

 English
English文献解读|Nat Commun(16.6):肾脏转录组、蛋白质组和多组学的遗传分析阐明了新的血压和高血压靶点
✦ +
+
论文ID
原名:Genetic imputation of kidney transcriptome, proteome and multi-omics illuminates new blood pressure and hypertension targets
译名:肾脏转录组、蛋白质组和多组学的遗传分析阐明了新的血压和高血压靶点
期刊:Nature Communications
影响因子:16.6
发表时间:2024.03.19
DOI号:10.1038/s41467-024-46132-y
背 景
持续升高的血压 (BP)(高血压)是全球死亡的最重要的单一危险因素,BP调节的遗传机制仍然不明确。
实验设计
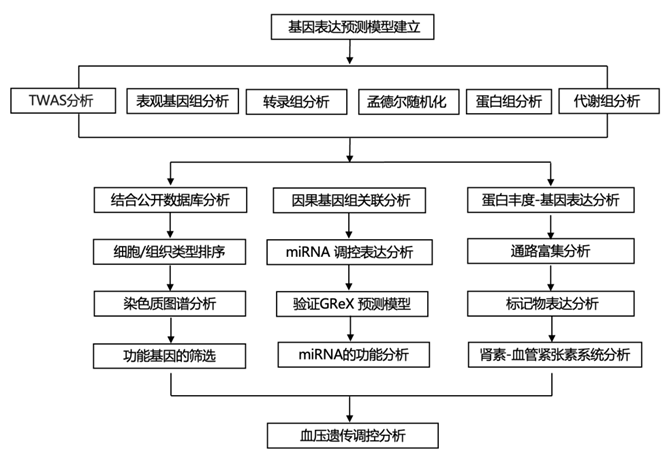
结 果
01

通过全转录组关联研究对人类细胞类型/组织与血压的相关性进行优先排序
全转录组关联研究 (TWAS) 不仅可以揭示细胞类型/组织对性状/疾病的生物学重要性,还可以揭示新的和意想不到的组织-疾病关联,例如肠道与精神疾病或单核细胞与帕金森病的关联。研究团队应用弹性网方法来预测基因型组织表达 (GTEx) 中数值相同的个体组中 49 种人类细胞类型和组织的基因表达的遗传调控成分,使用 S-PrediXcan 来估计各种 GTEx 组织中基因表达水平对收缩压 (SBP) 和舒张压 (DBP) 的调控作用。然后,他们计算了每种人类细胞类型/组织的整体 SBP 和 DBP TWAS 分数,根据它们与 SBP 和 DBP 遗传调控的相关性对其进行排序(图1A,图2A)。
培养的成纤维细胞和肾皮质分别显示出与 SBP 和 DBP 最强的总体关联(图2A)。在收缩压和舒张压的前10位组织中,只有5种细胞/组织——肾脏、培养的成纤维细胞、肾上腺、EBV转化的淋巴细胞和甲状腺(图2A)。血管系统(主动脉、胫动脉)和中枢神经系统(包括海马、皮质、小脑半球、额叶皮质、小脑和尾状基底神经节在内的几个大脑区域)在与BP最高相关的组织中都有很强的代表性(图2A)。淋巴细胞、脾脏和小唾液腺之间的关联支持免疫系统在血压调节和高血压中的新作用,而大脑区域与血压之间的联系不仅在高血压的发病机制中,而且在高血压介导的认知功能下降中。
他们应用染色质构象和表观基因组学模型进行预测 (PUMICE) ,这是一种集成了 3D 染色质组织数据和表观遗传注释的算法,以生成人类肾脏中遗传调控基因表达 (GReX) 的预测模型(图 1B)。为了获得 3D 肾脏基因组数据,他们首先使用 HK2 细胞系中的 H3K27ac Hi-ChIP 建立的流程生成高分辨率的以转录为中心的染色质图谱(图 2B)。总共,他们获得了 69036737 个基因组位点,并鉴定了 11569 个拓扑关联域 (TAD) 和 8934 个染色质环(图 2B)。对于表观基因组学数据,他们使用4个不同的轨道从ENCODE检索人类成年肾脏的信息,包括H3K27ac(127379个峰)、H3K4me3(70569个峰),DNA酶超敏位点(275004个峰)和CTCF(45832个峰)(图2B)。
通过使用PUMICE将这些数据与发现资源中478个肾脏的RNA测序衍生转录组整合(图1B),他们生成了8682个重要的基因表达预测模型(图2C)。然后,他们在作为验证资源收集的 222 个肾脏的独立数据集中验证了这些模型(图 1C)。内质网氨肽酶2 (ERAP2)基因是肾脏基因,在发现和验证资源中,其预测的GReX和观察到的表达之间有很好的相关性(图2D-E)。总体而言,在发现和验证肾脏资源之间,PUMICE衍生模型的预测性能有很强的相关性(图2F)。在三个主要基因生物型之间,肾脏的基因比例无显著差异(图2G)。
除了验证预测模型的计算稳健性之外,他们还试图检测它们的估算基因表达是否与其肾脏中分子活性的生化读数相关。在具有强大、独立验证的 GReX 的肾脏基因中,他们选择了:溶质载体家族 2 成员 9 基因 (SLC2A9)、溶质载体家族 22 成员 12 基因 (SLC22A12)、溶质载体家族 2 成员 2 基因 (SLC2A2)、肌醇六磷酸激酶 3 基因(IP6K3)、钙敏感受体基因(CASR)分别作为肾小管上皮中尿酸盐、葡萄糖、磷酸盐和钙转运调节因子的例子(图 2H)。他们证明,在英国生物银行(UK Biobank)的321210名个体中,通过基因推断的这些基因在肾脏的表达显示出与相关血液生化表型的预期关联;例如,SLC22A12和SLC2A9的肾脏表达增加(分别编码顶膜URAT1和基底侧GLUT9——肾小管近端尿酸重吸收的主要转运蛋白)与血清尿酸水平的增加相关(图2H)。SLC22A12和SLC2A9是近端肾小管重吸收尿酸的主要转运蛋白。
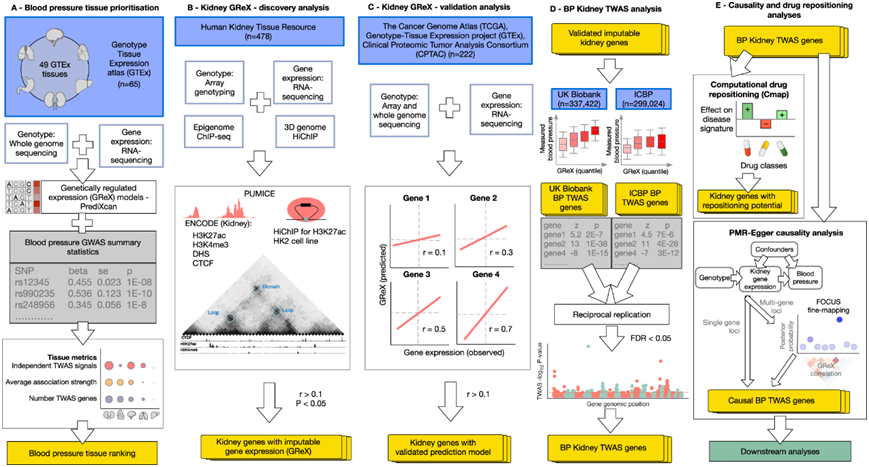
图1. 全转录组关联研究、肾脏和血压、分析过程和输出数据的示意图。
(a) 血压组织优先顺序。 (b) 肾 GReX 通过使用染色质构象和表观基因组学 (PUMICE) 算法进行预测-发现分析得出。(c) Kidney GReX -验证分析。 (d) BP 肾脏 TWAS 分析。(e) 因果关系和药物重新定位分析。
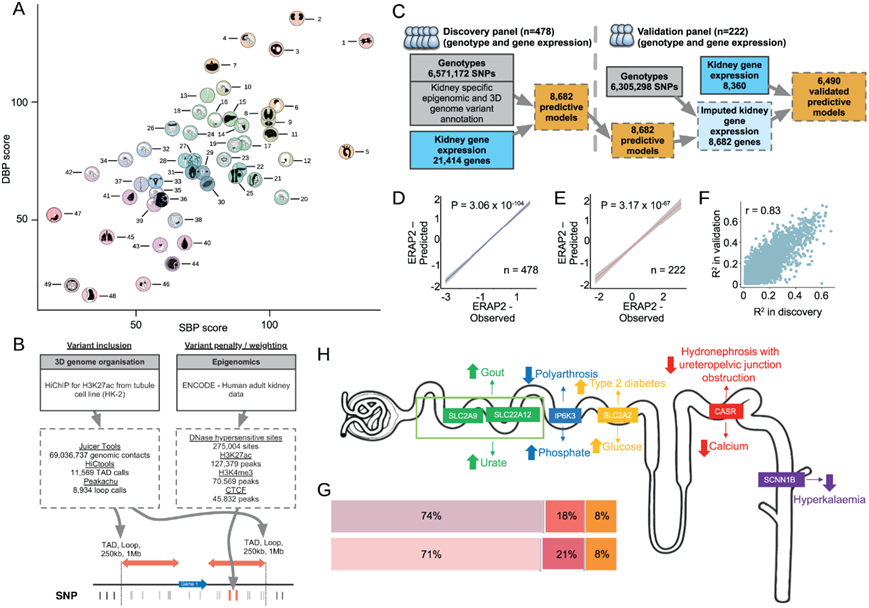
图2. 血压 TWAS – 从人体组织的优先顺序到增强基因表达预测肾脏模型的开发。
(A) GTEx 项目中 49 种人体组织和细胞类型的代表。 (b) 增强的基因表达预测。 (c) TWAS 模型预测工作流程。(d-e) HKTR(蓝色)和 NIH 资源(红色)中预测的基因调控表达 (GReX) 与观察到的ERAP2表达之间的相关性。(f) 来自发现资源与验证资源的基因表达模型的预测性能。(g) 生物型中可推定基因(上)和所有表达基因(下)的百分比。(h) 与溶质细胞转运相关的肾脏基因。
02
孟德尔随机化和因果基因组分析
他们的孟德尔随机化(MR)研究随后对因果基因集进行了精细定位,优先考虑了 399 个肾脏基因,显示出对 BP 的潜在因果影响(图 3)。近一半(182,45.6%)作为单个基因映射到特定的 BP GWAS/TWAS 位点,其中 29 个对所有三个 BP 性状都有因果影响(图 3)。
他们根据这399个基因已知的功能及其与人类健康和疾病的相关性,将它们分配到16个生物学主要主题中,观察到在BP肾脏组织特异性关联分析基因中有非常强烈的人类代谢足迹(图3)。这包括负责氨基酸[例如,胍基酶基因(AGMAT)和丝氨酸消旋酶基因(SRR)]、碳水化合物[例如,溶质载体家族5成员11基因(SLC5A11)、溶质载体家族2(促进葡萄糖转运)、成员4基因(SLC2A4)和淀粉结合域1基因(STBD1)]、脂类[例如,酰基辅酶A硫酯酶8基因(ACOT8)、ELOVL脂肪酸延长酶7基因(ELOVL7)和脂肪酸去饱和酶1(FADS1)]、维生素[例如,叶酸受体α基因(FOLR1)]和氧化磷酸化[例如,内膜线粒体蛋白基因(IMMT)和烟酰胺核苷酸转氢酶基因(NNT)]相关通路。这些基因在维持钠-水平衡的生理机制中具有重要作用,例如,水通道蛋白1基因(AQP1)和水通道蛋白4基因(AQP4)的肾脏表达增加—这两种水选择性通道分别在近端肾小管/亨利环的细长下降肢/下降的直小血管和集合管的主细胞中起作用,显示了对收缩压和舒张压增加的因果效应(图3)。
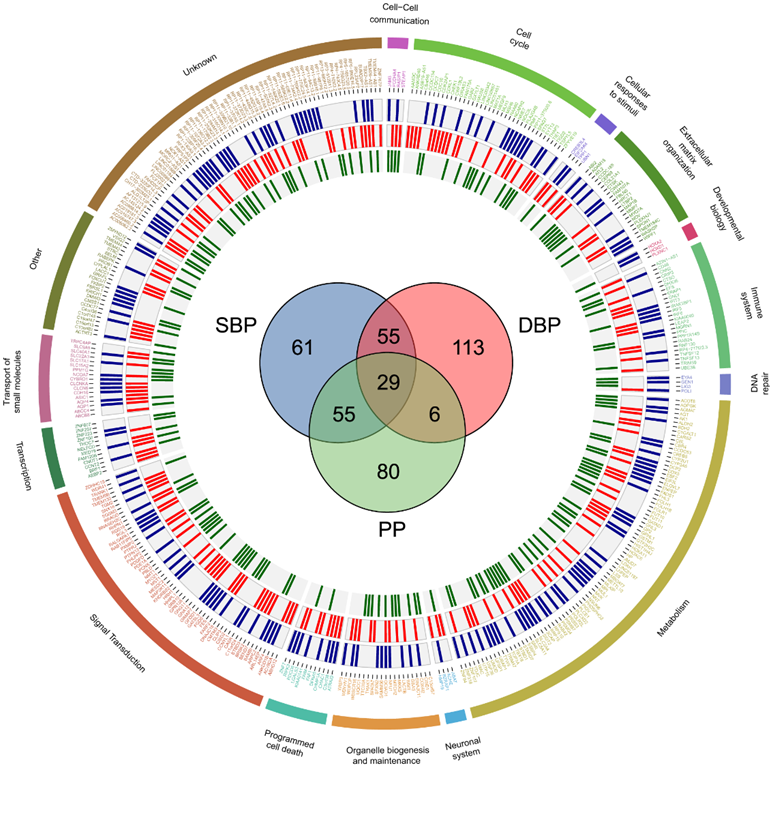
图3. 圆形表示 399 个假定的血压致病基因的信息以及它们与 SBP(蓝色)、DBP(红色)和 PP(绿色)的共同关联程度。
03
将血浆蛋白质组学和代谢组学的输出与肾脏转录组范围的关联研究进行三角测量
血浆蛋白质组学和代谢组学的遗传分析越来越多地用于深入了解复杂特征的发病机制,包括慢性肾病 (CKD) 和高血压。他们试图以SLC5A11、胍丁胺酶基因 ( AGMAT ) 和血管紧张素原基因 ( AGT ) 为例,探索这些组学层次如何帮助对肾脏 TWAS 结果进行功能分析(图 4)。
他们发现SLC5A11肾脏表达降低与PP升高有因果关系(图4A-B)。该基因编码钠/葡萄糖协同转运蛋白6 (SGLT6, SMIT2)。利用来自两个队列(INTERVAL和EPIC-Norfolk)的14296名个体的数据,他们证实肾脏SLC5A11确实与循环中的肌醇水平相关(图4C)。然后,他们发现肌醇和PP之间存在负相关关系(图4C)。通过进一步的中介分析,他们确定SLC5A11肾表达对 PP 的影响约 48.4% 是由血清肌醇水平介导的。
BP 肾脏 TWAS 还证明,基因决定的AGMAT肾脏表达增加与 SBP 增加存在因果关系(图 4D-E)。AGMAT负责在多胺生物合成替代通路上,将精氨酸下游的胍丁胺转化为腐胺(图4D)。他们还证实了肾脏AGMAT与4-胍丁酸(也称为γ -胍丁酸或γ -胍丁酸)血浆水平之间的关联,其唾液水平与AGMAT有关(图4F)。他们还发现血压与4-胍丁酸循环水平之间存在关联(图4F)。最后,通过进一步的中介分析,他们确定肾脏AGMAT表达对收血压的影响约12.3%是由血浆中4-胍丁酸水平升高介导的(图4F)。
通过TWAS还发现肾脏血管紧张素原基因(AGT)表达增加与血压之间存在强烈的因果关系(图4G-H)。血管紧张素原mRNA的表达通过肾内RAS活性影响血压(图4G),他们试图量化检测到的效应在多大程度上确实独立于全身(循环)血管紧张素原。利用Fenland研究中10708人的数据和现有的血浆血管紧张素原蛋白水平,他们首先证实了血浆血管紧张素原与BP之间的因果关系在预期方向上(图4I)。进一步的中介分析表明,大约44.7%的肾脏AGT mRNA对血压的影响与循环血浆血管紧张素原无关(图4I),可能反映了肾脏血管紧张素原的局部活性。
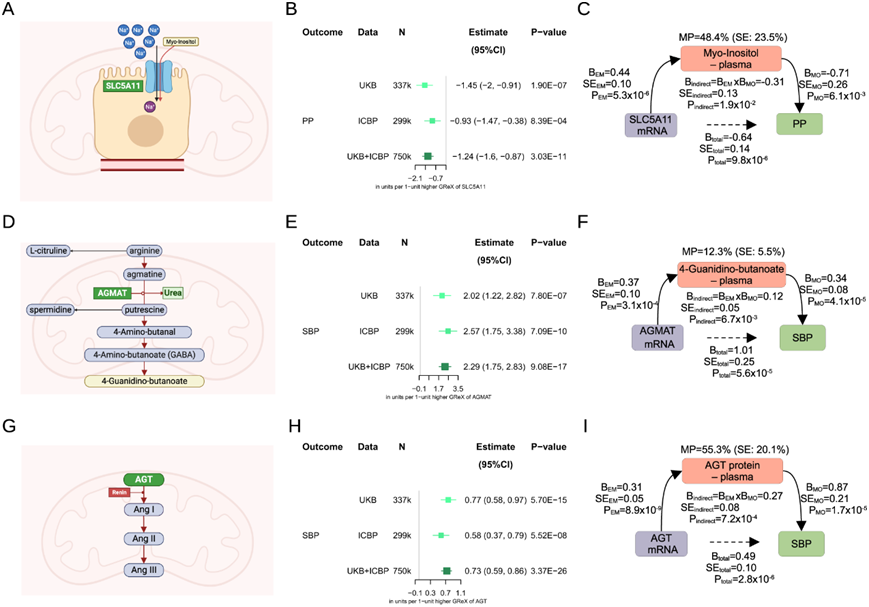
图4. 肾脏基因的基因预测表达及其生化读数-综合多组学分析。
(a) SLC5A11编码的钠/肌醇共转运蛋白2 (SMIT2)在肾近端小管细胞顶部的定位。 (b) 全转录组关联研究 (TWAS) 中肾脏SLC5A11的基因调控表达 (GReX)对脉压 (PP) 的影响。(c) 表示SLC5A11 mRNA表达对由血浆肌醇水平介导的PP的58.4%影响。(d) 精氨酸分解代谢的多胺通路。(e) 肾脏AGMAT (GReX) 对 TWAS 收缩压 (SBP) 的影响。(f) 表示AGMAT mRNA 表达对 4-胍基丁酸血浆水平介导的 SBP的 11.7% 影响。(g) 肾素催化从ANG (血管紧张素原)到血管紧张素 I (AngI)的反应,随后通过血管紧张素转换酶 (ACE) 将其转化为 AngII。(h) 肾脏AGT mRNA (GReX) 对 TWAS 中 SBP 的影响。(i) 表示AGT mRNA 表达对血管紧张素原循环血浆蛋白水平介导的 SBP产生 52.7% 的影响。
04
肾脏中miRNA的基因调控表达与血压有关
为了系统地分析肾脏 miRNA 是否与 BP 相关,他们首先在本研究的“发现资源” (HKTR) 平台创建了一个包含 339 个 miRNA 表达谱的存储库,其中包含匹配的基因型信息。他们发现了 1459 个肾脏 miRNA,其中大部分由内含子序列编码(图 5A)。正如预期的那样,与蛋白质编码基因和长非编码RNA相比,miRNA在肾脏基因中所占的比例要小得多(图 5B)。与其他生物型相比,较少的肾脏 miRNA 占肾脏中累积基因表达的相同比例(图 5C)。然后,他们将肾脏miRNA目录与miRNATissueAtlas2管理的35种人体组织的表达谱进行整合,以检测miRNA对肾脏组织的特异性程度。在肾脏目录和miRNATissueAtlas2之间重叠的1431个miRNA中,有49个在肾脏表达升高以及有3个在肾脏显著富集(图5D)。MiR-30a-3p、miR-30a-5p和miR-188-5p在肾脏中强富集,其中一些先前有证据表明它们与肾脏疾病有关(图5D)。利用发现资源中的339个肾脏数据,他们生成并交叉验证了201个肾脏miRNA的GReX预测模型(图5E)。其中143个可供验证,其中80个在由150个具有匹配基因型和小RNA-seq衍生表达谱的美国国立卫生研究院(NIH)肾脏(TCGA和CPTAC)组成的独立资源中进行了验证(图5E-F)。miR-196a-3p是miRNA的一个例子,其强大的遗传调控成分使其成为TWAS的绝佳靶标(图5F)。
他们鉴定了11个肾脏miRNA,它们的遗传输入表达在两个队列中与BP相关(图5G)。他们定位了9个独立的BP GWAS位点,代表了肾脏丰度、组织特异性和遗传起源的不同谱(图5H)。
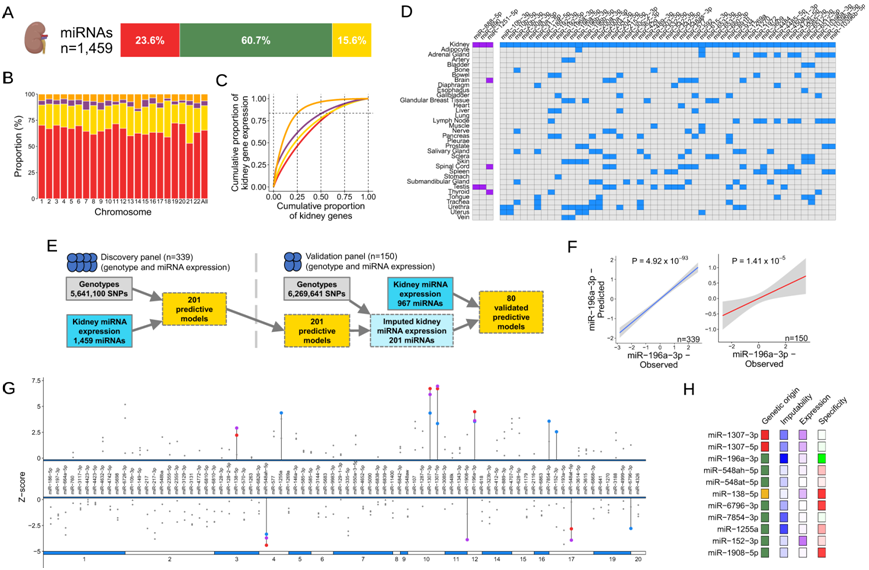
图5. 肾脏 miRNA 和血压。
(a) 人类肾脏中表达的 miRNA 数量及其遗传起源的分布。 (b) 肾脏中表达的基因生物型的累积比例,按染色体分层。(c) miRNA 表达量和基因表达量之和占 miRNA 和基因总数的累积比例,按表达量降序排列。(d) 肾脏中表达富集/增强水平最高的 52 种 miRNA 的组织富集概况。(e) TWAS 中使用的预测 miRNA 表达模型流程。(f) 预测的 GReX 与观察到的 miR-196-3p 表达之间的相关性。(g) 80 个可推算的、经过验证的肾脏 miRNA 与血压特征之间关联的概述。(h) 选定的 miRNA 特征与至少一种血压特征显著相关。
05
肾脏蛋白质组关联研究发现了与血压相关的新蛋白质
使用 72 个 NIH-CPTAC 人类肾脏以及通过液相色谱质谱法表征的组织蛋白质组,他们鉴定了 7291 个蛋白质;其中大多数归类为预测的细胞内蛋白(89%)。可溶性蛋白、膜结合蛋白和分泌蛋白的比例与全球人类蛋白质组的基于组织的图谱数据一致(图6A)。正如预期的那样,他们发现对肾脏具有最高特异性的基因编码的蛋白富集程度最强(图6B)。
然后,他们研究了与血压调节和高血压相关的蛋白质的丰度如何与其各自基因的表达相关。在NIH-CPTAC数据集中,对总共7036个基因在蛋白质水平上的丰度进行定量。已知的孟德尔高血压/低血压综合征基因和降压药物的肾脏靶点基因与各自的蛋白质具有最高的正相关性,且富集程度与HPA肾脏富集/增强基因相当(图6C)。例如,SLC12A1和SLC12A3表现出非常强的mRNA -蛋白相关性(图6D)。在肾脏表达可归因于遗传的815种蛋白,在两个独立队列(英国生物样本库和ICBP)中,有97种蛋白与至少一种血压性状呈相互关联(图6E)。对于46个BP相关的肾脏蛋白,他们确定了在其他分子水平与BP/高血压相关的信息。事实上,在肾脏BP TWAS中,40个蛋白的亲本基因与BP相关(图6F)。
总体而言,BP PWAS蛋白发生显著富集(图6G)。事实上,与一组随机的肾脏蛋白相比,他们的97个BP PWAS蛋白显示出通过小分子和靶向嵌合体(PROTAC)模式具有发现潜力的蛋白的强富集(图6G)。通路富集分析进一步揭示了高血压风险的BP相关蛋白的富集,以及RAS和参与固有免疫和适应性免疫的蛋白的富集(图6H)。
总的来说,他们发现与其亲本 mRNA 呈正相关的肾脏蛋白富含与血压、遗传介导的高血压/低血压和抗高血压治疗相关的基因。
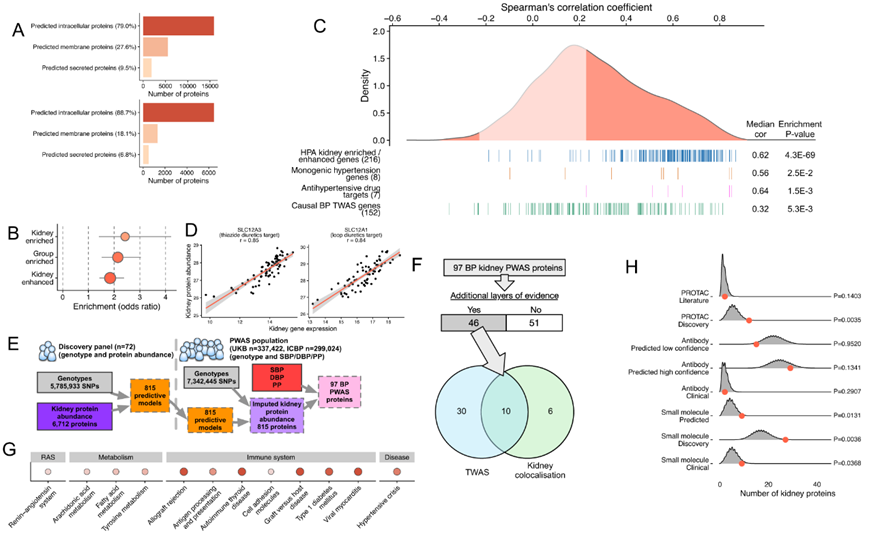
图6. 肾脏组织蛋白质组学和血压 PWAS。
(a) 通过预测定位对人肾中 20082 个蛋白质[来自人类蛋白质图谱 (HPA)]和 7291 个可测量蛋白质(来自 CPTAC)进行分类。 (b) 与所有 HPA 蛋白(20082 种蛋白)相比,可测量的肾脏蛋白(6608 种蛋白)具有组织特异性。(c) 6712 个蛋白质/基因对的蛋白质丰度和基因表达之间的 Spearman 相关系数的密度。 (d) 两个降压治疗靶点(SLC12A3和SLC12A1)的肾组织基因表达与组织蛋白丰度之间的相关性。(e) 人体肾组织 PWAS 工作流程概述。(f) 维恩图的每个区域显示了基因的数量。 (g) KEGG 通路和人类疾病关联分析。(h) 97 BP PWAS蛋白在三种治疗方式下的药物可追踪性富集。
06
对从尿液中收集的细胞进行转录组分析
他们检测到泌尿细胞中 21981 个基因的表达,与人肾组织相比,尿液中表达的主要基因生物型的数量相似(图 7A)。泌尿细胞和肾组织之间的转录组复杂性大致相似(图 7B)。由于尿液供体来源不同(即肾切除术和肾活检)(图7C),没有出现明显的转录组差异。
他们对尿液和肾脏组织中主要高表达的100个基因所做的分析表明,分别有33条和35条通路过度表达,其中24条通路在尿液细胞和肾脏中共同存在(图7D),并且有44条不同的通路过度表达(图7D)。在尿液细胞中,前100个高度表达的基因在反映肾对离子交换贡献的通路上也表现出富集(图7D)。他们还发现,尿液细胞中的基因富集于葡萄糖代谢,糖酵解和糖异生(图7D)。后者与这些细胞的肾脏起源相一致,因为除了肝脏外,肾脏(主要是近端小管)是唯一能够重新产生葡萄糖的器官。
然后,他们研究了来自数据集的尿液细胞和来自GTEx的54种人体组织类型之间的转录组相似程度。19273个蛋白编码基因在肾皮质和肾髓质与泌尿系细胞的表达相关程度最高(图7E-F)。他们用一组独立的430个肾组织样本证实了尿细胞和肾脏之间的相关性(图7F)。
他们还发现肾皮质标记基因(20个基因)和髓质标记基因(11个基因)的表达量与尿细胞的表达量相关(图7G)。尿细胞表达髓质标记物之间有特别强的相关性(图7G)。尿细胞中的UMOD与髓质相当,高于皮质,这与Henle上升环内UMOD的产生一致(图7G)。对于皮质标志物,NAT8, MIOX, RIDA, BHMT在尿液中的相对表达水平与皮质样本相当。
最后,他们检测了 399 个肾脏基因表达的尿细胞-肾脏相关性,在他们的分析中显示与 BP 存在因果关系。泌尿细胞和肾脏之间这些基因的表达存在很强的正相关性(图7H)。
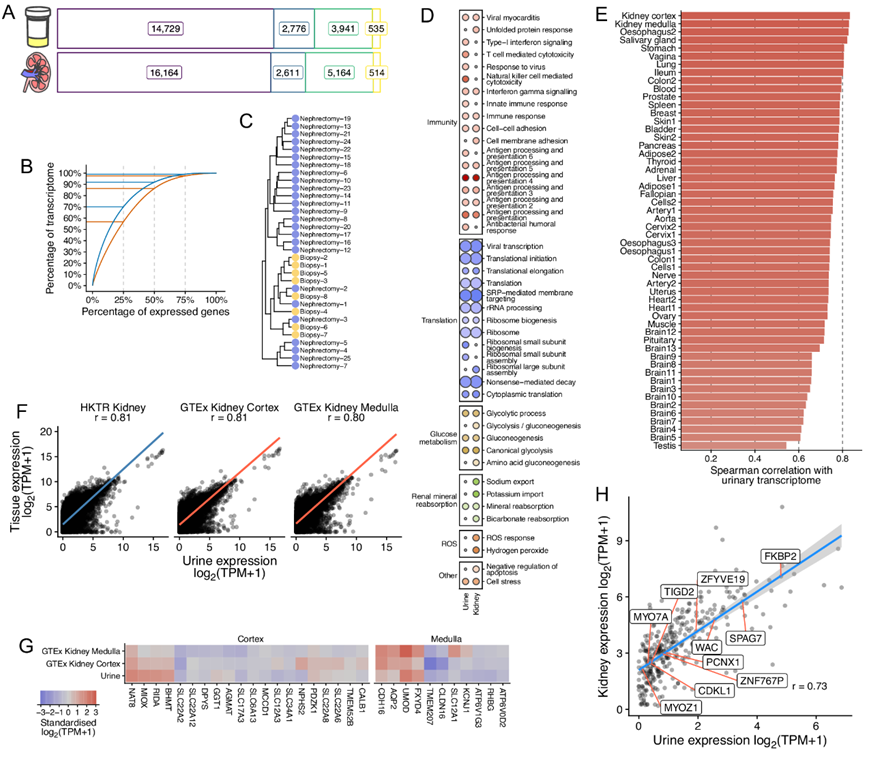
图7.对泌尿细胞进行转录组分析。
(a) 按主要基因生物型划分的尿细胞沉淀和肾脏的表达基因数量。 (b) 泌尿细胞和肾脏的转录组复杂性。(c) 来自肾切除术和活检样本的 33 个尿细胞转录组的层次聚类。(d) 来自肾切除术和活检样本的 33 个尿细胞转录组的层次聚类。 (e) 尿转录组和 54 GTEx 组织之间的转录组相似性。(f) 泌尿细胞和肾脏之间 19273 个蛋白质编码基因表达的相关性。 (g) 肾皮质和髓质特异基因的表达谱。(h) 339 个因果 BP TWAS 基因的尿细胞和肾脏中值表达之间的相关性。
07
基因组水平、肾脏转录组和蛋白质组数据整合
ENPEP编码谷氨酰氨基肽酶——一种在血管紧张素II转化为血管紧张素III的过程中负责从N端切割天冬氨酸的酶(图8A)。ENPEP是在肾脏显著富集的基因之一,它在肾脏中的表达量比大脑皮层中的表达量高约 36 倍(图8B)。
本项研究的肾脏 BP TWAS 研究表明,在两个独立数据集中,基因决定的肾ENPEP表达增加与所有 BP 特征下降之间始终存在很强的关联,并且在汇集了大约 750000 名个体的联合分析和进一步的 MR证明了因果效应(图 8C-D)。在UK Biobank的 337350 名个体中,基因确定的ENPEP肾脏表达与尿钠增加有关(图 8E);这可能表明肾脏 Ang III 介导的尿钠排泄作用是相关机制。通过BP 肾 PWAS,他们还证明了谷氨酰氨肽酶蛋白与 BP 之间的关联方向一致,蛋白质丰度的降低与 DBP 值较高相关(图 8F)。rs33966350的罕见等位基因变异体(A)导致提前终止密码子,其复合注释依赖缺失(CADD)评分为43,是功能变异体的高置信度丧失之一(图8G)。
在FinnGen联盟377,277名个体的2269个二元性状的全表型关联中(图8H),他们确定高血压是rs33966350的最高关联信号。他们在UK Biobank的研究证实rs33966350也与尿钠有关(图8I)。然后,他们发现,在对478个肾脏的分析中,与野生型基因型(GG)相比,具有一个rs33966350 A等位基因的携带者ENPEP mRNA的表达水平降低了约4.9倍(图8J)。他们在对CPTAC的65个肾脏进行的分析中重复了这一观察结果——罕见纯合子基因型携带者的ENPEP水平显著低于GG基因型携带者(图8K)。他们在蛋白水平进一步验证了这些观察结果(图8L)。
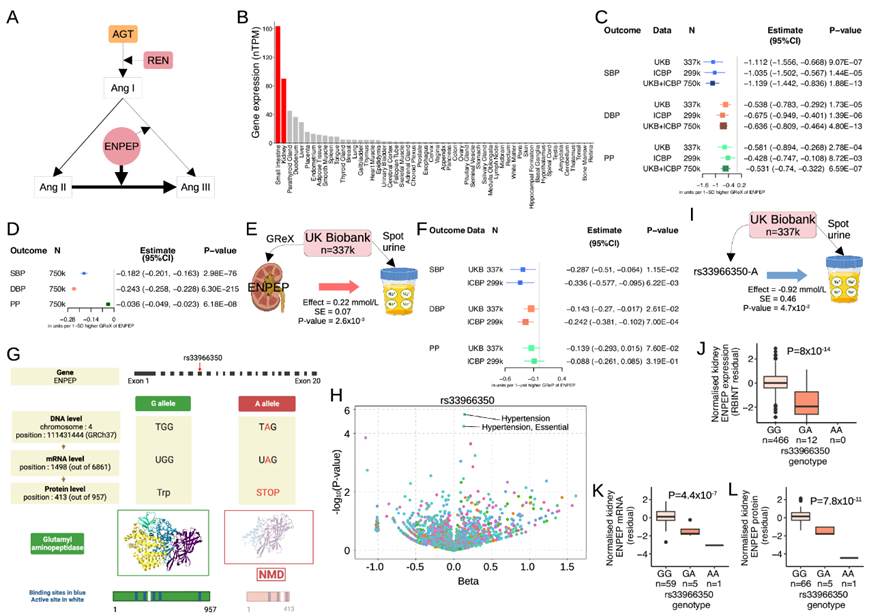
图8. 肾脏谷氨酰氨肽酶基因 (ENPEP) 和血压。
(a) 简化的肾素-血管紧张素系统。(b) GTEx 中标准化的ENPEP表达。(c) ENPEP的基因调控表达 (GReX)对肾脏 TWAS 的收缩压 (SBP)、舒张压 (DBP) 和脉压 (PP) 的影响。(d) 肾脏ENPEP的 GReX对 UKB 和 ICBP 中 SBP/DBP/PP的因果影响。(e) ENPEP的 GReX对 UKB 中尿钠的影响。(f) 谷氨酰氨肽酶基因调节蛋白 (GReP) 对肾脏 PWAS 的 SBP/DBP/PP 的影响。(g) ENPEP [A -次要等位基因,G -主要等位基因]的rs33966350基因导致第6外显子413位(共957个)过早终止密码子。(h) rs33966350-A在FinnGen中的作用。 (i) rs33966350-A 对 UKB 尿钠的影响。(J) rs33966350 对 HKTR 中ENPEP表达的影响。(k-L) rs33966350 对ENPEP的影响CPTAC 中的表达和蛋白质丰度。
+ + + + + + + + + + +
结 论
本项研究利用肾脏特异性表观基因组注释和 3D 基因组信息,生成并验证了基因表达预测模型,以便在 700 个人类肾脏中进行全转录组关联研究,鉴定了 889 个与血压相关的肾脏基因,其中 399 个优先考虑为血压调节的贡献者。肾脏蛋白质组和 microRNA组的估算发现了 97 种肾脏蛋白质和 11 种 miRNA 与 BP 相关。与血浆蛋白质组学和代谢组学的整合阐明了肌醇、4-胍基丁酸和血管紧张素原的循环水平作为多个肾脏 BP 基因(分别为SLC5A11、AGMAT、AGT)的下游效应子。基因决定的肾脏表达减少可能模仿罕见的功能丧失变异对肾脏 mRNA/蛋白的影响,并导致血压升高(例如ENPEP)。本项研究证明了从尿液和肾脏收集的细胞之间蛋白质编码基因的表达存在很强的相关性,突出了尿细胞转录组学的诊断潜力。
+ + + + +



