

 English
English文献解读|Nat Commun(14.7):宫颈癌的蛋白质组学分析揭示了治疗和生物学见解
✦ +
+
论文ID
原名:A proteogenomic analysis of cervical cancer reveals therapeutic and biological insights
译名:宫颈癌的蛋白质组学分析揭示了治疗和生物学见解
期刊:Nature Communications
影响因子:14.7
发表时间:2024.11.22
DOI号:10.1038/s41467-024-53830-0
背 景
宫颈癌 (CC) 仍然是女性第四大常见恶性肿瘤和第四大癌症死亡原因。与许多其他与病毒感染没有直接病因关联的癌症不同,持续性人乳头瘤病毒 (HPV) 感染是 CC 的主要原因,超过 90% 的 CC 患者在诊断时与 HPV 感染有关。尽管 HPV 疫苗接种和筛查策略降低了高收入国家的 CC 发病率,但 CC 仍然是威胁低收入国家妇女健康的主要公共卫生问题。
实验设计
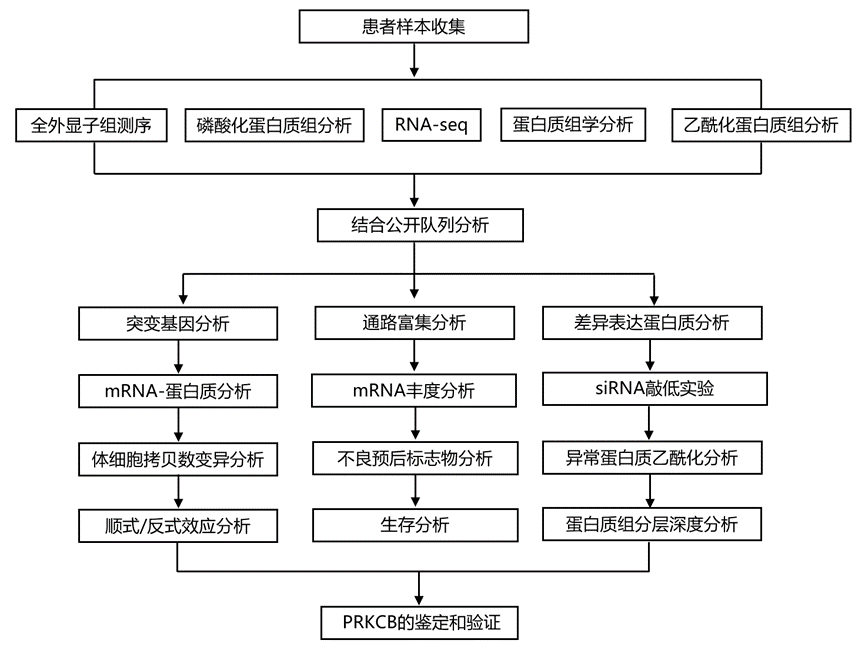
结 果
01

中国 CC 人群的蛋白质组学概况
为了全面了解中国CC的分子特征并提供有价值的生物学见解来指导临床治疗,研究团队使用全外显子组测序(WES)技术、转录组分析(RNA-seq)技术、蛋白质组学技术、磷酸化蛋白质组学技术和乙酰化蛋白质组学技术进行了全面分析。该分析是基于对从北京协和医学院(PUMC)前瞻性收集的总共139个未治疗过的肿瘤样本和33个正常邻近组织(NAT)。在139名患者中,112名诊断为鳞状细胞癌,19名患有腺癌,2名患有腺鳞癌,4名患有宫颈小细胞神经内分泌癌(NECC),2名患有其他混合组织学类型。
在 134 名 CC 患者原发性肿瘤的 WES 数据中共检测到 31047 个非沉默点突变和 1050 个插入-缺失突变。PUMC 队列中最常见的突变基因包括PIK3CA(32.8%)、KMT2C(18.7%)、KMT2D(17.2%)、FAT1(13.4%)、AR1D1A(11.2%)、FBXW7(11.2%)、EP300(8.2%)、TP53(8.2%)、PTEN(7.5%)和ERBB3 (7.5%)(图 1a)。PIK3CA是 CC中最常见的突变基因。KMT2C和KMT2D在本队列中频繁突变,并且在各种类型的癌症中也观察到。在本研究的队列中,宫颈腺癌中KRAS的突变率高于鳞状细胞癌,与 TCGA CC 研究(图 1b)。在本研究中,通过 RNA-seq 定量了 24595 个基因;基于同位素串联质谱标签(TMT)的蛋白质组学分析鉴定了 9600 种蛋白质;磷酸化蛋白质组学分析鉴定了 41448 个高度可靠的磷酸位点,对应 7721 个磷蛋白,乙酰蛋白质组学检测到 5749 个高度可靠的乙酰位点,对应着 2456 个乙酰蛋白。此外,基于数据独立采集(DIA)的蛋白质组学分析鉴定出11904种蛋白质,与TMT蛋白质组学数据具有良好的一致性。
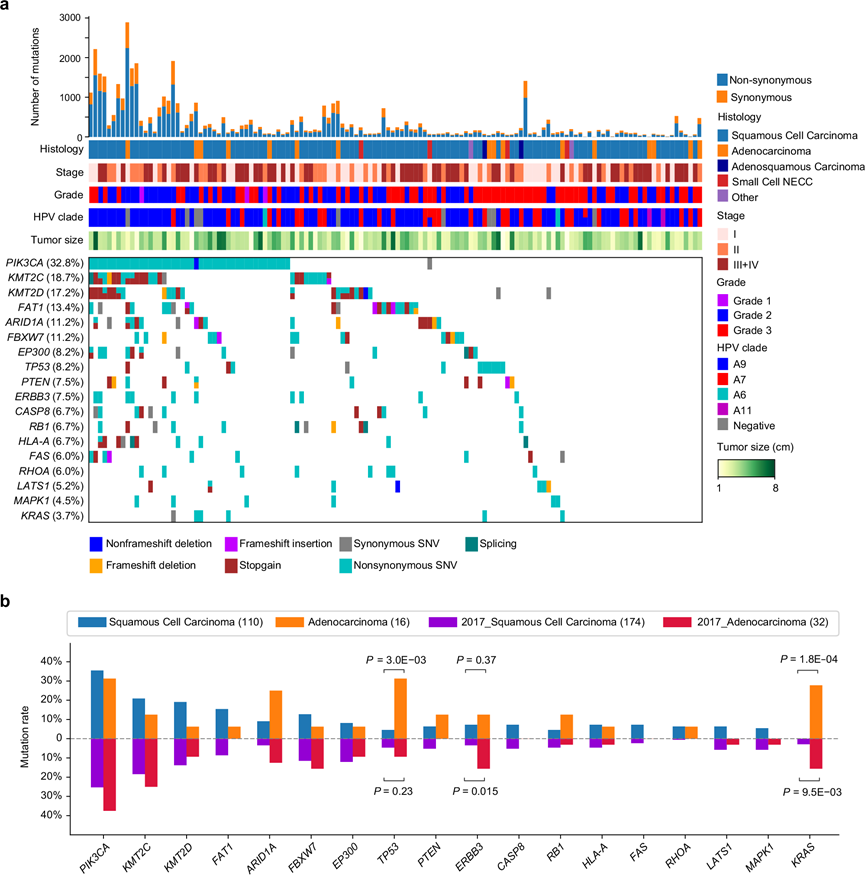
图1. 基于WES的中国CC人群突变谱。
(a) 134 名 CC 患者的基因图谱。(b) 中国和 TCGA CC 队列中宫颈鳞状细胞癌和腺癌的常见突变基因比较。
02
体细胞拷贝数变异的多组学分析
对于 134 个 CC 样本,基于 WES 数据得出体细胞拷贝数变异 (SCNA)。总体而言,全基因组局部变异在染色体 1q、3q、5p 中出现频率最高的增加,在染色体 2q、3p、11q、13q 中出现频率最高的丢失。基因拷贝数的增加和减少通常以顺式或反式作用调节 mRNA、蛋白质和磷蛋白的丰度(图 2a)。拷贝数变异(CNA)-蛋白质和 CNA-磷蛋白之间的顺式和反式关联比 CNA-RNA 的关联更弱(图 2a)。此外,基于 TMT 和 DIA 蛋白质组学数据集,鉴定出 1358 种常见的减毒蛋白,主要在 mRNA 加工、mRNA 剪接和核糖体生物合成等生物过程中富集。因此,翻译后修饰(PTM)可能在决定蛋白质半衰期方面起关键作用,导致基因剂量和蛋白质丰度之间的相关性降低。
为了识别中心 CNA 基因,他们筛选了肿瘤间相关的 CNA、mRNA 和蛋白质水平以及肿瘤和 NAT 之间一致的蛋白质水平变化。178 个基因鉴定为具有顺式效应的拷贝数增加基因(CNG- Cis基因),并在蛋白酶体、RNA 转运、核黄素代谢、剪接体、DNA 复制和 N-糖生物合成中富集(图 2b),突出了 CNA 对肿瘤增殖和进展的功能性影响。值得注意的是,具有淋巴结阳性的肿瘤表现出比具有阴性淋巴结的肿瘤更高的平均 CNG- Cis蛋白丰度(图2c)。同时,鉴定了 28 个具有顺式效应的拷贝数丢失基因(CNL- Cis基因),这些基因参与细胞粘附分子和缬氨酸、亮氨酸和异亮氨酸降解通路(图 2d)。CNL- Cis基因的平均蛋白质表达水平与肿瘤大小呈负相关(图 2e)。此外,具有阳性淋巴结的肿瘤显示出比具有阴性淋巴结的肿瘤明显更低的 CNL -Cis蛋白丰度(图2f)。在CNG -Cis基因中,50 个(28.1%)位于染色体 3q 上,而 16 个(57.1%)CNL- Cis基因位于染色体 11q 上。同时,他们发现 Chr3q 和 Chr11q25 CNA 在蛋白质和磷蛋白水平上具有更多的反式效应(图 2a)。通路富集分析显示,Chr3q 的增加和 Chr11q25 的缺失在蛋白质和磷蛋白水平上对上调细胞周期通路和下调粘着斑通路 表现出相同的反式效应(图2g-j)。此外,还观察到 Chr3q 上的 CNG 和 Chr11q25 上的 CNL 同时出现。值得注意的是,Chr11q25 的缺失与肿瘤尺寸增大和阳性淋巴结转移相关,也可能导致 CC 细胞的增殖和转移。
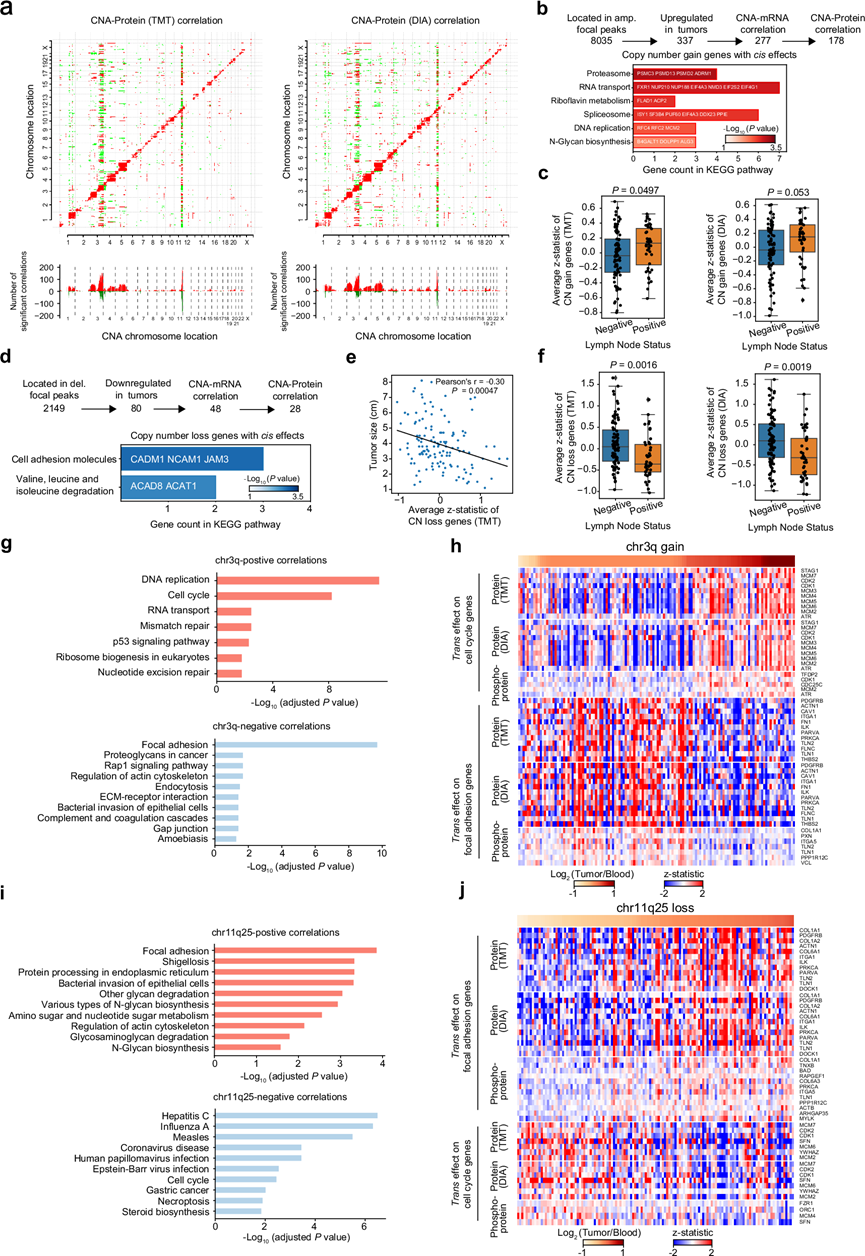
图2. SCNA 的效果。
(a) CNA(x 轴)与 TMT(左)和 DIA(右)蛋白表达(y 轴)的相关性突出显示了 CNA顺式和反式效应。 (b) 识别 CNG- Cis基因的方法示意图。(c) 淋巴结阳性患者的 CNG-Cis基因蛋白表达水平高于淋巴结阴性患者。(d) CNL-Cis基因识别流程图。(e) CNL- Cis基因的蛋白质丰度(TMT)与肿瘤大小呈负相关。(f) 淋巴结阳性患者显示出比淋巴结阴性患者更低的 CNL- Cis基因蛋白质表达水平。(g) KEGG 通路富集分析。(h) 染色体3q 的 CNG 热图以及细胞周期和粘着斑相关蛋白的蛋白质/磷酸化蛋白质丰度。(i) KEGG 通路富集分析。(j) 染色体 11q25 的CNL热图以及细胞周期和粘着斑相关蛋白的蛋白质/磷酸化蛋白质丰度。
03
HPV 相关蛋白质组学变化概况
HPV检测及分型基于RNA-seq数据,鳞状细胞癌样本中HPV阳性率最高(98.2%),其中77.7%感染HPV A9亚型;腺癌样本中A9和A7亚型感染率分别为16.7%和33.3%,腺癌样本中HPV阴性率为44.4%(图 3a)。且所有肿瘤中HPV阳性样本比例为92.6%,明显高于NAT中的42.9%(图 3a)。肿瘤中的病毒基因 mRNA 丰度(E1/2、E5/6/7、L1/2)显著高于 NAT,表明严重的 HPV 感染主要局限于宫颈病变内。此外,感染 HPV A9 的肿瘤样本的免疫浸润评分高于感染 HPV A7 的肿瘤样本(图 3b)。
与之前研究的E6和E7能够通过形成复合物促进宿主细胞的恶性转化一致,在肿瘤样本中E6和E7的mRNA丰度之间存在很强的正相关性。为了进一步阐明病毒基因和蛋白质组与肿瘤表型之间的关系,他们对E6进行了相关性分析,发现与E6表达正相关的蛋白质在细胞周期途径中显著富集,而与E6表达负相关的蛋白质与粘着斑、内质网中的蛋白质加工和N-糖生物合成有关。同时,在E6 mRNA丰度较高的肿瘤中H2AX和WEE1的磷蛋白上调,而它们的蛋白质表达没有明显改变(图 3c-e)。先前的研究表明,HPV 阳性肿瘤对 WEE1 抑制剂 Adavosertib 的治疗有显著反应。H2AX的磷酸化是 DNA 双链断裂的标志38。肿瘤样本中的两个关键 HPV 衣壳基因 L1 和 L2 表现出高度相关性。L1 和 L2 共同构成 HPV 的衣壳,这对于 HPV 在环境中的生存、传播和扩散至关重要。与 L1 mRNA 丰度正相关的蛋白质在 DNA 复制中富集,而负相关的蛋白质参与细胞粘附、血小板活化、Fc γ 受体介导的自噬和白细胞跨内皮迁移。有趣的是,L1 基因的转录水平与免疫浸润评分和基质评分呈负相关。
感染HPV16和HPV18的病变的致癌进展通常与病毒基因组整合到宿主细胞染色体有关,而这一过程常常导致 E2 基因表达的缺失。此外,他们观察到 E2 和 E5 的 mRNA 丰度在该队列中呈现双峰分布,因此他们将肿瘤聚类为 E2/E5 mRNA 丰度高或低的两组。VRK2 和 MTDH 在 HPV_G2 肿瘤中表现出较高的表达(图 3f)。VRK2 已确定为肝癌、肾癌和胰腺癌的不良预后标志物,而 MTDH 是肺癌、肾癌和胰腺癌的不良预后标志物。此外,与 HPV_G1 相比,HPV_G2 中 SMIM13 和 VANGL1 的磷酸化增加(图 3f)。VANGL1 在调节癌症发展中的极化细胞行为方面起着重要作用。HPV_G2 中 HPV A7 感染的患病率更高(图 3f)。此外,HPV_G2 中的患者明显比 HPV_G1 中的患者更年轻(图 3g),无进展生存期 (PFS)更差(图 3h)。总之,这些数据揭示了 HPV 相关蛋白质组学变化的概况以及 HPV 对预后的影响。

图3. 基于 HPV E2/E5 mRNA 水平的 HPV 相关蛋白质组学景观和患者亚组。
(a) 中国 CC 队列中 HPV 感染类型分布。(b) 箱线图显示不同类型的 HPV 进化枝之间的免疫评分比较。(c) 散点图显示肿瘤样本中蛋白质(x 轴)/磷蛋白(y 轴)与 HPV E6 mRNA 丰度之间的 Spearman 相关性。(d) 散点图显示 HPV E6 mRNA 与 H2AX 蛋白/磷蛋白丰度之间的 Spearman 相关性。(e) 散点图显示 HPV E6 mRNA 与WEE1蛋白/磷蛋白丰度之间的 Spearman 相关性。(f) 根据 HPV E2/E5 mRNA 水平对患者进行亚组分析。(g) 两个 HPV 亚组患者之间的年龄差异。(h) 基于HPV 亚组的根治性放射治疗 CC 患者的 PFS 的 Kaplan-Meier 曲线。
04
与 CC 肿瘤发生相关的蛋白质组学改变
肿瘤和配对的 NAT 显示蛋白质、磷酸位点和乙酰位点存在显著差异。总计 3268 个(38.8%)蛋白质表现出显著差异表达,其中 483 个蛋白质上调,1041 个蛋白质下调,其丰度变化超过 2 倍(图 4a)。上调蛋白质在 DNA 复制、错配修复、p53 信号通路和细胞周期通路中富集,而下调蛋白质则在细胞粘附分子、粘着斑、补体和凝血级联等通路中富集(图 4b)。使用改变的磷酸位点或乙酰位点相关蛋白进行富集分析,显示出相似的结果。此外,他们将 23 种蛋白质定义为 CC 相关蛋白质,这些蛋白质在至少 80% 的所有肿瘤-NAT 对中升高了 2 倍以上,并通过人类蛋白质图谱注释了其潜在的临床效用(图 4c)。此外,使用 DIA 蛋白质组学验证了这些蛋白质在肿瘤中的表达升高。
接下来,他们比较了宫颈腺癌和鳞状细胞癌之间的分子差异,重点关注在多组学水平上与 NAT 相比在肿瘤中丰度增加的分子(图 4d)。对于蛋白质,他们发现免疫相关的生物过程,例如对 I 型干扰素的反应、对干扰素β/γ 的反应以及先天免疫反应的调节在鳞状细胞癌中显著上调,而几种代谢过程在腺癌中富集(图 4d)。干扰素诱导的抗病毒 RNA 结合蛋白(IFIT1/2/3)、干扰素调节因子 6(IRF6)、干扰素诱导的 GTP 结合蛋白(MX1/2)和 STAT1/2 的表达仅在鳞状细胞癌肿瘤中显著上调(图 4e)。此外,免疫检查点和MHC分子(包括CD274、CD44、CXCL10和HLA-E/F/G)在鳞状细胞癌肿瘤中的表达也上调(图 4f)。因此,他们推测鳞状细胞癌具有活跃的免疫微环境,可能从免疫治疗中受益。
为了探索CC肿瘤进展的分子动力学,他们在不同阶段(I,II,III + IV)进行了差异蛋白表达分析。这些差异表达蛋白的KEGG通路富集分析表明,在肿瘤进展过程中,丙酮酸代谢、糖酵解/糖异生、碳代谢和半乳糖代谢等几种代谢途径发生失调(图 4g)。值得注意的是,14种蛋白质在CC进展过程中呈现总体正调控趋势,在肿瘤和NAT之间显著上调,而9种蛋白质呈现相反趋势,在肿瘤中下调(图 4h)。此外,TCGA CC队列用于更好地了解这些代谢相关蛋白的临床结果,并表明高转录水平,包括B4GALT1、DHCR24、AGPS、ACACA、HK2和SLC2A1,与CC中较差的总体生存率(OS)呈正相关(图 4i-l)。值得注意的是,HK2 表达与肿瘤大小、病理分级和预后呈正相关,并通过 AKT 通路在 CC 中起致癌作用。SLC2A1高表达作为 OS 的独立预后因素可能与 CC 的免疫调节有关。然而,B4GALT1、DHCR24、AGPS 和 ACACA 与 CC 的相关性尚未在以前的研究中报道。除了与 CC 分期相关外,他们发现上述四种蛋白的表达在淋巴结阳性的患者中更高。为了进一步验证这些基因在 CC 中的驱动作用,他们进行了 CCK-8 和克隆形成实验。这些结果表明,通过 siRNA 敲低 B4GALT1、DHCR24、AGPS 和 ACACA 显著降低了 SiHa 细胞的细胞增殖和集落形成能力,进一步表明它们在 CC 进展中的致癌作用。

图4. 与 CC 肿瘤发生相关的蛋白质组学改变。
(a) 火山图描绘了配对肿瘤和 NAT 之间差异表达的蛋白质。(b) 通路分析,显示显著上调/下调的蛋白质。(c) 箱线图显示配对肿瘤和 NAT 之间 CC 相关蛋白质。(d) GO分析。(e) 宫颈鳞状细胞癌和腺癌之间显著上调的I型干扰素相关蛋白的反应相对丰度的热图。(f) 免疫检查点和 MHC 分子在蛋白质水平的表达。(g)KEGG通路分析。(h) 不同阶段之间以及肿瘤与 NAT 之间差异表达的代谢相关蛋白质的总结。(i-l) 在 NAT 和不同阶段肿瘤之间的蛋白质表达比较。
05
异常蛋白质乙酰化调节CC进展
迄今为止,很少有人尝试系统地描述和利用 CC 中的乙酰化组调控。在本研究中,他们对赖氨酸乙酰转移酶 (KAT)、含溴结构域蛋白 (BRD) 以及具有蛋白质和乙酰化丰度的组蛋白进行了差异分析。与 NAT 相比,这些蛋白质的乙酰化在肿瘤中相对于其蛋白质丰度显著上调(图 5a)。值得注意的是,他们发现 EP300 在其组蛋白乙酰转移酶结构域 (HAT) 中具有大量的 K1542、K1546、K1549 和 K1590 过度乙酰化(图 5a)。其中,K1549 是活化环中可能激活 EP300 活性的关键自乙酰化位点,在肿瘤中显著上调。
然后,他们通过检测 EP300 本身的乙酰化来分析对 EP300 活性的潜在影响,并与乙酰蛋白质组学数据进行相关性分析。因此,EP300 的高乙酰化水平与 FOSL2 的高乙酰化相关(图 5b),FOSL2 是一种属于 AP-1 家族的转录因子,据报道参与细胞增殖和分化。然而,FOSL2 在 CC 中的乙酰化或功能尚未研究过。有趣的是,他们发现 FOSL2-K222 乙酰化显著上调,但其蛋白质丰度没有上调(图5c)。
为了进一步探究 EP300 与 CC 中的 FOSL2-K222 之间的联系,他们进行了一系列全面的实验。首先,共转染 EP300 而非 KAT2B、KAT5 或 KAT2A 可促进 FOSL2 乙酰化 (图 5d)。质谱 (MS) 分析进一步证实了 EP300 共转染后 FOSL2-K222 乙酰化水平的升高。与 FOSL2-WT (野生型) 相比,当 FOSL2-K222R 发生突变(乙酰化缺陷)时,与 EP300 共转染引起的 FOSL2 乙酰化水平显著降低(图 5e),这表明 K222 是 EP300 对 FOSL2 进行乙酰化的主要位点。FOSL2 已证明是多种癌症的癌蛋白,包括肺癌、乳腺癌和肝细胞癌,而 WNT5A 已证明是 FOSL2 的转录靶基因。为了探索 FOSL2-K222 乙酰化在 CC 中的作用,他们进行了实时定量 RT-PCR (qRT-PCR) 以验证 FOSL2-WNT5A 在 SiHa 细胞中的调控(图 5f)。他们建立了稳定表达 FLAG 标记的 FOSL2-WT、FOSL2-K222R 和 FOSL2-K222Q 的 SiHa 细胞(图 5g)。有趣的是,他们发现乙酰化模拟 FOSL2-K222Q 突变体在提高 WNT5A 转录方面比 FOSL2-WT 更有效(图 5h)。此外,与 FOSL2-WT 和 FOSL2-K222R 相比,表达 FOSL2-K222Q 的细胞表现出更强的增殖率(图 5i),并在异种移植肿瘤发生试验中形成最大的肿瘤(图5j-l),表明 FOSL2-K222 乙酰化具有很强的促肿瘤作用。因此,这些结果共同表明 EP300 至少通过 FOSL2-K222 乙酰化公平地促进了 CC 细胞增殖。
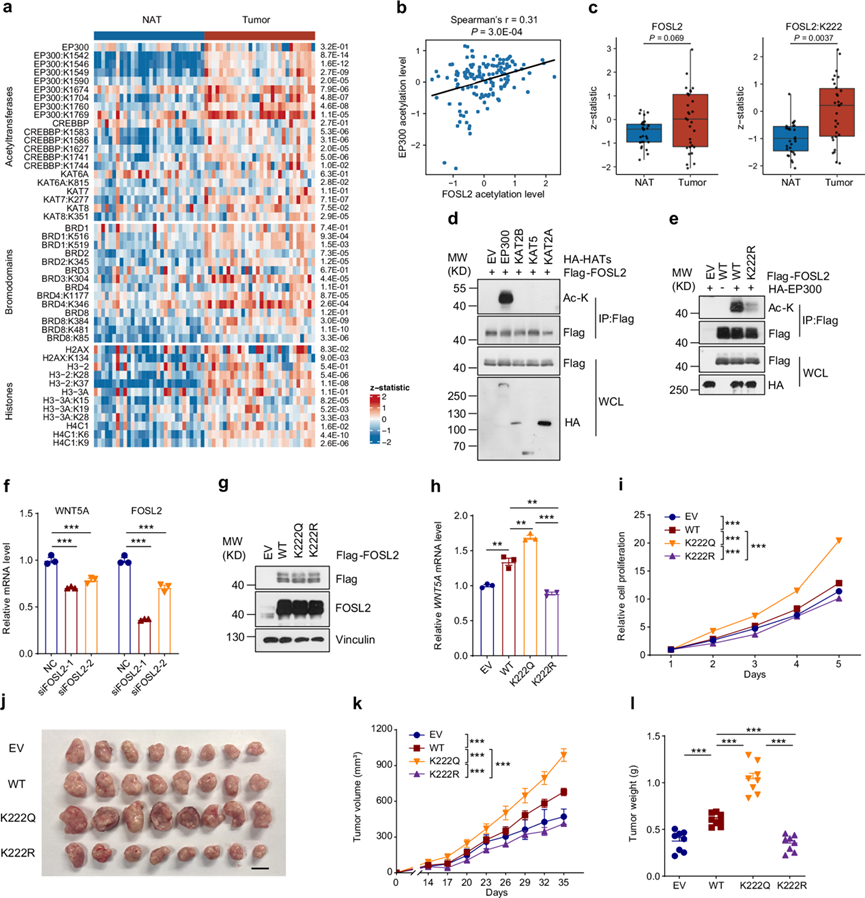
图5. 异常的蛋白质乙酰化调节 CC 进展。
(a) 热图显示配对肿瘤和 NAT 之间 KAT、BRD 和组蛋白的蛋白质和乙酰化丰度。(b) 散点图显示 FOSL2 乙酰化水平(x 轴)和 EP300 乙酰化水平(y 轴)的 Spearman 相关性。(c) 箱线图显示配对(n = 30)肿瘤和 NAT 之间 FOSL2 蛋白质或 K222 乙酰基位点表达的比较。(d) 免疫印迹分析。(e) K222突变显著降低了 EP300 对 FOSL2 的乙酰化。(f) WNT5A 和 FOSL2 的相对 mRNA 水平。(g) 免疫印迹分析。(h) 采用qRT-PCR方法检测WNT5A在SiHa细胞中的相对mRNA水平。(i) 检测指示SiHa细胞的增殖情况。(j) 不同组裸鼠异种移植瘤图片。(k) 裸鼠皮下注射指示SiHa细胞的肿瘤生长曲线。(l) 各组裸鼠肿瘤重量。
06
具有不同生物学和临床特征的蛋白质组亚群
他们根据 TMT 蛋白质组学数据进行了无监督共识聚类,得出三个蛋白质组学亚组(图 6a)。进一步关注具有 PFS 信息的根治性放射治疗患者,观察到三个蛋白质组学亚组之间的 PFS 存在显著差异。亚组 3 中的患者表现出最佳 PFS,即使进一步按已知的预后临床病理变量(例如分期、淋巴结状态、分级和组织学类型)划分时也是如此(图 6b)。基于 DIA 的蛋白质组学和基于 TMT 的磷酸化蛋白质组学/乙酰化蛋白质组学数据也分别用于将 CC 患者聚类为三个亚组,与 TMT 蛋白质组学亚组一致,证明了蛋白质组学亚组划分的可靠性和稳健性。基于转录组的亚组与 TMT 蛋白质组亚组的一致性较低,并且它们的 PFS 差异不如基于蛋白质组学分析的 PFS 差异明显,凸显了蛋白质组亚组分析的优越性。
他们发现在亚组 1 中腺癌和 HPV-A7 感染的患病率较高,而异常 SCC 水平的发生率较低。亚组 2 显示 11q25 缺失和 3q29 扩增的发生率最高(图 6a)。监督差异表达分析揭示了亚组特异性蛋白质。在亚组 1 中,与角质形成细胞分化和 KRT 家族 KRT4、KRT5、KRT6A、KRT6B) 以及 S100A 家族成员 (S100A7、S100A8、S100A9) 相关的蛋白质发生下调,这表明亚组 1 可能与之前报道的角蛋白-Low聚类14有关。在亚组 2 中,395 种蛋白质发生上调并在 DNA 复制中富集(MCM 家族),表明这些肿瘤具有增殖特征。在亚组 3 中,381 个蛋白质上调,并富集在补体和凝血级联以及中性粒细胞胞外陷阱形成中,表明亚组 3 的炎症状态。通路富集分析还显示,亚组 1 中的蛋白质丰度高于亚组 2 和 3,这些蛋白质在 ECM-受体相互作用、肌动蛋白细胞骨架的调节以及氨基糖和核苷酸糖代谢通路中富集;亚组 1 和 2 中上调的蛋白质与核糖体相关,表明这两个亚组中的 CC 肿瘤核糖体生物合成过度激活(图 6a)。此外,亚组3的免疫浸润评分最高,其次是亚组2,然后是亚组1(图 6c)。这可以进一步解释亚组3患者对放射治疗的反应率最高。虽然亚组1患者的肿瘤较小,但其较差的PFS可以归因于其明显较低的免疫浸润评分(图 6d)。进一步比较细胞异质性的情况发现,亚组1患者的基质细胞比例最高,NK、γδT和DC细胞较少,亚组3患者的单核细胞和中性粒细胞比例明显较高(图 6e),这表明亚组3患者可能从免疫治疗中受益。
为了研究CC亚群的磷酸化蛋白质组学特征,他们对差异表达的磷酸位点进行无监督聚类,揭示了与四个蛋白质模块相对应的四个磷酸化模块(图 6f)。应用激酶-底物富集分析(KSEA)进一步鉴定亚群特异性激酶(图 6g)。PRKACA,PRKACB,PRKAA1和PRKAA2鉴定为亚群1特异性激酶;CDK1,CDK2,EIF2AK2,MAPKAPK5和TTK鉴定为亚群2特异性激酶,与不良预后相关。PRKCB和PRKCD鉴定为亚群3特异性激酶。PRKCB参与B细胞活化和诱导凋亡,是结肠癌的肿瘤抑制因子。SiHa 细胞中 PRKCD 的过表达增强了放射敏感性,而通过 siRNA 沉默 ME180 细胞中的 PRKCD 表达则降低了放射敏感性。在 TMT 和 DIA 蛋白质组学数据集中,这些亚类特异性激酶的蛋白质丰度显示出与磷酸化相似的表达模式,进一步证明了磷酸化功能与蛋白质表达之间的紧密关联(图 6h)。
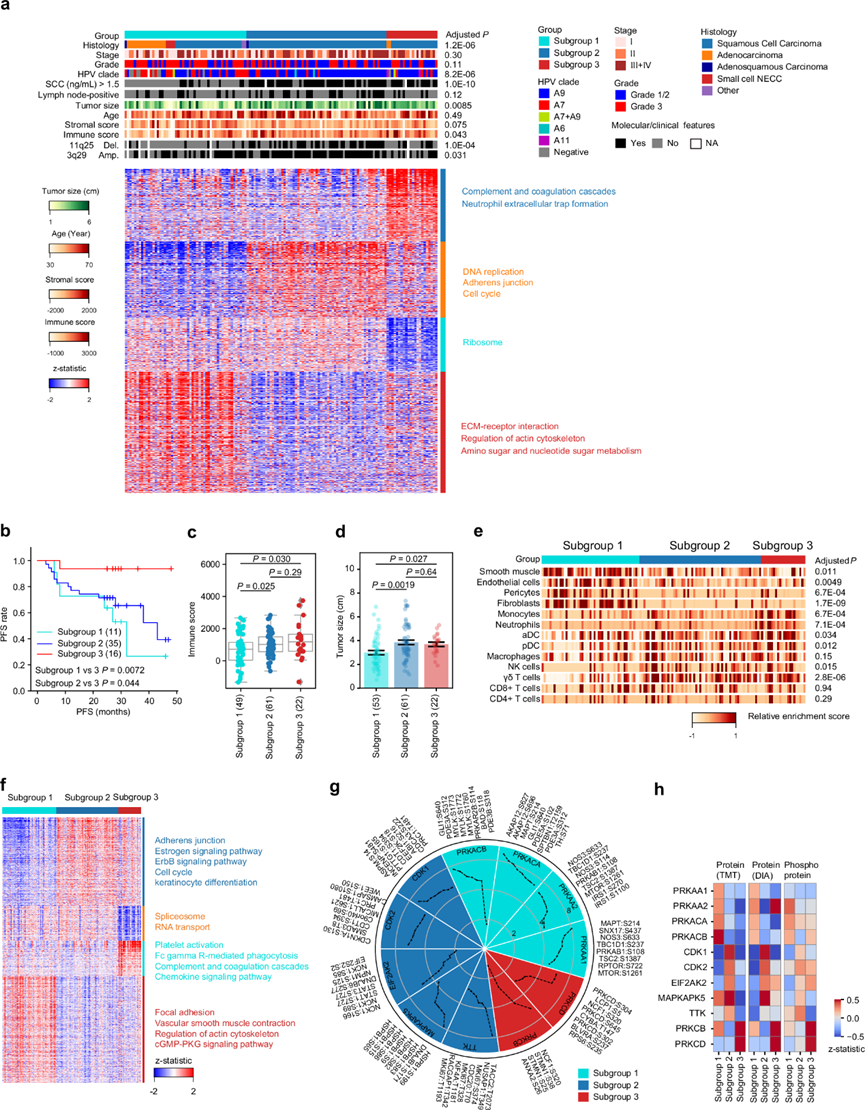
图6. CC 队列的蛋白质组分层和相应的蛋白质通路和亚组特异性激酶。
(a) 蛋白质组学分析的患者亚组分析确定了三个蛋白质组亚组。上:标注了蛋白质组亚组与临床特征和体细胞突变的关联。下:差异表达蛋白分析。(b) 各 TMT 蛋白质组亚组中根治性放疗患者的 PFS 的Kaplan -Meier 曲线。(c-d) TMT 蛋白质组亚组间免疫评分和肿瘤大小的比较。(e) 反卷积分析。(f) 三个 TMT 蛋白质组亚组之间的差异磷酸化位点。(g) 圆形图描绘了每个 TMT 蛋白质组亚组中的活性激酶与 KSEA 鉴定的所有其他亚组相比的情况。(h) 热图显示 TMT 亚组特异性激酶的蛋白质(TMT 和 DIA)和磷酸化丰度。
07
放射反应相关生物标志物和风险评分模型的鉴定和验证
通过将 PFS 数据与根治性放疗患者的蛋白质表达相结合,他们确定了 92 种与放射反应相关的生物标志物,包括 37 种有利和 55 种不利候选物。值得注意的是,已确定为有利的生物标志物之一的亚组3特异性激酶PRKCB在肿瘤中下调(图7a)。免疫组织化学(IHC)图像也显示与NAT相比,肿瘤中的PRKCB表达降低。此外,PRKCB蛋白水平与临床特征没有显著相关性,包括不同阶段、淋巴结转移和分级状态。高PRKCB的肿瘤显示Fc Gamma R介导的吞噬作用、自然杀伤细胞介导的细胞毒作用、T细胞受体信号通路和B细胞受体信号通路的特异性上调,以及细胞周期、DNA复制、剪接体和ErbB信号通路的下调(图 7b)。此外,PRKCB 蛋白(TMT/DIA)表达较高的根治性放疗患者 PFS 较好(图 7c-d)。此外,在 TCGA CC 队列中,PRKCB mRNA 水平升高与 OS 较好呈正相关(图 7e)。来自接受根治性放疗的 124 名患者的独立队列的 IHC 结果进一步验证了 PRKCB 表达较高与放疗疗效较好呈正相关(图 7f)。
为了探索 PRKCB 在 CC 细胞生长中的作用,他们进行了 CCK-8 实验,并证实了在 SiHa 细胞中 PRKCB 的过度表达会抑制肿瘤细胞增殖(图 7g-h)。在异种移植模型中,PRKCB 过表达细胞形成的肿瘤比对照细胞小(图7i-k)。值得注意的是,PRKCB 的过度表达显著增强了辐射的杀伤作用并诱导 SiHa 细胞中的G2/M 周期停滞,这意味着 PRKCB 可能通过调节细胞周期进程来增强放射敏感性(图7l-m)。总之,这些数据表明 PRKCB 可能是预测放射治疗效果的有希望的生物标志物,并可能在 CC 中发挥肿瘤抑制作用。

图7. 放射反应相关生物标志物 PRKCB 的鉴定和验证。
(a) NAT 和 TMT 蛋白质组亚群中 PRKCB 的相对蛋白质(TMT 和 DIA)和磷蛋白丰度。(b) PRKCB 蛋白丰度与多组学谱的关联。(c-f) Kaplan-Meier 曲线。(g) 免疫印迹分析。(h) SiHa 细胞的增殖。(i) 不同组裸鼠异种移植肿瘤大小。(j) 皮下注射到裸鼠体内的指示 SiHa 细胞的肿瘤生长曲线。(k) 不同组裸鼠的肿瘤重量。(l) SiHa 细胞中PRKCB过表达的放射增敏作用。(m)流式细胞分析。
+ + + + + + + + + + +
结 论
本项研究对从 139 名中国女性中获得的 CC 肿瘤进行了全面的蛋白质组学分析。综合蛋白质组学分析将遗传畸变与下游发病机制相关途径联系起来,并揭示了 HPV 相关多组学变化的概况。发现 EP300 可增强 FOSL2-K222 的乙酰化,从而加速 CC 细胞的恶性增殖。蛋白质组学分层确定了三个患者亚组,它们在预后、基因改变、免疫浸润和翻译后修饰调控方面具有不同的特征。PRKCB 是 CC 患者潜在的放射反应相关生物标志物。这项研究为研究人员和临床医生提供了宝贵的公共资源,以深入研究 CC 的分子基础、确定潜在的治疗方法并最终推进临床实践。
+ + + + +



