录组和蛋白质组在对突触功能和可塑性至关重要的蛋白质调控机制上趋于一致。
SLM直接接收来自内嗅皮层、联合核和颞下皮层等脑区的输入,其转录组特征反映了这一点,突出了细胞黏附、细胞外基质重塑、生长因子信号传导和代谢活动(图2e)。Ndnf转录本在此富集,可作为该层中具有密集树突的神经胶质样GABA能中间神经元的标志。SLM还包含不同的星形胶质细胞亚型,驱动相关转录本Slc1a3、Fam107a、Agt和Gfap的富集。虽然SLM中富集程度最高的蛋白质包括受体(Grm2、4、6、8)和黏附分子(Ntng1、Cd44、Fbln5),表明大部分富集蛋白质是线粒体蛋白。已知线粒体在神经元不同区室中存在形态差异,这可能反映了处理和整合输入信号或为远离细胞体的局部蛋白质合成提供能量所需的代谢需求增加。
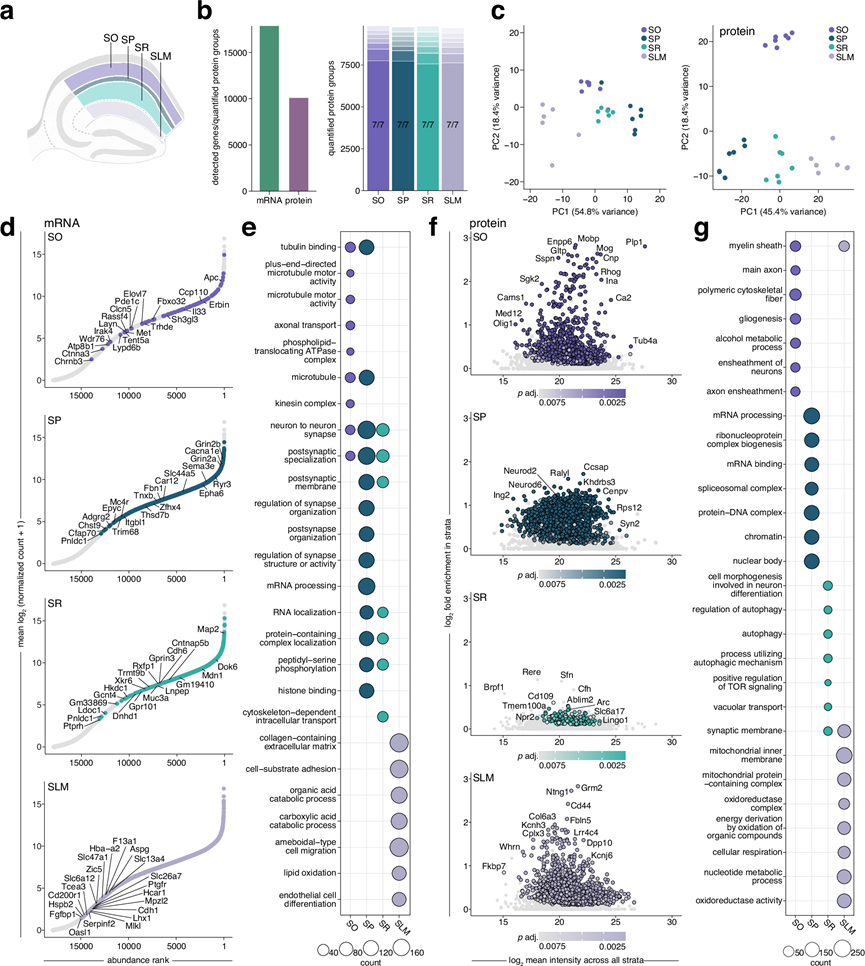
图2. CA1 中转录本和蛋白质的层特异性组织产生了专门的身份。
(a) 示意图显示显微切割的 CA1 层 SO、SP、SR 和 SLM。(b) 左图:所有层中检测到的总 mRNA 转录本和定量蛋白质组数据。右图:每个层定量蛋白质组的有效值条形图。(c) 转录组(左)和蛋白质组(右)的PCA显示了层特定的聚类。(d) mRNA的等级丰度图表示与整个 CA1 相比,给定层中转录本的相对丰度。(e) GO分析。(f) 蛋白表达强度分析。(g)GO分析。
03
转录本和蛋白质的空间靶向分析揭示了与神经元信号传导、代谢和疾病相关的功能模块
CA1区各层次的分子多样性促进了不同特性的形成,其空间区室化塑造了神经元信号传递和新陈代谢等过程。通过分析归属于五个GO注释的所有分子(聚焦于膜蛋白)的分布,对于所有离子通道类别,其mRNA均显示出在SP最显著的富集趋势,这与该层作为转录发生、富含锥体神经元细胞核的特性一致(图5a)。钾通道蛋白在锥体细胞层中呈耗竭状态,而在所有其他层次中均呈现正向富集分布。对于钠或钙可透过的离子通道复合物,其表达分布更具极性。体现这种组织结构的代表性蛋白包括富集于SO而在SLM耗竭的AMPA型谷氨酸受体亚基Gria4,以及在SO耗竭的NMDA型谷氨酸受体亚基Grin2a和Grin2b。这与已知的沿CA1树突的钾、钠、钙通道梯度以及海马体内整体分子梯度一致(图3b)。与受体分布也遵循脑内梯度的现象一致,他们发现GPCR相关蛋白呈现从始层主要耗竭向SLM逐渐富集的转变,GPCR mRNA的分布则较不均匀。线粒体mRNA遵循相似模式,而在SP和SLM中,线粒体蛋白几乎仅在后者的SLM富集。这反映了远端树突因局部线粒体融合/分裂动态而具有的高线粒体含量(图3b)。通过分析涉及线粒体融合与裂变的蛋白质及其mRNA分布,他们发现融合调节因子确实在SLM富集,融合与运输的双重调节因子在SR富集,而代谢与质量控制相关蛋白的混合组则在SO富集。值得注意的是,Mfn1蛋白及其mRNA在SLM共富集,暗示了局部翻译机制的存在。
CA1各层中mRNA和蛋白质的空间分布也对疾病具有重要意义。海马体特定层的脆弱性与多种神经系统疾病相关,因为分子紊乱通常会对某些层产生不成比例的影响。例如,亨廷顿病的关键分子Htt mRNA和蛋白质在SR层富集(图3c-d),而SR层的可塑性缺陷和突触功能障碍已充分证实,包括SR-CA1突触损伤和AMPA受体扩散失调。单细胞转录组分析(scRNA-seq)研究揭示了阿尔茨海默病(AD)患者海马体中显著的转录组变化,尤其与SO层及其邻近的肺泡内的突触功能障碍、炎症反应和代谢改变有关,这些区域还存在不成比例的高水平细胞凋亡。这与之前的研究结果一致,即在阿尔茨海默病(AD)中,SO细胞表现出大量的抑制性中间神经元丢失,从而损害了反馈抑制。这种损害因轴突和髓鞘的损伤(已得到充分证实)以及错误折叠的朊病毒蛋白和β-淀粉样蛋白的积累而加剧,其中SO细胞受到的影响尤为严重。他们在SO细胞中检测到Prnp和App的富集(图 3c-d),这与这些已知的疾病表型相符。Fmr1基因突变是脆性X综合征的特征。在本研究数据集中,Fmr1蛋白和mRNA在SP细胞中含量最高(图3c-d),这与其作为翻译抑制因子在翻译中的作用一致。在本研究的数据集中,其他区室中检测到的Fmr1 mRNA与其作为RNA结合蛋白参与mRNA运输的额外功能相符。如Fmr1敲除细胞所示,该过程的紊乱会损害突触mRNA靶向和局部蛋白质合成,而这些紊乱可能是常见的脊柱异常的潜在原因。总而言之,这些例子凸显了理解CA1区分子层特异性分布的重要性,因为这可能揭示神经系统疾病的潜在脆弱性。
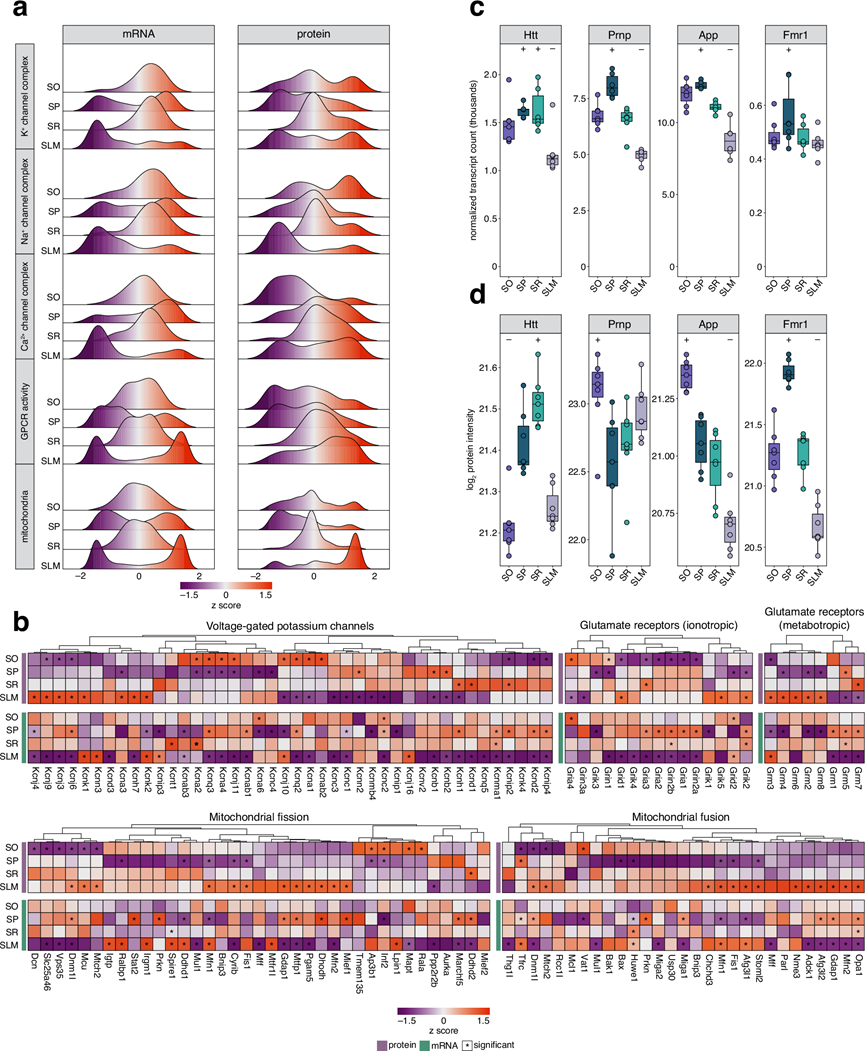
图3. 功能基因和致病基因的空间组织。
(a) 选定基因集的 CA1 层间差异表达基因的岭图。(b) 与 (a)中所示的过程或细胞成分相关的转录本和蛋白质的 z 得分的候选热图。(c-d) 在疾病中起致病作用并显示出层特异性富集的 mRNA 和蛋白质丰度箱线图。
04
mRNA和蛋白质丰度比较揭示了亚细胞关系模式和动态变化
由于他们在CA1区域观察到候选mRNA和蛋白质的分布模式既有重叠也有相反的,他们进一步整合了数据集,以观察这些分子之间关系的更广泛趋势。首先探究单个mRNA及其对应蛋白质的丰度是否相关,以及这种相关性是否随层级变化。与之前的报道一致,他们发现所有层级中mRNA和蛋白质的丰度均存在中等程度的相关性,其中SP层的相关性最高(图 4a)。为了推断层级间蛋白质水平的差异在多大程度上是由相应mRNA的差异造成的,他们分析了各层级间mRNA-蛋白质相对丰度变化的相关性(图 4b)。结果显示,存在数百个高度正相关和负相关的基因对。正相关的例子包括突触支架蛋白Dlg1(在SLM中富集度最高)以及RNA结合蛋白Caprin1和RNA剪接蛋白Rbfox1(二者在SP中相对丰度最高)。负相关的分子包括蛋白酶体亚基Psma1以及核糖体蛋白Rpl26和Rps8(图 4b)。为了探究所有正相关和负相关蛋白对相关的生物学过程和分子功能,他们进行了GO富集分析。正相关的mRNA和蛋白质富集于蛋白质-RNA调控复合物以及与神经元结构和动力学相关的过程(图 4c)。这表明,对于这些蛋白类群而言,它们在不同层级中的相对丰度与mRNA的可用性密切相关。相比之下,负相关的基因对主要与核糖体相关,以及与突触小泡动力学、蛋白质降解和髓鞘形成相关的基因(图 4c)。这些分子可能受到转录后或翻译后调控,其蛋白质水平与mRNA水平脱钩。蛋白质周转也可能影响这种关系,因为半衰期不同的蛋白质表现出不同的mRNA-蛋白质依赖性;例如,树突相关蛋白的半衰期比参与核糖体或蛋白酶体功能的蛋白短。他们通过将正相关和负相关的mRNA和蛋白质与描述海马神经元中蛋白质半衰期的数据集进行比较,验证了这一假设。他们将总共 2956 个基因与其蛋白质半衰期进行了比对。分析表明,正相关的 mRNA-蛋白质对的蛋白质半衰期显著较短(图 4d),这提示周转率较高的蛋白质可能通过其对应 mRNA 的局部翻译得以维持。相反,负相关的蛋白质半衰期显著较长,表明它们的水平更稳定,受局部 mRNA 可用性的影响较小(图 4d)。
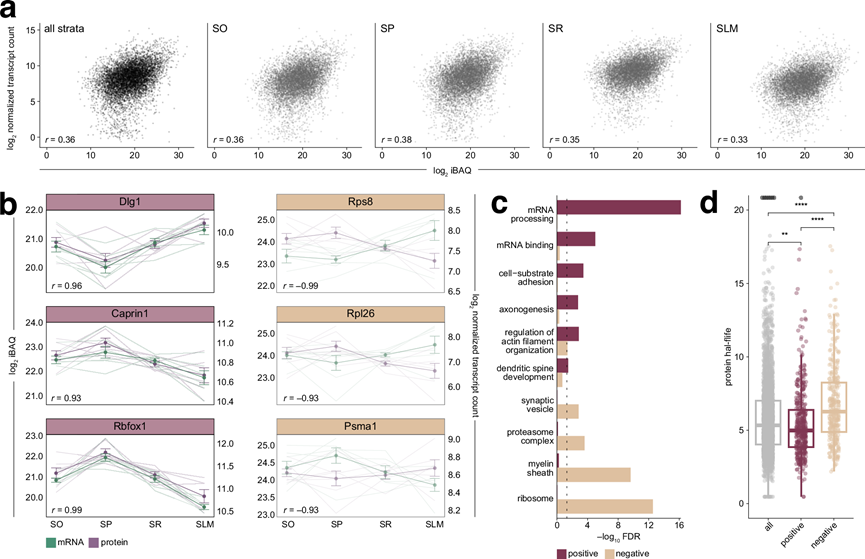
图4. 锥体神经元区室之间 mRNA 和蛋白质的相关性分析。
(a) CA1 层和每个层内mRNA 平均丰度及其各自蛋白质的 Pearson 相关性。(b) 选定的示例显示它们在各个层的平均丰度变化具有强正(≥0.9)和负(≤-0.9)Pearson 相关性。(c) GO分析。(d) 强正相关和负相关的mRNA-蛋白质对的蛋白质半衰期。
05
追踪CA1各层中mRNA的定位和蛋白质分布以及局部翻译的潜在热点
这些发现引出的一个核心问题是:mRNA水平和定位在多大程度上能够预测其编码蛋白在不同细胞区室的分布和丰度?以及mRNA与其对应蛋白的共富集在不同细胞区室间是否存在差异?为了探究这个问题,他们系统地评估了CA1各层内及各层间富集mRNA与其对应蛋白的重叠程度。该分析假设,mRNA和蛋白在同一细胞区室的共富集强烈提示该区室存在翻译活动;而mRNA在SP区的富集以及蛋白在其他任何细胞区室的富集则与细胞体中的合成和后续的转运过程相符。本研究的分析揭示了几个关键模式(图 5a),首先,SLM区的富集重叠程度最高,表明最远端的树突是局部翻译的热点区域。其次是SP区,其中共富集的基因主要与mRNA加工和核输出等过程相关。第三,SP 富集的 mRNA 也映射到 SLM 中富集的蛋白质,其中许多是膜蛋白,已知在运输之前会经历广泛的高尔基加工,例如受体和离子通道。
为了验证这些富集模式是否确实归因于局部合成而非转运,他们将本研究的数据集与公开发表的关于CA1胞体(SP)和神经毡(SR、SLM)翻译组的数据进行了比较。结果表明,在SLM中,45%的mRNA和蛋白质共富集的基因优先在神经毡中翻译,而只有6%主要在胞体中翻译(图 5b)。此外,65%的SP-SP共富集mRNA-蛋白质对和71%的SP-SLM共富集mRNA-蛋白质对优先在胞体中翻译,这表明这些mRNA在蛋白质转运之前就已经在SP中翻译(图 5c)。综上所述,这些发现突显了CA1中mRNA定位和蛋白质合成之间存在空间调控关系,不同的mRNA定位和翻译模式塑造了神经元区室中的蛋白质分布。
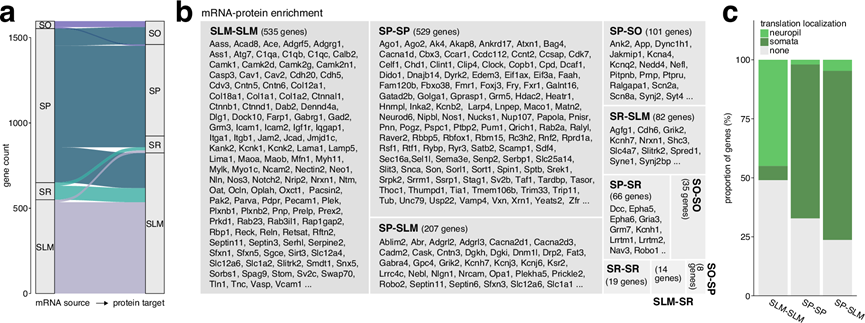
图5. 比较 CA1 层中的 mRNA 定位和蛋白质分布。
(a) 冲积图显示了不同 SO、SP、SR 和 SLM 组合中 mRNA 及其相应蛋白质的富集位点。(b) 平铺图显示了基于 mRNA-蛋白质富集的候选基因。(c) 根据其编码的蛋白质的富集位点,在体细胞或神经丛中优先翻译的 mRNA 比例。
06
海马亚区突触多样性是由不同的神经元群体和连接模式塑造的
为了更好地了解突触mRNA和蛋白质的空间组织,他们对海马组织和各层应用了突触纯化策略,使用泛突触Syn-1标记来捕获不同亚区和层中的突触,从而反映多种细胞类型的贡献。他们成功地从海马亚区(CA1、CA2/3、DG)(图 6a-b)和SO、SP、SR、SLM(图7a)中纯化了突触体。在过滤掉富集于前体组分中的残留污染物分子后,亚区域和层突触中定量蛋白质和检测到的 mRNA 转录本的数量分别总计超过 5000 个和 15000 个,重复实验之间具有很高的可重复性(图 6c和 图7b)。
为了确定海马体内的突触是否可以根据其转录组和蛋白质组的来源进行区分,他们应用了偏最小二乘判别分析(PLS-DA)。该分类方法成功地根据突触mRNA和蛋白质谱将CA1、CA2/3和DG区分开来(图 6d-e)。前三个主成分的载荷最高,代表了驱动这种区分的最重要特征,涵盖了一系列存在于兴奋性和抑制性突触(Gria3、Gabra3、 Gabrr3 )中突触前和突触后区室的分子(Arhgap32、Atg9a、Homer3、Shank1),这表明转录组和蛋白质组的变异性使海马体内的突触群体多样化。对载荷的进一步分析表明,特定的分子标记在区分某些亚区方面尤为重要。例如,当使用变量重要性投影(VIP)评分评估不同突触群的载荷效率时,他们发现突触后蛋白在区分CA1和CA2/3与DG中起关键作用。例如,Homer3在CA2/3区域的VIP评分最高,这归因于其在该区域的显著富集;而Shank1在DG区域的VIP评分较高,反映了其在该区域的相对缺失。随后,他们比较了所有转录本和蛋白质在各个亚区域的丰度,以鉴定具有显著表达模式的其他分子。抑制性回路的区域特异性特化表现为Gabrr3在CA2/3区域和Gabra1在CA1区域的显著富集。定义海马通路的分子特征也存在差异性富集。其中包括富含亮氨酸重复序列(LRR)的突触黏附分子,它们在组织突触特异性和多样性方面发挥着至关重要的作用。例如,他们发现Lrrtm4及其相互作用蛋白Gpc4的mRNA在海马CA1区富集,它们与从外侧内嗅皮层到齿状回颗粒细胞远端树突的突触模式形成有关。他们还观察到Lrrtm1在CA1区的富集,这与其在Schaffer侧支突触的突触后定位相一致。这些发现共同突显了海马亚区独特的分子特征,强调了回路特异性机制以及黏附分子在回路组织中的关键作用。
传统上,突触是根据神经递质类型进行分类的,但现在人们认识到,突触的分子组成更为多样化。他们假设突触的空间分布可能进一步加剧这种复杂性。CA1锥体神经元由来自海马内外的不同突触前输入组成。通过将对CA1的显微解剖与FASS(图7a-b)相结合,他们探究了这些汇聚于同一神经元群的突触是否存在差异。利用PLS-DA,他们发现这些突触的转录组具有足够的差异性,可以按层级进行区分(图 7c)。同一脑区内同一类型突触的蛋白质组数据高度相似,锥体神经元突触蛋白质组的分类也印证了这一点,PLS-DA 最清晰地区分了接收丘脑皮质输入的 SLM 突触与其他所有层级的突触(其中大部分输入来自海马内部)。接收 CA3 输入的 SR 突触仍能由部分区分,而 SO 和 SP 突触则相互交织。
通过提取PLS-DA分析中的主要载荷,他们进一步探究了哪些类型的分子对于定义这些空间受限的亚群最为关键(图 7d)。许多鉴定出的mRNA与激酶活性(Camk2b、Grk2)以及神经传递过程中的突触重塑(Cnih2、Gabbr3、Dock10)相关。相反,关键蛋白则参与细胞黏附(Afdn、Cadm2、Plxna4)以及细胞骨架的组织和运输(Actn1、Coro1a 、Coro2b、 Kif5c)。这些分子的VIP评分(表明它们区分不同突触群体的能力)也因层而异(图 7d)。例如,与其他层相比,Cnih2 mRNA或Actn1蛋白在SR层中表现出较高的VIP评分,这表明它们可能在该层的突触重塑中发挥关键作用。
为了进一步探究这些分子类别,他们检测了编码激酶和其他突触信号分子的关键转录本(图 7e)。他们发现Ca2+/钙调蛋白依赖性激酶、丝裂原活化蛋白激酶和丝氨酸/苏氨酸蛋白激酶的转录本以及编码突触小泡胞吐调节因子(RIMs)和支架蛋白(DOCK、DLGAP、DLG家族)的mRNA均呈现空间富集模式(图 7e)。先前对谷氨酸能突触体的研究表明,与神经递质释放相关的mRNA(例如RIMs)富集,他们在SLM中也观察到了类似的趋势(图 7e)。已知激酶在突触局部翻译,直接调节受体聚集和离子通道表达等过程¹³²,其差异表达会影响突触可塑性。他们发现Camk2n1、Mapk8ip1和Prkacb mRNA均优先定位于SLM突触,而Camk2a和Prkcb mRNA则富集于SR突触。这与先前关于它们在这些区域翻译的证据相符,并提示它们可能与这些层中发生的独特可塑性形式有关。
在突触蛋白质组中,一些黏附、轴突导向和神经元识别家族的成员成为其标志性特征。这些家族包括钙黏蛋白、神经连接蛋白、神经连接蛋白、Ephrin蛋白和Plexin蛋白,它们都参与突触的形成和重塑(图7e)。与激酶类似,这些蛋白质赋予突触可塑性。例如,N-钙黏蛋白(Cdh2,此处在SLM中富集)的内吞作用对于将突触活动与突触强度和结构的长期变化联系起来至关重要。Cdh6和Cdh9相互作用介导高强度的长时程增强作用,并影响蘑菇状棘突的密度,这种现象在SO中独有;他们观察到Cdh6和Cdh9在SO中富集的趋势(图 7e)。Ephrin蛋白,例如Epha4,在SR层中特异性富集,在SLM层中富集程度较低,它们可能在树突棘的组织和形成中发挥作用。一些具有富集趋势的分子也定位到支配连接处,例如SLM层中的Epha5,它参与建立内嗅皮层-海马连接。突触前黏附分子Nrxn1和Nrxn2也在SLM层中富集,这与其在投射到该层的丘脑神经元中的表达一致。有趣的是,他们还观察到肌动蛋白在整个数据集中的差异表达,其中F-肌动蛋白Actn1和Actn2在SR层中富集(图 7e)。先前的研究表明,树突棘处的肌动蛋白存在双向调控,强直刺激后F-肌动蛋白的表达会增加。他们纯化的突触体中驱动蛋白和微管蛋白的差异性富集也表明,特定突触的物质运输可能受到局部调控。Kif21a在SLM中的富集与该层GABA受体的运输和插入需求相对应,而Kif5a在SR中的富集可能与其在AMPA受体运输中的作用有关。综上所述,这些结果揭示了突触多样性的清晰空间特征,其特征在于CA1锥体神经元上突触前、突触后和跨突触分子的mRNA和蛋白质特征各不相同。
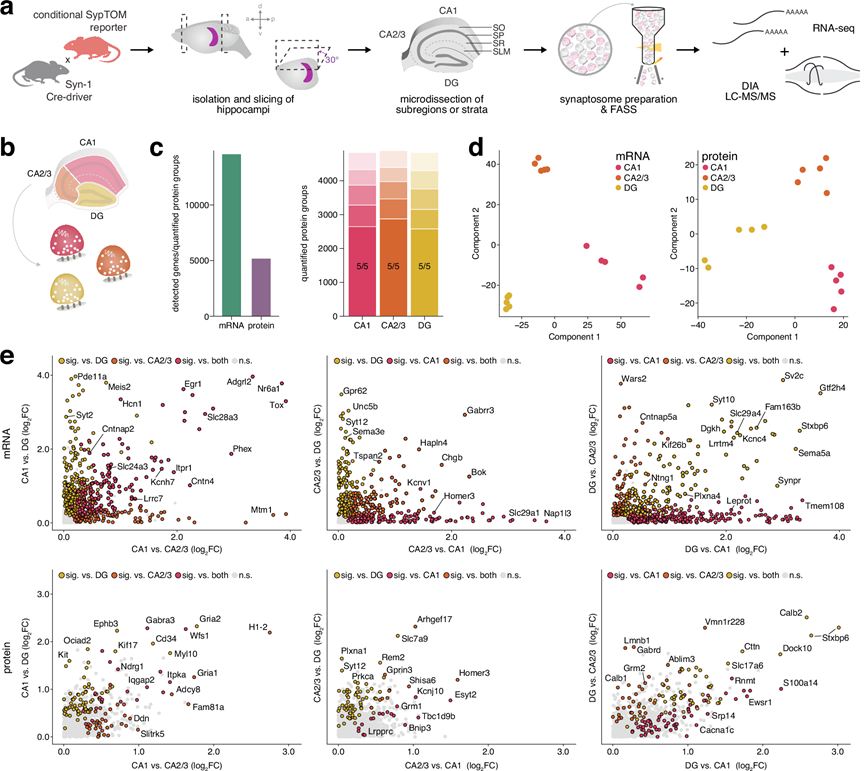
图6. 亚区域特定的神经元群体塑造海马中的突触多样性。
(a) 突触体分离实验流程。(b) 来自微切割亚区域的突触体示意图。(c) 左图:对每个亚区域进行污染物过滤后,在纯化的突触体中检测到的转录本和定量蛋白质组。右图:每个亚区域定量蛋白质组的有效值条形图。(d) PLS-DA 图显示转录组和蛋白质组按亚区域分离。(e) 转录本和蛋白质的散点图,每个亚区域的突触体均呈正向富集。
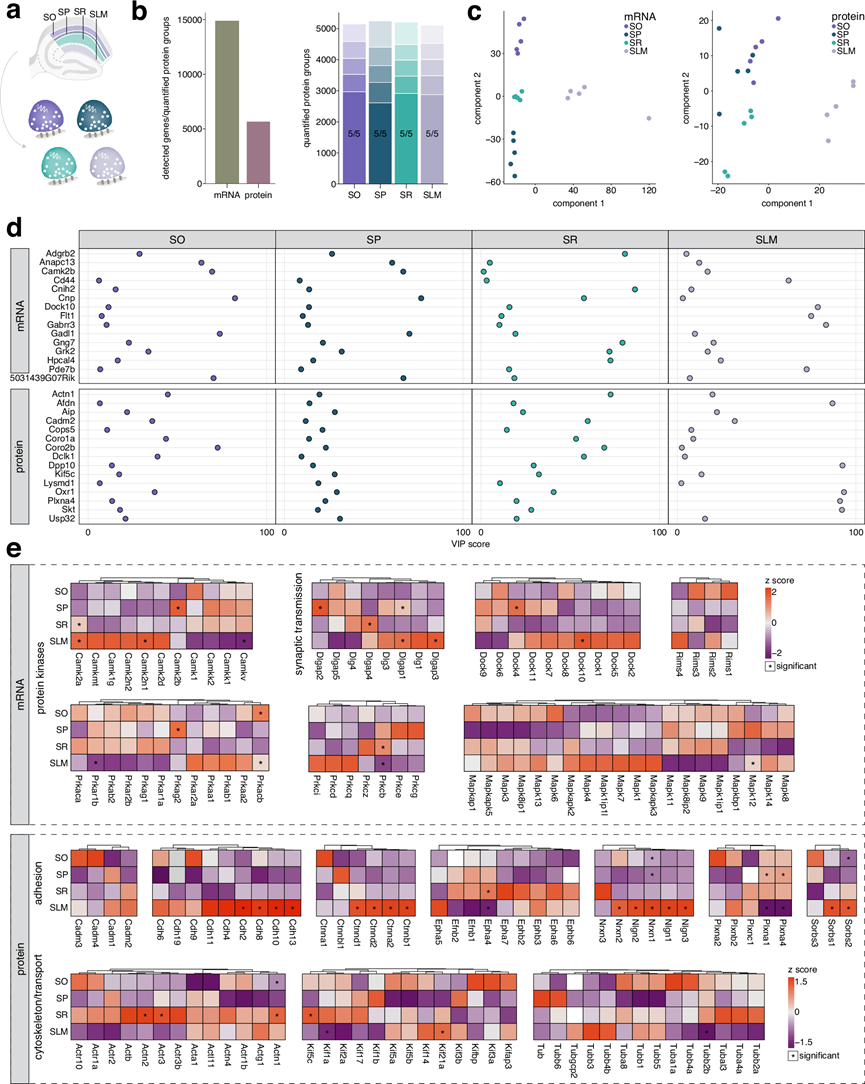
图7. 锥体神经元中隔室特异性突触 mRNA 和蛋白质特征的分类。
(a) 显微切割层的突触体示意图。(b) 左图:每个层经过污染物过滤后,在纯化的突触体中检测到的转录本和定量的蛋白质组。右图:每个层定量蛋白质组的有效值条形图。(c) PLS -DA 图显示转录组和蛋白质组的层分离。(d) 投影变量重要性 (VIP) 图显示 PLS-DA 中前三个成分的前五个负载的 VIP 分数。(e) 候选热图显示 CA1 神经元沿突触的 mRNA 和蛋白质表达模式。
+ + + + + + + + + + +
结 论
本研究对小鼠海马进行了深度空间分子分析,结合了对3个亚区和4个层的显微解剖、荧光激活突触体分选、转录组学和蛋白质组学数据,该方法揭示了数千种局部富集的分子,涵盖了多种受体、离子通道、代谢和黏附分子家族。转录组和蛋白质组数据的整合突显了与mRNA可用性紧密相关或不相关的蛋白质,部分原因是蛋白质半衰期的差异。多组学数据的整合揭示了蛋白质运输和局部翻译在锥体神经元区室化组织构建中的作用,其中远端树突对局部蛋白质合成的依赖性更高。CA1突触的分类揭示了激酶、细胞骨架成分和黏附分子在决定突触特异性方面所起的作用。这项研究共同提供了海马体及其突触的分子图谱,并揭示了空间转录-蛋白质关系。
+ + + + +



