

 English
English文献解读|Nat Metab(20.8):整合多组学揭示了在甲基丙二酰-CoA突变酶缺乏症中的代谢补偿性重组
✦ +
+
论文ID
原名:Integrated multi-omics reveals anaplerotic rewiring in methylmalonyl-CoA mutase deficiency
译名:整合多组学揭示了在甲基丙二酰-CoA突变酶缺乏症中的代谢补偿性重组
期刊:Nature Metabolism
影响因子:20.8
发表时间:2023.01.26
DOI号:10.1038/s42255-022-00720-8
背 景
甲基丙二酸尿症(MMA)是一种先天性代谢缺陷(IEM),具有多种单基因原因,且发病机制尚不清楚,导致缺乏有效的病因治疗方法。
实验设计

结 果
01

通过多组学进行单基因疾病变异检测
为了将对MMA的理解从致病的基因组病变扩展到受影响的生化过程,研究团队对230名个体(210名受MMA影响,20名未受影响)的成纤维细胞进行了高质量的全基因组测序(WGS)、转录组分析(RNA-seq)和基于diA - ms的蛋白质分型,这些个体主要来自欧洲,收集时间超过25年(图1a)。来自150名MMA患者的成纤维细胞的甲基丙二酰coa变位酶 /变位酶(MMUT)活性降低(MMUT缺陷),其中123名在补充辅助因子后没有增加,而60名患者的MMUT活性与对照组相似(其他MMA)(图1b)。在MMUT缺陷样本中,他们检索了WGS数据集,寻找MMUT基因的致病变异,并在150个人中确定了148人的分子致病原因。致病变异包括165个错义等位基因、105个截断等位基因、21个剪接等位基因、2个缺失等位基因和3个拷贝数变异,其中41个为新变异(图1c-d)。RNA-seq鉴定出与其他组相比,来自MMUT缺陷个体的细胞中MMUT RNA表达减少(图1e)。基于 DIA-MS 的蛋白质组检测显示,MMUT 缺陷的原代成纤维细胞中 MMUT 蛋白水平降低(图1f),分布在所有变体类型中。与所有其他样品相比,MMUT 代表了 MMUT 缺陷样品中最显著失调的 RNA 和蛋白质(图1g)。在剩下的60个样本中,他们在MMUT以外的基因中鉴定了22个个体的双等位致病变异:ACSF3、TCN2、sucl2和MMAB(图1h)。
他们发现210个受影响个体中有177个(84%)诊断为受影响个体(图1i),其中150个患有MMUT缺陷,19个患有ACSF3破坏性变体,占ACSF3缺陷的绝大部分。

图1. 多方面组学视图使 84% 的个体能够进行分子诊断。
(a) 研究概述,描述了丙酸途径,包括其前体和 MMUT 催化的途径。(b) 每个研究子队列的 MMUT 酶活性。(c) MUT基因上发现的所有致病变异的棒棒糖图。(d) MMUT基因上识别的变异类型的比例。(e-f) 按研究子队列划分的 MMUT 转录本和蛋白质水平。(g) 基因根据蛋白质组学和转录组学数据中由Welch 's t检验(双侧)计算的P值进行排序。(h) ACSF3中鉴定的致病变异的棒棒糖图。(i) 整个队列中确定的受影响基因的比例。
02
表型描述及其与疾病严重程度的关系
在排除了9个非特异性和/或相互依赖的特征后,他们生成了一个表型变量的相关矩阵,涵盖了出现时和疾病过程中的临床症状、临床治疗和治疗反应、血液或组织的临床化学(包括代谢物测量和体外生化参数),揭示了一组特征(MMUT活性、PI),这些特征在许多变量之间显示出很强的相关性(图2a-b)。
随后他们建立了临床严重程度评分(CSS),纳入了五个典型临床特征的结果,其中0分表示没有这些典型的MMA特征,1分表示轻度MMA, 2分或更高(最大5分)表示中度至重度MMA疾病。CSS与所有表型参数的比较表明,与49个个体变量存在显著相关性(图2c),包括MMA的许多经典表型症状,如酸中毒、高氨血症和肌肉张力低下,以及对饮食和药物干预的需求。
多重相关分析发现,在羟钴胺素(PI+)存在的情况下,PI与42个表型特征显著相关,是所有个体连续变量中相关性最大的(图2c)。例如,这与发病年龄形成对比,后者仅与16个参数显著相关(图2c)。进一步检测发现PI+与疾病严重程度呈负相关,包括与pH或发病年龄显著正相关,与血浆中甲基丙二酸浓度、存在临床干预措施负相关(图2d)。

图2. 型组学分析揭示了疾病严重程度的两个主要替代标志物(CSS 和 PI+ 活性)。
(a) 所有连续数值和离散表型变量的相关矩阵。(b) 根据五个表型子类别的表型性状数量。(c) 选定表型性状的面板及其代表整个表型组数据集(此处称为疾病严重程度)的整体强度,通过对数转换后的线性模型进行评估。 (d) PI+活性变量与其余表型变量相比的线性回归结果。
03
TCA循环和相关途径的破坏
为了确定与疾病相关的基因、蛋白质和途径的表达改变,他们尝试与上面确定的定量表型变量相结合,对转录本和蛋白质表达进行全面评估。由于 TCN2、SUCLA2 和 ACSF3 缺陷的患者缺乏经典 MMA 的大多数典型体征和症状,因此他们将 MMUT 缺陷患者与所有非 MMUT 缺陷样本(对照)进行比较。多组学因素分析,整合了遗传数据层和蛋白质分型数据,确定了线粒体代谢途径,特别是,电子传递链和三羧酸(TCA)循环在缺乏mutt的样品中发生富集(图3a)。更详细地说,在tca相关基因集中的蛋白质分型数据中,SLC16A3、CS、MDH2和OGDH蛋白是该特定因子变异的主要驱动因素(图3b)。转录组学和蛋白分型之间共有的基因的线性判别分析表明,MMUT是最强的,sucl2、OGDH和PDHB是MMUT缺陷和对照样品分离的主要驱动因素(图3c)。
此外,通过CSS和PI+根据疾病严重程度进行样本分层的基因集富集分析也发现,氧化磷酸化和TCA循环在蛋白质组学(CSS和PI+)和转录组学(CSS)数据集中是过度代表的途径(图3d)。这些变化与MMA半合子小鼠模型的发现一致(图3e)。突变动物的大脑、心脏、肾脏、肝脏、血浆和尿液的非靶向代谢组学证实,突变动物体内的甲基丙二酸代谢产物水平升高,而通路富集分析则指出,所有组织和尿液中的TCA循环途径失调(图3f)。脑组织转录组学进一步证实突变小鼠的Mmut转录减少了50%,同时电子传递链和氧化磷酸化途径富集(图3g)。

图3. 组学数据层的非针对性整合突出显示了 TCA 循环和相关途径以及 MMA 中失调的氧化磷酸化基因集。
(a) 使用多组学因子分析工具(MOFA)进行基因集富集分析。(b) 蛋白质组数据中MOFA之后的顶级富集基因集的详细特征统计。(c) 转录本的线性判别模型将 MMUT 缺陷与由MMUT和与 TCA 循环相关的其他基因驱动的控制分开。(d) 基于差异表达分析得出的ES排序的基因集富集分析。(e) Mmut缺陷小鼠的饲养方案。(f) 小鼠组织和体液中的非靶向代谢组学。(g) 小鼠脑组织的RNA-seq。
04
MMUT缺乏导致近端TCA酶的改变
由于数据驱动和表型分层分析都表明,TCA和相关途径在疾病中发生破坏,他们对TCA循环酶进行了协同调查,包括那些补充TCA循环中间体和去除TCA循环中间体反应的酶,从中他们获得了RNA和蛋白质信息。作为对照,他们纳入了不参与这些途径的TCA酶的异构体(图4a)。直接比较MMUT缺陷细胞和对照细胞之间的RNA和蛋白质表达,发现MMUT明显失调(图4a)。
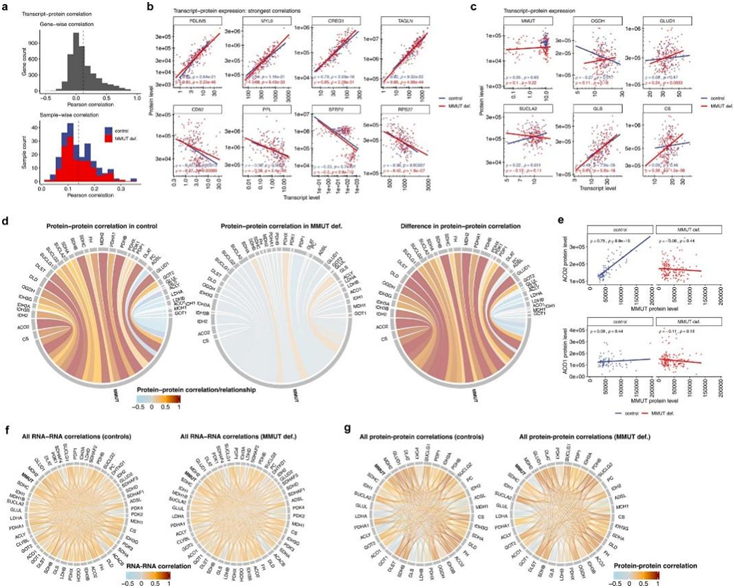
MMUT在RNA和蛋白质水平上在疾病中都显著下调,而催化甲基丙二酸和甲基丙二酸半醛之间交换的ALDH2在两者中都上调(图4a-b)。进一步上调的转录本是PDK4(图4b),它负责磷酸化,并因此导致丙酮酸脱氢酶复合物失活;然而,总体效应最大的蛋白是OGDH(在疾病中下调)和GLUD1(在疾病中上调),这两种酶都参与了谷氨酰胺的逆转(图4b)。与研究结果一致,其他人对人类MMA肝脏的蛋白质组学研究同样发现了GLUD1和ALDH2的上调,并且对小鼠MMA模型中肝脏来源的分离线粒体的靶向分析显示OGDH蛋白水平和酶活性降低。对所有样本中rna -蛋白表达相关性的检测显示,基因水平(4318对转录蛋白对)的Spearman相关性为0.14,样本水平的Spearman相关性为0.40,与其他人的发现相似(图4c,图S5a)。与对照组相比,mmut缺陷细胞的rna -蛋白相关性显示,两种基因型中有1158对显著相关(图4d,图S5b)。其中,OGDH、GLUD1、CS和GLS在mmut缺失样本中的rna -蛋白相关性高于对照组,而sucl2的rna-蛋白相关性降低(图4a-d,图5c)。OGDH和sucl2是基因型依赖性rna -蛋白相关变化最强的基因(图4d)。值得注意的是,他们发现在对照细胞和MMUT缺陷细胞中,MMUT的rna -蛋白相关性都很差(图4a-d,图5c)。
最后,在对照组中,MMUT蛋白水平与许多TCA和补偿性酶的蛋白水平呈正相关,而在MMUT缺陷细胞中则没有,而在两种基因型中,MMUT和非TCA蛋白亚型之间几乎没有蛋白表达相关性(图4a,图5Sd)。这种关系在MMUT:ACO2和MMUT:ACO1中得到了体现(图4a,图S5e),这表明MMUT可能是迄今未知的与这些线粒体TCA循环和补偿性酶相互作用网络的一部分。对这些通路中所有蛋白质和转录本之间的配对相关的检验(图S5f-g)表明,TCA循环和补偿性酶彼此呈正相关,总之,上述研究结果表明,MMUT RNA和蛋白表达的破坏驱动了某些TCA和补偿性酶的调节变化。

图4. 转录本-蛋白质和蛋白质-蛋白质相关性分析揭示了 MMUT 和 TCA 基因和蛋白质之间的协调关系,但不是它们的亚型。
(a) Circos 图描绘了转录本和蛋白质的差异倍数 (FC)、差异表达分析得出的效应大小 (ES)、转录本-蛋白质相关性 (rho) 以及 MMUT 蛋白质与 TCA 蛋白质及其相应亚型的相关关系。 (b) Q-Q和火山图说明了基于线性混合建模方法的差异表达分析的结果。(c) 4318个转录蛋白对(左)和221个样本(右)的Spearman相关性直方图。 (d) MMUT 缺陷与对照的 Spearman 相关性散点图。
图S5. 转录蛋白和蛋白-蛋白相关分析。
(a) 4318个转录蛋白对(上)和221个样本(下)的Pearson相关性直方图。(b) 转录本-蛋白质最强正相关和负相关的散点图(按mmut缺陷组和对照组Pearson相关系数的平均值排序)。 (c) TCA循环相关基因的转录蛋白Pearson相关图。(d) 对照(左)、MMUT缺陷样本(中)的弦图显示了MMUT蛋白与TCA循环和相关蛋白及其亚型的选择的Spearman相关性,以及前两个图的差异(右)。(e) MMUT蛋白与顺乌头酸酶两种亚型(ACO1和ACO2)的散点图,Pearson相关线性回归。(f) 和弦图显示了TCA循环和相关转录本或蛋白质的所有相关关系。

图S5. 转录本-蛋白质和蛋白质-蛋白质相关性分析说明了 MMUT 与大多数 TCA 转录本和蛋白质的协调调节。
(a) 4318 个转录本-蛋白质对(上)和 221 个样本(下)的 Pearson 相关性直方图。(b) 最强的正向和负向转录本-蛋白质相关性的散点图。(c) 所选TCA循环相关基因的转录本-蛋白质Pearson相关图。(d) MMUT 蛋白与选择的 TCA 循环和相关蛋白及其亚型的 Spearman 相关性。 (e) MMUT蛋白与乌头酸酶两种亚型(ACO1和ACO2)的散点图。(f-g) 和弦图显示了TCA循环和相关转录本或蛋白质的所有相关关系。
05
代谢组学突显TCA循环补偿性代谢的重组
为了检查上述 RNA 和蛋白质表达变化的功能后果,他们对一组来自患者的 6 个 MMUT 缺陷的成纤维细胞和来自未受影响个体的 6 个对照原代成纤维细胞进行了非靶向代谢组学分析。虽然总离子电流在mutt缺陷和对照样品中相当,但他们发现谷氨酰胺和丙氨酸的减少以及己糖、甲基柠檬酸、氧己酸、氨基己酸和丙酮酸的增加是变化最显著的代谢物(图5a)。逆转代谢物的变化与匹配模型系统中观察到的RNA和蛋白质表达的变化一致,表明MMUT缺乏症中TCA循环逆转发生破坏(图5b)。
为了深入研究突变改变,他们对具有野生型(WT)、MMUT敲除(KO)或dst -KO遗传背景的293T细胞进行了靶向代谢组学研究,并通过Western blotting分析和酶活性测量进行了验证。dst-ko细胞用作模拟MMUT缺乏症中OGDH蛋白减少的额外对照。在mutt-KO细胞中,他们发现TCA循环中间体明显减少,表明TCA代谢物含量总体减少,而DLST的KO导致大多数TCA循环代谢物几乎不存在(图5c)。这些结果表明疾病的调节依赖于谷氨酰胺代偿性途径;他们通过评估[U-13C]谷氨酰胺在TCA循环和相关中间体中的相对碳掺入量进一步验证了这一假设(图5d)。在本实验中,谷氨酰胺的代偿性代谢产生M + 5(谷氨酸和氧戊二酸盐)和M + 4(琥珀酸盐、富马酸盐、苹果酸盐和柠檬酸盐)化合物,而TCA循环的氧化循环将吸收大部分未标记的碳(例如来自葡萄糖和甲基丙二酰辅酶a)和谷氨酰胺衍生的13C。在mutt-ko条件下,他们发现所有研究的TCA循环代谢物中M + 5氧戊二酸和M + 4同位素的比例分数有增加的趋势,而M + 0的比例分数则有所降低(图5e)。这表明MMUT受损的细胞增加了谷氨酰胺的使用,作为一种代偿性过程。此外,与OGDH活性降低相一致的是,在MMUT缺乏的情况下,柠檬酸盐的M + 5/M + 4比值增加,表明了还原性TCA循环途径的相对偏好(图5d-f)。
为了检测直接补充核心TCA相关代谢物是否可以弥补TCA循环中间产物的减少,他们在没有干预的情况下或补充柠檬酸盐、苹果酸盐或二甲基氧葡萄糖酸盐后,在293T细胞和成纤维细胞中重新进行了代谢组学研究(图5)。二甲基-氧戊二酸是2-氧戊二酸的膜渗透性替代品,以前用于OXPHOS功能障碍模型。补充柠檬酸盐和苹果酸盐增加了细胞内代谢物的数量,但对其他TCA中间体没有广泛的影响(图5g)。相比之下,二甲基-氧戊二酸除了增加氧戊二酸库外,还增加了293T细胞中的琥珀酸盐、苹果酸盐和富马酸盐以及原发患者成纤维细胞中的柠檬酸盐(图5g)。另外两个观察结果使得二甲基-氧戊二酸盐成为一种潜在的治疗化合物:在患者成纤维细胞中,二甲基-氧戊二酸盐强烈减少丙酰肉碱含量,丙酰肉碱是MMA疾病的生物标志物和有毒代谢物丙酰辅酶a的衍生物(图5h),并且它导致MMUT缺乏症中柠檬酸盐的M + 5/M + 4比率降低的趋势(图5i),表明在这种治疗下可能抑制和重新平衡还原性TCA循环。

图5. CRISPR/Cas9 KO 293T 细胞和原代患者成纤维细胞的极性代谢组学和谷氨酰胺示踪研究突出了差异性谷氨酰胺重组。
(a) 差异表达代谢物的火山图。 (b) TCA循环和相关重组的示意图。(c) 对照和 CRISPR/Cas9 KO 293T 细胞中的代谢物。(d) TCA循环标记和通过回补衍生自标记谷氨酰胺的相关代谢物的示意图。(e) 谷氨酰胺标记后TCA循环代谢物同位素异构体的相对丰度。(f) M + 5 和 M + 4 柠檬酸盐同位素同源物的比率。(g) 不同处理条件下TCA循环代谢物的总库大小。(h) 接受治疗的原代患者成纤维细胞中丙酰肉碱的水平。(i) 在原代成纤维细胞中处理下的M+5和M+4柠檬酸盐同位素同源物的比率。
06
MMUT 与其他代偿性酶发生物理相互作用
上述观察到的谷氨酰胺回补调整的调节病因尚不清楚。基于 MMUT 与近端 TCA 循环和代偿性酶之间的强蛋白质表达相关性(图4a,图S5d),他们假设这些蛋白质可能是代谢复合物的一部分,可能促进 TCA 循环的调节蛋白质-蛋白质相互作用导致谷氨酰胺回补。
为了深入了解这种潜在的物理关系,他们利用了293T细胞中c端flag标记的功能性MMUT版本的过表达,MMUT辅助因子(MMAA和MMAB)和底物(MCEE)合成的三个途径成员参与任何含有MMUT的多蛋白复合物,以及三个阴性对照[空载体(EV)、VLCAD和ACO2)](图6a)。他们对四种 MMUT 途径蛋白(MMUT、MMAA、MMAB 和 MCEE)和两个阴性对照(EV 和 VLCAD)进行了 IP 联用质谱分析。他们鉴定了37个由MCEE、MMAA、MMAB和MMUT拉下而不由EV或VLCAD拉下的靶标蛋白,包括MMUT、OGDH、DLST和GOT2,以及20个由MCEE、MMAB和MMUT拉下而不由EV或VLCAD拉下的蛋白质,包括MMAB和GLUD1(图6b)。对MMUT与EV和VLCAD的生物重复进行方差分析(ANOVA),发现MMUT样品中有22个蛋白显著富集(图6c)。所有的蛋白都由UniProt标记为具有线粒体定位,包括GLUD1、GOT2和DLST(图6c)。方差分析比较了MMUT、MMAA、MMAB和MCEE中至少三种蛋白的重叠,但EV或VLCAD均未鉴定出11种相互作用蛋白,包括GLUD1和GOT2;而MMUT、MMAA、MMAB和MCEE两种阴性对照的交叉鉴定出包括DLST在内的13种相互作用蛋白(图6d)。最后,通过免疫印迹进一步证实MMUT和DLST之间形成复合物(图6e)。这些数据表明MMUT是近端代谢酶复合物的一部分,包括GLUD1和氧戊二酸脱氢酶复合物组分DLST,并表明这些相互作用的破坏可能是它们在疾病中的调节改变的基础。

图6. MMUT 与 GLUD1、DLST 和 GOT2的物理相互作用分析。
(a) 实验组和对照组的轮廓,表明在交联亲和纯化实验中哪种蛋白质与FLAG标签一起使用,并随后通过质谱法对pull-down样品进行分析。 (b-c) 由不同 FLAG 标记的蛋白质pull-down的蛋白质的维恩图。(d) 显著富集的蛋白质的相互作用网络。
+ + + + + + + + + + +
结 论
本项研究采用多组学技术结合 MMA 患者的生化和临床特征,对177 例患者样本进行分析,其中大多数样本表现出甲MMUT的致病变异特征。通过对小鼠模型的多器官代谢组学分析以及通过识别 MMUT 和谷氨酰胺补充酶之间的物理相互作用,可以证明这些代谢紊乱的相关性。使用稳定同位素示踪,发现用二甲基酮戊二酸治疗可以恢复缺陷的三羧酸循环功能,本项研究强调了谷氨酰胺补充是 MMA 中潜在的治疗治疗靶点。
+ + + + +



