

 English
English文献解读|BMC Genomics(4.547):RNA-Seq揭示miRNA在山羊棕色脂肪组织产热调控中的作用
✦ +
+
论文ID
原名:RNA-Seq reveals miRNA role in thermogenic regulation in brown adipose tissues of goats
译名:RNA-Seq揭示miRNA在山羊棕色脂肪组织产热调控中的作用
期刊:BMC Genomics
影响因子:4.547
发表时间:2022.3.7
DOI号:10.1186/s12864-022-08401-2
背 景
在哺乳动物中,脂肪组织分为棕色脂肪组织(BAT)、白色脂肪组织(WAT)和米色脂肪组织。与WAT相比,BAT可以通过消耗多室脂滴来实现非颤抖性产热,这对哺乳动物新生儿的生存具有重要意义。目前,对BAT的研究大多集中在人类和小鼠上,对农场动物的研究较少。山羊BAT是重要的产热器官,分布于肾周,在出生后1个月左右消失,其在维持正常体温及降低死亡率方面起着至关重要的作用。因此,探究新生羔羊在适应寒冷的宫外环境过程中,BAT的产热调节机制是十分必要的。本研究通过RNA测序筛选了WAT和BAT之间差异表达的miRNA,以期更好地了解在山羊BAT发育中具有调控功能的miRNA,为理解miRNA在调控山羊BAT产热和能量消耗中的作用提供了证据。
实验设计
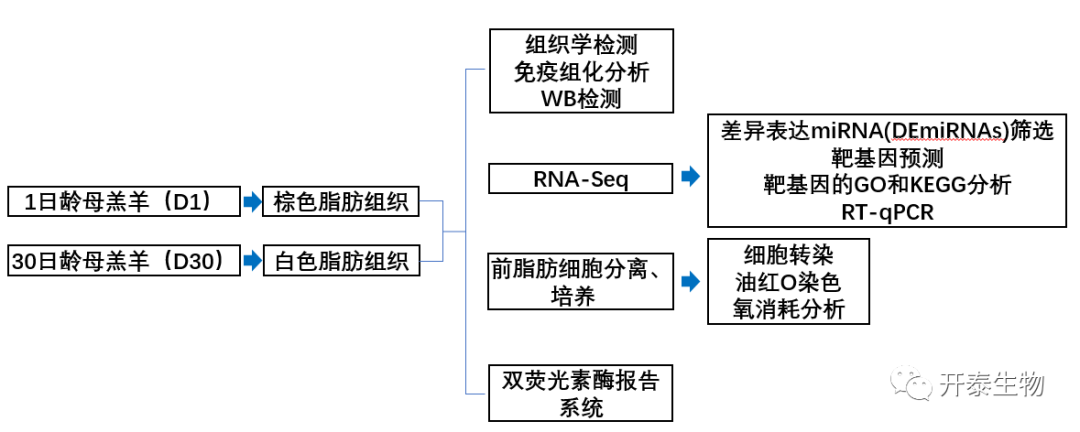
结 果
01

山羊出生后1天(D1)和30天(D30)的肾周脂肪特征
与D30的肾周脂肪相比,D1的肾周脂肪脂滴较小,具有典型的多腔结构,D30的肾周脂肪组织脂滴较大(图1A)。此外,免疫组化观察到D1的肾周脂肪中UCP1蛋白高表达(图1B)。WB检测结果表明,UCP1蛋白在在D1的肾周脂肪组织中高表达,在D30时无法检出(图1C)。UCP1、PGC1-α和DIO2等BAT的标记基因在D1时高表达(图1D)。这表明BAT在新生儿体内存在,并在D30时动态转化为WAT。
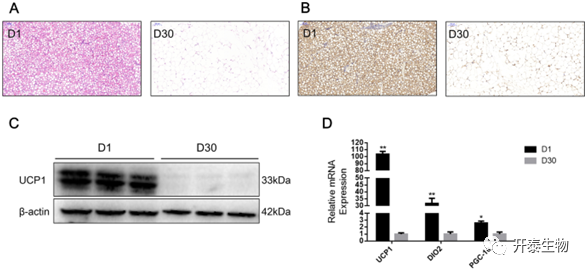
图1 肾周脂肪组织特征
02
小RNA测序分析
不同阶段样本间的高度相关性表明同一时期样本内具有较好的生物学重复(图2A)。在6个文库中总共鉴定了2072个miRNA,其中包括238个已知miRNA和1834个新miRNA。在WAT和BAT中共发现了1651个miRNA。此外,188个miRNA在BAT中特异性表达,233个miRNA在WAT中特异性表达(图2B)。miRNA的长度大多在21 ~ 23 nt之间,且以22 nt为主(图2C、D),符合miRNA典型的长度特征。
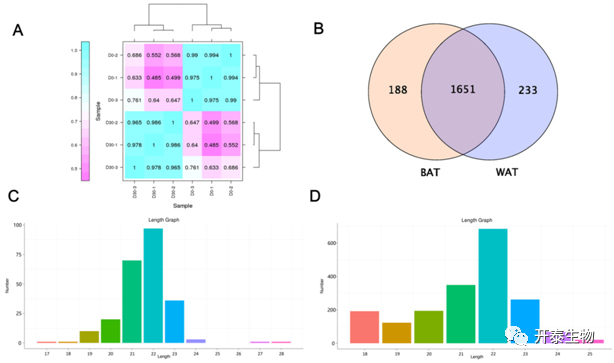
图2 小RNA测序及注释
03
WAT和BAT差异表达miRNAs(DEmiRNAs)的鉴定
在WAT和BAT之间共获得了395个DEmiRNAs,其中167个上调,228个下调(图3A)。聚类分析显示WAT和BAT之间miRNAs表达模式存在差异(图3B)。数据库分析表明,山羊、小鼠和人BAT中分别富集了29、59和9个miRNA。5个miRNA是山羊和小鼠BAT中共有而在人BAT中不存在的,2个miRNA是人和小鼠BAT中共有而在山羊BAT中不存在的。但是,miR-99b-5p在山羊和人BAT中共同表达,但在小鼠中不存在;miR-23b-5p在3个物种中都表达(图3C)。
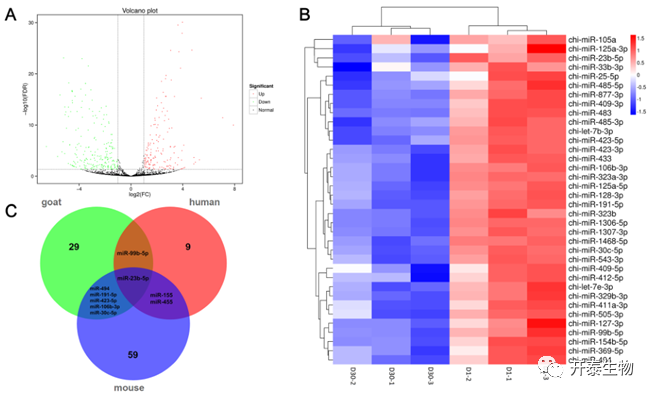
图3 DEmiRNAs分析
04
RT-qPCR验证
为了验证测序数据的准确性,选择3个已知的miRNAs(miR-106b-3p、miR-543-3p和miR-29a-3p)进行RT-qPCR检测,结果和测序数据一致(图4)。
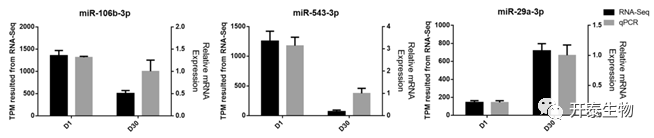
图4 RT-qPCR验证
05
miRNA靶基因GO和KEGG分析
为了解BAT中特异性表达的miRNA的调控功能,作者对BAT富集的miRNA的靶基因进行了GO和KEGG分析。GO分析显示,这些靶基因主要富集在细胞过程、细胞功能和结合功能中(图5A)。KEGG分析显示,这些靶基因主要参与轴突导向、内吞作用、Notch信号通路、Hippo信号通路和MAPK信号通路(图5B)。这表明BAT中特异性表达的miRNA可能通过靶向这些通路中的基因而发挥调节作用。
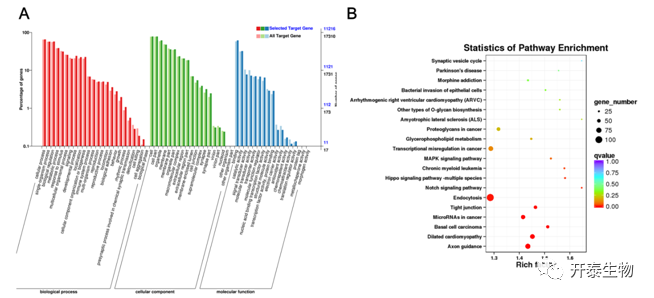
图5 miRNA靶基因的GO和KEGG分析
06
miR‑433阻止棕色脂肪细胞的分化和产热
在BAT富集的miRNA中,miR-433是BAT中上调差异倍数最大的miRNA之一,其在BAT中的功能尚不清楚。miR-433在增殖的棕色前脂肪细胞中高表达,但随着细胞的分化而下降(图6A)。将miR-433模拟物转染到前脂肪细胞中,与NC模拟物相比,miR-433的过表达减少了棕色脂肪细胞的脂质沉积(图6 B)。此外,miR-433显著降低了脂肪分化标志基因FASN和FABP4的表达(图6C)。同时,miR-433显著降低了UCP1和线粒体相关基因COX1和ATP6的表达(图6C)。另一方面,沉默miR-433后,棕色脂肪细胞的脂质沉积增多(图6D),并提高了包括UCP1、PPARg、FASN、FABP4和ATP6在内的几种产热和脂肪形成标志基因的表达水平(图6 E)。
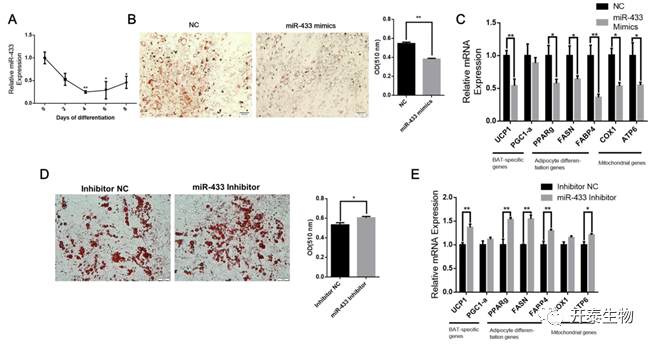
图6 miR-433抑制棕色脂肪细胞早期分化
为了进一步验证miR-433是否可以调节成熟棕色脂肪细胞,作者转染了miR-433模拟物至分化第8天。结果显示miR-433过表达抑制了脂滴的沉积(图7A),降低了棕色脂肪细胞和脂肪形成因子的mRNA水平,同时线粒体相关基因的表达也下降(图7 B)。
作者进一步进行耗氧测定以评估miR-433是否影响产热。加入异丙肾上腺素后,转染miR-433模拟物的棕色脂肪细胞中UCP1依赖性耗氧速率显著降低(图7 C)。沉默miR-433增加了棕色脂肪细胞脂滴的沉积(图7 D),同时上调了UCP1、PGC1-a、PPARg、FASN、FABP4、COX1和ATP6的表达(图7 E)。加入异丙肾上腺素后,miR-433抑制剂处理的棕色脂肪细胞的UCP 1依赖性耗氧量也显著高于NC抑制剂处理的棕色脂肪细胞(图7 F)。这表明miR-433抑制了山羊棕色脂肪细胞的分化和产热。
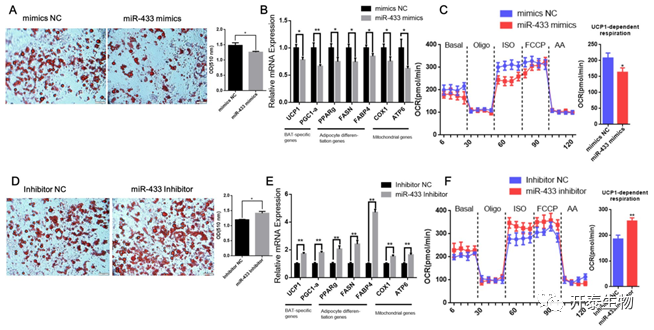
图7 miR-433抑制成熟棕色脂肪细胞分化和产热
07
miR‑433对白色脂肪细胞的分化和产热没有影响
为了研究miR-433在白色脂肪细胞分化中的作用,作者将miR-433模拟物转染到白色脂肪细胞中。结果表明,miR-433模拟物未导致脂质沉积的显著变化(图8A)。脂肪生成标记物和线粒体相关基因的表达无显著变化(图8B)。反之,沉默miR-433并不能促进白色脂肪细胞的分化(图8 C)。脂肪生成标记物和线粒体相关基因的表达也无显著变化(图8D)。这表明miR-433对白色脂肪细胞分化和产热没有影响。
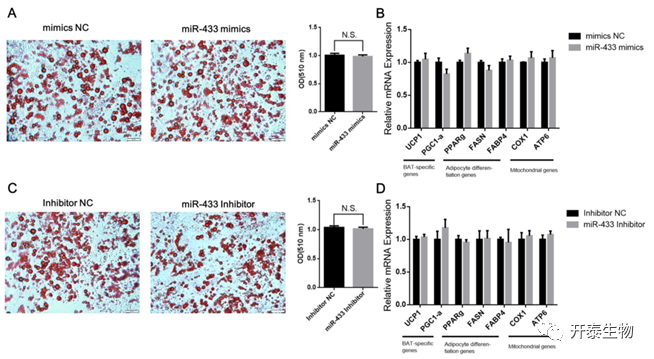
图8 miR‑433对白色脂肪细胞的分化和产热没有影响
08
MAPK8是miR‑433的靶基因
靶基因预测软件在MAPK8基因的3’ UTR内鉴定了一个高度可能的miR-433结合位点,该位点在哺乳动物中高度保守(图9A)。为了验证MAPK8是否是miR-433的直接靶基因,将MAPK8的野生型和突变型3' UTR分别构建到psiCHECK2载体中(图9B)。miR-433模拟物与山羊MAPK8基因野生型3' UTR结合,导致荧光素酶报告基因的活性降低(图9C)。相反, MAPK8 突变型3'UTR无法结合miR-433模拟物,所以荧光素酶活性无显著变化(图9C)。经miR-433模拟物或抑制剂处理的MAPK8基因的表达相应的上调或下调(图9D)。结果表明,miR-433通过靶向MAPK8基因3’ UTR抑制MAPK8的表达。
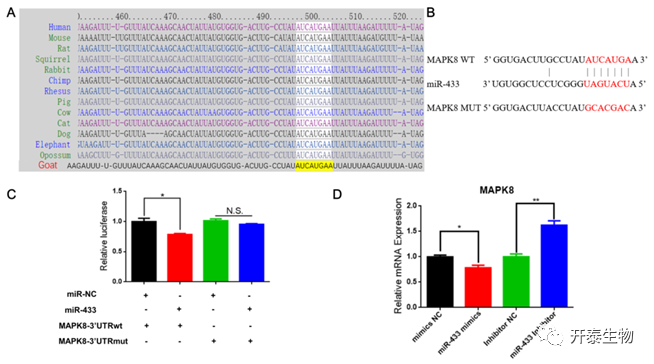
图9 MAPK8是miR‑433的靶基因
+ + + + + + + + + + +
结 论
本研究通过RNA测序比较了BAT和WAT中差异表达的miRNA。对于BAT富集的miRNA,体外细胞实验证实miR-433对棕色脂肪细胞的分化和产热有抑制作用。本研究将为今后研究miRNA调节山羊BAT产热提供参考和依据。
+ + + + +



