

 English
English文献解读|Nature(48.5):多组学分析揭示健康成年人与年龄相关的免疫动态
✦ +
+
论文ID
原名:Multi-omic profiling reveals age-related immune dynamics in healthy adults
译名:多组学分析揭示健康成年人与年龄相关的免疫动态
期刊:Nature
影响因子:48.5
发表时间:2025.10.29
DOI号:10.1038/s41586-025-09686-5
背 景
追踪个体终生健康免疫力的动态变化对于理解感染易感性、疫苗反应以及免疫相关疾病的发生至关重要,这些疾病在衰老过程中表现出不同的发病率。虽然许多关于免疫年龄的研究利用免疫系统的单一快照来推断衰老过程的共同特征,但免疫细胞的功能始终受到时间因素的影响。固有免疫细胞(即单核细胞和中性粒细胞)主要参与快速且随机的反应(持续数小时至数天),而适应性免疫细胞(T细胞和B细胞)则介导较慢但更持久的记忆反应(持续数天至数十年)。事实上,近期一些研究着重于在感染、疫苗接种和体内平衡的背景下进行纵向监测,为大众提供了免疫系统与年龄相关的变化的独特视角,并加深了对人类免疫力随时间动态相互作用的理解。
实验设计
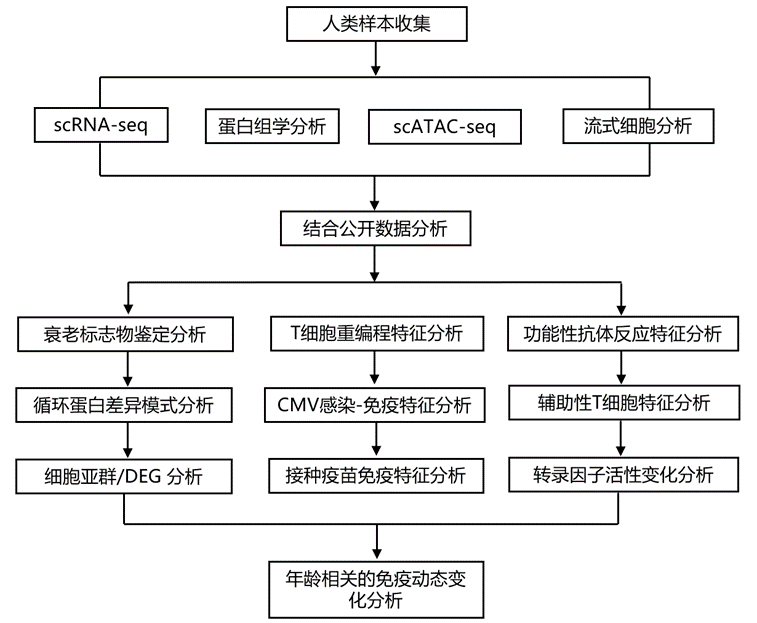
结 果
01

健康人类免疫细胞转录组
关于健康个体免疫反应纵向动态的研究十分有限。为了弥补这一空白,研究团队启动了“健康生活项目”(Sound Life Project),该项目前瞻性地招募了1349名青年人(25-35岁)和47名老年人(55-65岁),并对他们进行了为期两年的随访。在此期间,所有参与者均接种了两次季节性流感疫苗,并采集了多达十次血液样本(图1a)。他们在每个时间点采集了外周血单核细胞(PBMC)和血浆,并利用转录组分析(scRNA-seq)技术、光谱流式细胞技术和血浆蛋白质组学技术进行分析。他们首先分析了循环蛋白质组数据,发现由69种蛋白质标记的细胞亚群的频率随年龄变化而差异表达,包括先前报道的衰老标志物CXCL17、WNT9A和GDF15(图1b-c)。循环蛋白质变化与巨细胞病毒 (CMV) 感染状态[CMV 血清阳性 (CMV+) 或 CMV 血清阴性 (CMV−)]之间无显著相关性,仅在接种流感疫苗后观察到一处变化。因此,未达到老年且无全身炎症的情况下,循环衰老标志物在成年人中持续存在。
接下来,他们利用scRNA-seq技术研究了健康成年人体内免疫细胞的动态变化,构建的纵向数据集包含71个免疫细胞亚群,这些亚群均使用新开发的“人类免疫健康图谱”(Human Immune Health Atlas)进行标记。他们检测了35个T细胞亚群、11个B细胞亚群、7个单核细胞亚群、6个NK细胞亚群以及包括树突状细胞和造血前体细胞在内的12个其他亚群的RNA表达(图1d),发现T细胞的转录水平随年龄增长而发生显著变化(图1e)。在非T细胞亚群中,几乎未观察到与年龄相关的转录变化。分化早期T细胞亚群显示出最多的与年龄相关的差异表达基因(DEG),其中幼稚T细胞的数量最多,其次是记忆T细胞(TCM),最后是效应记忆T细胞(TEM)。一年后,T细胞亚群中与年龄相关的DEG数量仍然相似。此外,T细胞特异性转录变化的模式与性别差异、慢性巨细胞病毒(CMV)感染或急性流感疫苗接种引起的转录变化模式截然不同(图1e)。与年龄相关的转录变化与细胞组成没有直接相关性(图1f)。事实上,核心幼稚CD4 T细胞的转录组变化最为显著,且其频率随年龄增长无显著变化;而核心幼稚CD8 T细胞的转录组变化和频率变化均随年龄增长而变化。因此,T细胞在衰老早期表现出最显著的转录组和组成改变,而其他固有免疫和适应性免疫细胞的基线变化则较少。
稳态衰老过程与转录变异相关;然而,免疫细胞编程如何随年龄变化尚不清楚。为了进一步确定T细胞中与年龄相关的转录变化的稳定性,他们分析了8个差异性免疫细胞亚群在600天内的转录动态:核心幼稚CD8 T细胞;核心幼稚CD4 T细胞;CD8 TCM细胞;GZMK+CD27+CD8 T EM细胞;幼稚CD4调节性T细胞(Treg);CD4 TCM细胞;GZMB− CD27− CD4 T EM细胞;以及GZMB− CD27+CD4 T EM细胞(图1e)。为此,他们开发了RNA年龄指标(RAM),这是一个针对上调(和下调)DEG的亚群特异性RNA综合评分,作为与年龄相关的转录编程的概括性指标。正如预期,在所有亚组中,老年人的RAM值均高于年轻人,且转录重编程发生在个体T细胞群中。在为期两年的随访期间,老年人的RAM值也高于年轻人(图1g)。他们进一步将RAM(作为转录重编程的指标)与两个已确立的衰老参数进行了比较:(1)基于scRNA-seq数据中细胞频率的IMM-AGE;以及(2)基于队列样本近期生成的全血转录组学数据的免疫健康指标。所有三个衰老指标均显著相关,表明免疫衰老发生在细胞内和多细胞水平。此外,这些数据共同表明,T 细胞区室在衰老早期和无炎症的情况下会经历稳定的转录重编程。
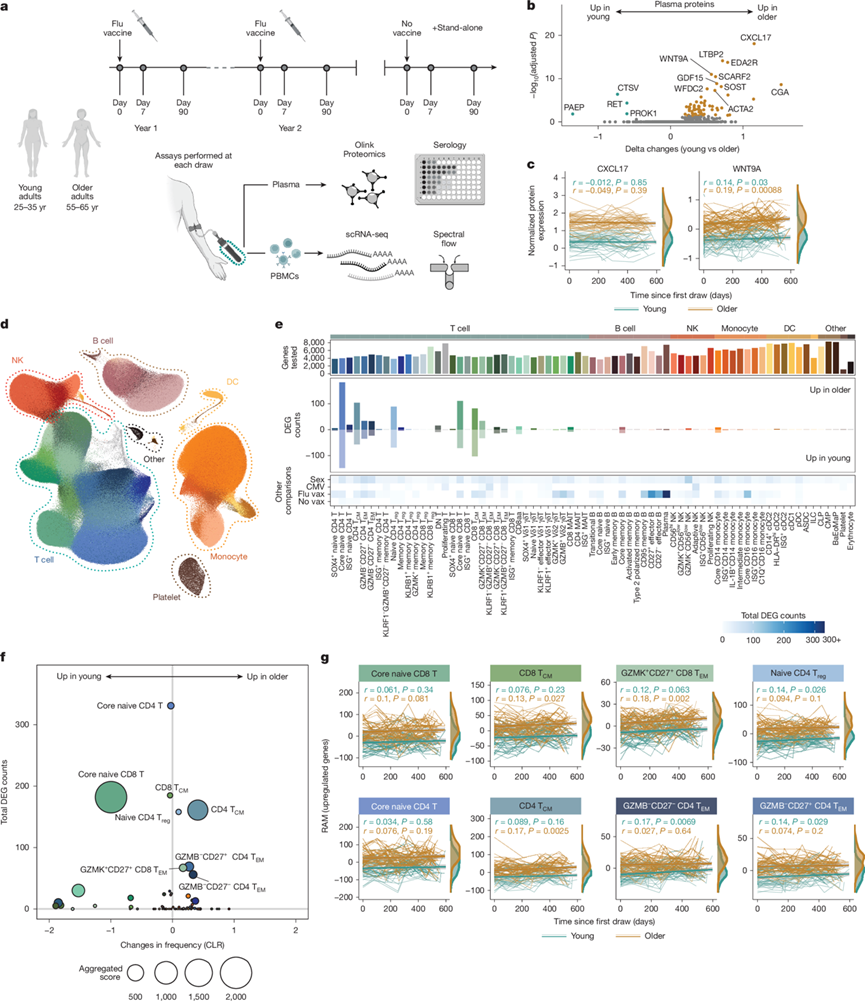
图1. 健康人类免疫细胞转录组随时间推移保持与年龄相关的变化。
(a) 健康青年和老年人的“健康生活”队列概览。(b) 循环血浆蛋白质组中与年龄相关的蛋白质表达差异的火山图。(c) 青年和老年人血浆中 CXCL17 和 WNT9A 蛋白表达随时间变化的标准化值。(d) 对包含1370万个PBMC的纵向人类免疫衰老数据集进行均匀流形逼近和投影(UMAP)分析。(e) 差异表达基因(DEG)分析。(f) 青年人和老年人之间差DEG频率和数量变化的气泡图。(g) 取每个包含超过20个DEG的子集中上调最显著的DEG的综合评分,并根据年轻和老年人群随时间的变化进行分析。
02
各个年龄段常见的T细胞重编程
为了解衰老过程中蛋白质组、组成和转录变化的更广泛动态,他们对第二组健康成年人(年龄跨度从 40岁到90岁以上)进行了免疫谱分析(图2a)。与本研究的纵向队列研究结果一致,蛋白质组分析表明,在健康年龄阶段发生改变的循环蛋白会随着年龄的增长而逐渐积累(例如CXCL17)。虽然IL-6和IL-11与高龄有一定程度的相关性,但他们发现经典炎症标志物TNF和IL-1B与年龄无关(图2b),这表明不同类型的炎症可能导致晚年免疫失调。
利用后续的包含超过320万个PBMC的scRNA-seq数据集,他们初步证实了已知的免疫衰老标志,包括核心幼稚CD8 T细胞的持续减少(图2c-d)和CD27⁻效应B细胞(即“年龄相关B细胞”)的扩增(图2e)。随后,他们使用RAM追踪T细胞重编程,发现8个年龄敏感型T细胞亚群的RAM值随年龄增长呈持续但非线性增加(图2f)。在两个独立的scRNA-seq数据集中也发现了类似的增加,这两个数据集包含超过1000名健康成年人(图2g)。通过评估各亚群中RAM的共有基因,他们鉴定出26个随年龄增长而下调的基因和36个随年龄增长而上调的基因,其中包括与效应细胞分化(如PTGER2)、细胞内信号传导反应(SESN3)、细胞状态极化(IL16)和细胞凋亡(STK17A)相关的基因(图2h)。利用近期发布的人类组织图谱,他们还发现人类淋巴结中多个T细胞亚群的RAM表达随年龄增长而增加(图2i),表明T细胞重编程并非仅限于外周血。此外,RAM可能有助于追踪与疾病发生相关的免疫年龄加速。为了验证这一理论,他们利用RAM分析了一组有罹患年龄相关性自身免疫性类风湿性关节炎风险的青年人(20-40岁)的转录衰老情况。值得注意的是,他们发现高危成年人CD4 T CM细胞中的RAM表达显著高于同龄健康对照组(图2j);已知该T细胞亚群参与临床诊断疾病患者自身抗体的产生。因此,这些分析揭示了T细胞亚群中保守的转录重编程,这种重编程在健康人群中随年龄增长而逐渐发展,并可作为健康和疾病状态下免疫衰老的指标。
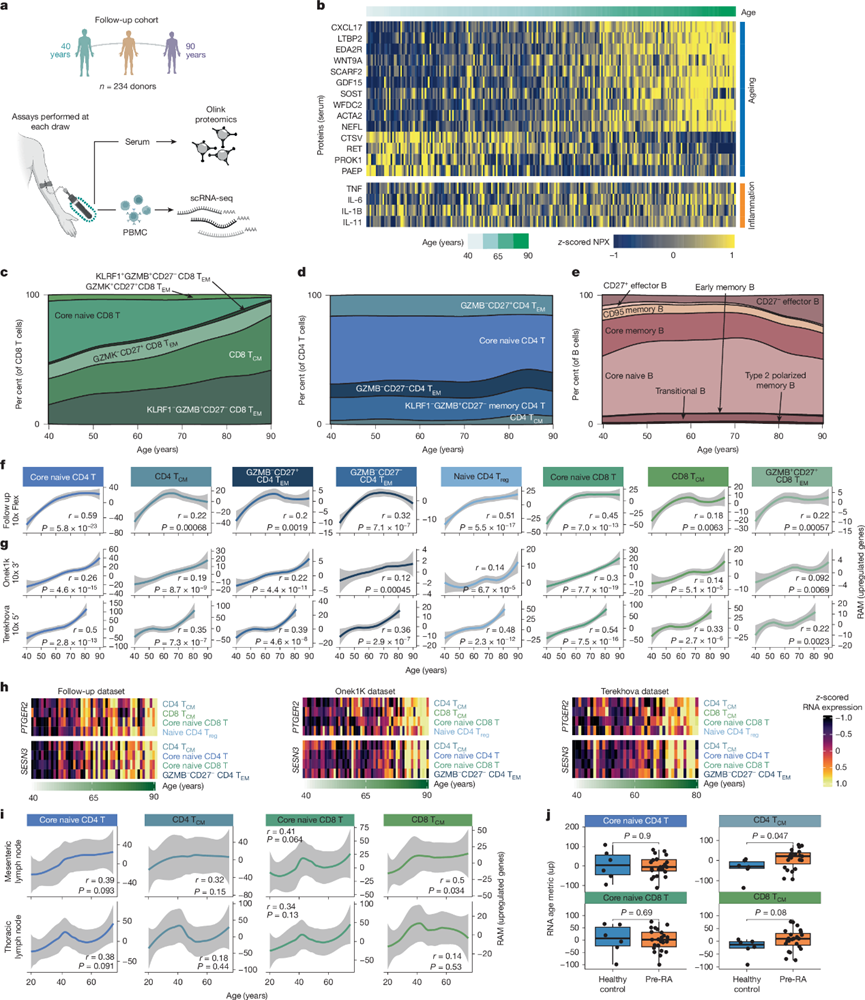
图2. 健康人体免疫景观随年龄变化的动态。
(a) 健康成年人随访队列概览。(b) 与年龄和炎症相关的血清蛋白的标准化蛋白表达(NPX)。(c-e) 来自320万个PBMC随访scRNA- seq数据集的CD8 T细胞、CD4 T细胞和B细胞组分在不同年龄段的组成。(f-g) 随访队列和两个外部免疫衰老数据集中,各年龄段受年龄影响最大的免疫细胞亚群的平均RAM(上调基因)。(h) 三个独立免疫衰老数据集中,从初始DEG分析中鉴定出的部分上调基因的平均RNA表达量随年龄变化的热图。(i) T细胞亚群 在不同年龄段的RAM比较。(j) T细胞亚群的RAM比较。
03
巨细胞病毒感染和年龄的不同影响
CMV是一种慢性病原体,可随时间推移导致低度炎症。然而,在65岁之前,CMV阳性和CMV阴性的健康成年人之间,循环蛋白质或全身炎症水平并无差异。因此,为了更好地评估CMV感染对免疫微环境的影响,他们对纵向队列(CMV阳性:n = 44;CMV阴性: n = 52)和随访队列(CMV阳性:n = 136;CMV阴性:n = 98)中CMV阳性和CMV阴性个体的71种免疫细胞亚群进行了详细比较,发现三种常规 T 细胞亚群(KLRF1+/-GZMB+CD8+ TEM 细胞和 KLRF1-GZMB +CD4+ TEM细胞)、两种非常规 T 细胞亚群(KLRF1+/-效应 Vδ1 γδ T 细胞)和一种NK细胞(适应性 NK 细胞)在 CMV 感染后持续显著增加(图3a)。这些相同的免疫细胞亚群在CMV血清转化后也出现扩增(图3b)。接下来,他们比较了 CMV 阳性个体中这些扩增亚群的年龄相关变化。在基线或 2 年随访期间,CMV 阳性成人中终末样效应 T 细胞亚群(KLRF1+/-GZMB+ CD8+TEM细胞)的频率未检测到年龄相关差异(图3c-d)。随着年龄增长,适应性NK细胞显著减少,且该减少趋势持续两年(图3e-f)。CMV阳性老年个体的适应性NK细胞中,谱系标志物KLRC2的表达也呈下降趋势(图3g),表明细胞内程序可能发生改变。值得注意的是,虽然CMV感染本身会改变应答亚群的转录组,但他们几乎没有发现这些亚群中存在与年龄相关的转录差异(图3h)。CMV感染也不会改变易感年龄的T细胞亚群中的RAM(图3i),表明慢性CMV感染不会加速T细胞的转录衰老。与此一致的是,他们未检测到衰老或增殖通路基因集富集随年龄增长而发生改变。因此,CMV会以一种能够长期维持的方式重塑外周免疫系统,但CMV感染引起的转录重编程与健康衰老不同。
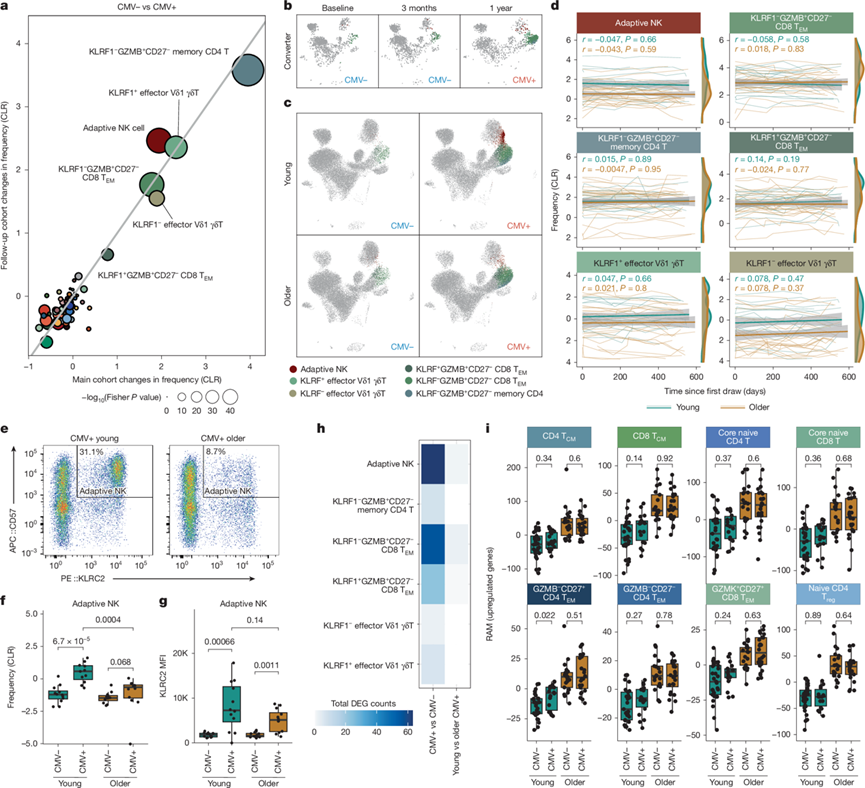
图3. CMV 感染和年龄对周围免疫细胞格局的影响。
(a) 气泡图比较了 主要队列中流感疫苗接种第一年第0天CMV阳性成人和CMV阴性成人以及 随访队列中CMV阳性成人和CMV阴性成人细胞类型频率的变化。(b-c) 分别展示了一名年轻成人CMV血清转换者和CMV阳性及CMV阴性年轻成人及老年人的T细胞和NK细胞亚群的UMAP图。(d) 展示了年轻和老年人外周血单核细胞(PBMC)中特定亚群频率随时间的变化。(e-g)流式细胞分析。(h) 热图显示了CMV阳性组与CMV阴性组以及年轻CMV阳性成人与年长CMV阳性成人 之间DEG的平均数量。(i) 根据年龄和巨细胞病毒 (CMV) 感染状态划分的年龄易感 T 细胞亚群中的 RAM(上调基因)。
04
随着年龄增长,B细胞对疫苗接种的反应会发生变化
众所周知,年龄会影响个体对疫苗产生有效且持久抗体反应的能力,但对于健康老龄化过程中功能性B细胞反应的了解却相对较少。为了研究这一问题,他们选取了43名青年人和45名老年人,在样本采集的第一年接种了相同的2020-2021年度灭活、无佐剂四价流感疫苗(图4a)。在接种疫苗后的第0、7和90天,使用针对多种流感疫苗株定制的血凝抑制(HAI)试验,检测了流感特异性免疫球蛋白G(IgG)抗体水平及其功能特性。除流感B/Phuket毒株(自2015年起成为流感疫苗的成分之一)外,年轻成人和老年人对大多数流感毒株的血凝抑制(HAI)水平差异甚微。老年人对流感B/Phuket毒株的HAI水平显著低于年轻成人,无论是在基线水平还是接种疫苗后7天。为了解独立于基线水平的免疫应答差异,他们通过线性模型评估了每种疫苗毒株的“应答”(第0天与第7天)和“持久性”(第7天与第90天),并考虑了性别、CMV感染和基线抑制水平等因素。虽然不同人群对多种流感病毒的反应存在异质性(图4b),但他们观察到老年人群中对 B/Phuket 流感病毒的高反应者数量较少,且这一现象在不同流感季均保持一致(图4c),这凸显了老年人群中较低的基线抑制水平和较少的高反应者(图4c-d)。他们未发现 HAI 反应的持久性存在年龄相关差异。这些数据表明,尽管大多数流感疫苗反应仍然有效,但在老年之前,对反复加强免疫的流感抗原的功能性抗体反应强度会降低。
此前观察到,接种疫苗后记忆B细胞亚群的转录水平发生改变(图1e);因此,他们接下来研究了流感疫苗诱导的B细胞反应随年龄的变化。在年轻成人和老年人中,接种疫苗后浆细胞、CD27-效应记忆细胞和CD95+记忆B细胞的频率均显著增加,而核心记忆细胞和2型极化记忆B细胞亚群在老年人中的扩增则有所减少(图4e)。他们通过流式细胞分析证实了这些观察结果,结果与RNA测序检测到的B细胞频率高度相关。总的来说,疫苗相关的浆细胞或CD27-效应记忆B细胞的频率随年龄变化不大。
接下来,他们分析了接种疫苗后第7天的转录组谱。浆细胞的通路富集表明其活化程度更高,而核心记忆B细胞在老年人中MYC活化程度较低,这与其扩增受限相符。相比之下,CD27-效应记忆B细胞在老年人中活性氧信号持续降低,表明其对流感疫苗的反应性降低。老年人与功能性效应记忆B细胞相关的基因表达也较低(图4f-g),包括谱系定义基因(例如FCRL5、CD19和MAS4A1)和活化基因(ZEB2、TBX21和BATF)。老年人中未发生类别转换的免疫球蛋白基因IHGM和IGHD的表达增加,提示B细胞亲和力成熟度降低。事实上,流感疫苗有效B细胞应答的关键特征是IgG类别转换,从而增加IgG的产生。值得注意的是,老年人CD27-效应记忆B细胞在第7天时IgG基因表达持续降低(图4h),突显了该记忆亚群的类别转换随年龄增长而发生改变。采用基于流式细胞分析的B细胞注释策略,证实了老年人接种疫苗后IgG+CD27 -效应记忆B细胞频率降低(图4i)。该亚群在老年人中CD19蛋白表达也较低,与其年龄相关的基因表达谱相符。因此,CD27 −效应记忆B细胞是流感疫苗抗体反应发展和维持中的一个重要亚群,随着年龄的增长,其转录组发生改变,IgG减少,这与老年人中B/Phuket特异性IgG较低相一致(图4j)。
流感疫苗接种通常会诱导强烈的IgG1和IgG3反应。值得注意的是,CD27−效应记忆B细胞亚群的IgG同型表达发生了改变,老年人中IGHG2的表达高于IGHG1、IGHG3和IGHG4(图4h)。因此,他们假设流感特异性IgG同型反应可能随年龄而改变。对流感毒株特异性IgG2和IgG3水平的评估显示,接种疫苗后所有毒株的IgG3水平均升高,而IgG2水平仅在B/Phuket毒株中升高。这种B/Phuket血凝素(HA)特异性IgG2的升高在青年人和老年人中均有观察到,但在老年人中更为显著。事实上,他们观察到老年人群中IgG2/IgG3比值存在显著偏斜,且第7天老年人群中B/Phuket特异性IgG2的相对含量增加(图4k)。在另一项独立数据集中,他们也发现老年人(65岁以上)流感特异性核心记忆B细胞中IGHG2/IGHG3 RNA比值呈上升趋势。IgG类别转换需要T细胞的辅助。与此一致的是,流感特异性IgG2/IgG3偏斜、效应记忆B细胞活化和HAI均与记忆T细胞的转录重编程相关(图4l)。这表明,记忆T细胞重编程的年龄相关性变化可能导致老年人对季节性流感疫苗接种的B细胞反应发生改变。
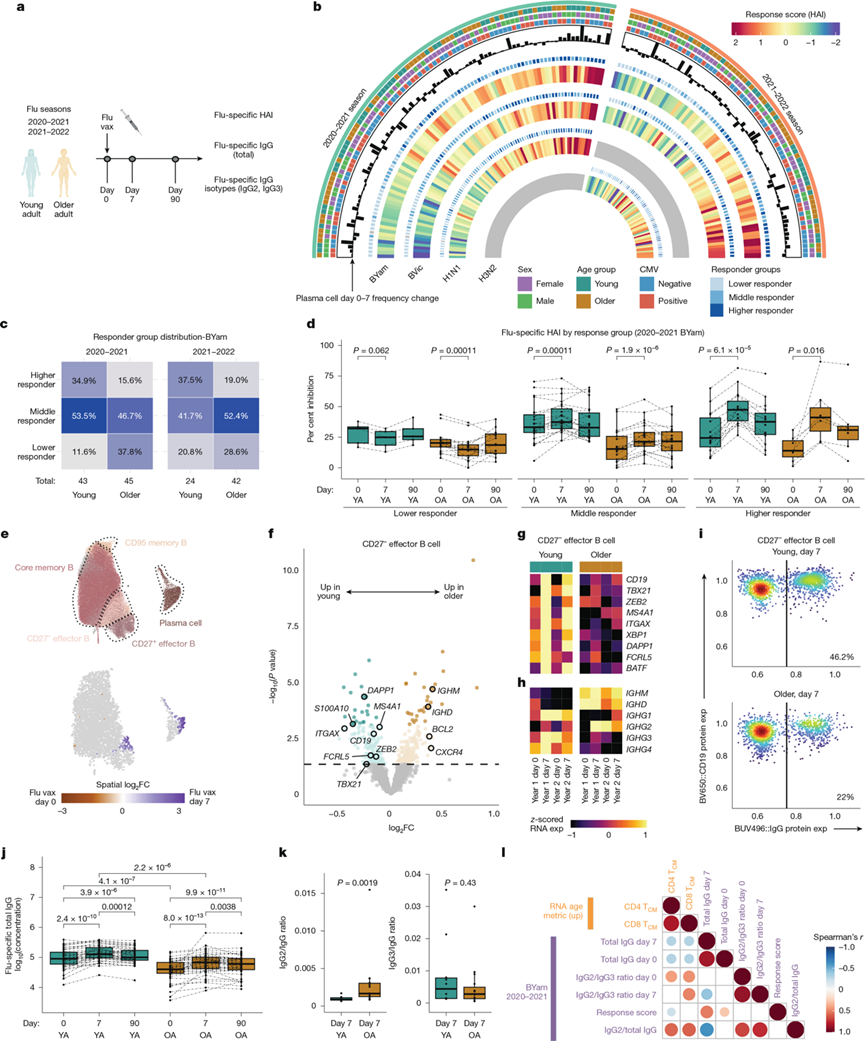
图4. 与年龄相关的 B 细胞对流感疫苗的反应。
(a) 流感疫苗接种队列和研究设计概述。(b) 2020-2021和 2021-2022 流感季接种疫苗后第 0 天和第 7 天的 HAI 反应 Circos 图。(c) BYam病毒株(B/Phuket) 的 HAI 反应按年龄分组分布。(d) HAI 平均抑制率。(e) 使用Milo对接种疫苗前和接种疫苗后第7天记忆B细胞亚群及其差异丰度进行UMAP分析。(f) CD27-效应记忆B细胞DEG 的火山图。(g-h) CD27-效应记忆B细胞中激活基因和免疫球蛋白基因的纵向表达。(i)流式细胞分析。(j) 血浆中B/Phuket特异性IgG水平。(k) 血浆中B/Phuket特异性IgG2和IgG3水平。(l) 比较流感特异性反应与TCM的RAM的Spearman相关性。
05
TH2 偏向和 B 细胞反应随年龄变化
体外人体研究表明,IgG2和IgG3 类别转换的偏倚与 IL-4 对 B 细胞亚群的刺激有关。因此,他们接下来研究了 T 细胞群中的辅助性T细胞 1 (TH1)样、TH2样和 辅助性T细胞 17 (TH17) 样状态随年龄的变化。虽然他们观察到循环中辅助性T细胞相关细胞因子水平并未随年龄增长而增加,但与年轻人相比,老年人CD4+TCM细胞中TH2样状态显著偏倚(图5a)。这种TH2样状态的增加在老年人中持续存在(图5b)。此外,他们发现 CD4+TCM细胞中TH2样状态持续增加,直至老年(图5c)。TH 1样状态和TH 17 样状态均未随年龄或时间发生显著变化。为了验证年龄与TH2 样状态发展之间的关联,他们利用三模态 TEA-seq 数据集(整合了scRNA-seq、表位和染色质可及性数据)同时检测了转录因子活性和 CD4 T 细胞状态。使用基于 RNA 的伪时间方法,根据转录相似性对 CD4 T 细胞进行排序(图5d),并分析了相应的单细胞转座酶可及染色质测序 (scATAC-seq) 数据,以检测开放染色质区域中转录因子基序的富集情况,并将其与预测的转录因子活性进行比较。GATA3是一种重要的TH2相关转录因子,其活性在记忆性CD4 T细胞群中随年龄增长显著升高,包括CXCR5+和CXCR5−CD4 TCM细胞(图5e)。事实上,GATA3活性与CD4 TCM细胞中TH2样转录状态呈正相关。此外,其他TH2相关转录因子,包括STAT6、STAT5A和IRF4的活性也随GATA3的增加而升高。主要的TH1相关转录因子TBX21的活性随年龄增长没有显著变化。值得注意的是,记忆性CD8 T细胞亚群也表现出GATA3活性随年龄增长而升高,表明广泛的TH1相关转录因子参与了记忆性CD8 T细胞群的形成。
传统上,TH2细胞偏向与IL-4分泌倾向增强相关。与较高的GATA3活性和TH2样转录状态一致,老年人CD4 TCM细胞中IL4基因座的染色质开放度增加(图5f),且TCR激活后T细胞产生的IL-4显著增多(图5g)。TCR刺激后,老年人IFNγ+IL-4+CD27+记忆性CD8 T细胞亚群数量增加,但差异不显著,且与接种疫苗后第7天B/Phuket特异性IgG3水平升高相关。此外,老年人中多种记忆性CD4和CD8 T细胞亚群的自发性IL-4产生显著升高,这与轨迹分析结果一致,表明多种T细胞亚群存在TH2样状态倾向(图5h-i)。自发性IFNγ− IL-4+ CD4 T CM细胞的频率与CD4 TCM细胞的RAM和IL-4产生呈显著正相关(图5j),表明CD4 TCM细胞的转录状态与其下游功能之间存在直接联系。此外,接种疫苗后第7天B/Phuket特异性IgG2水平与IL-4产生呈正相关。总的来说,成年人的记忆T细胞在老年之前会逐渐向 T H 2 样状态转变,并且这种重编程与记忆 B 细胞活化减少、类别转换改变以及随着年龄增长和反复接触抗原而导致的抗体功能减弱有关。
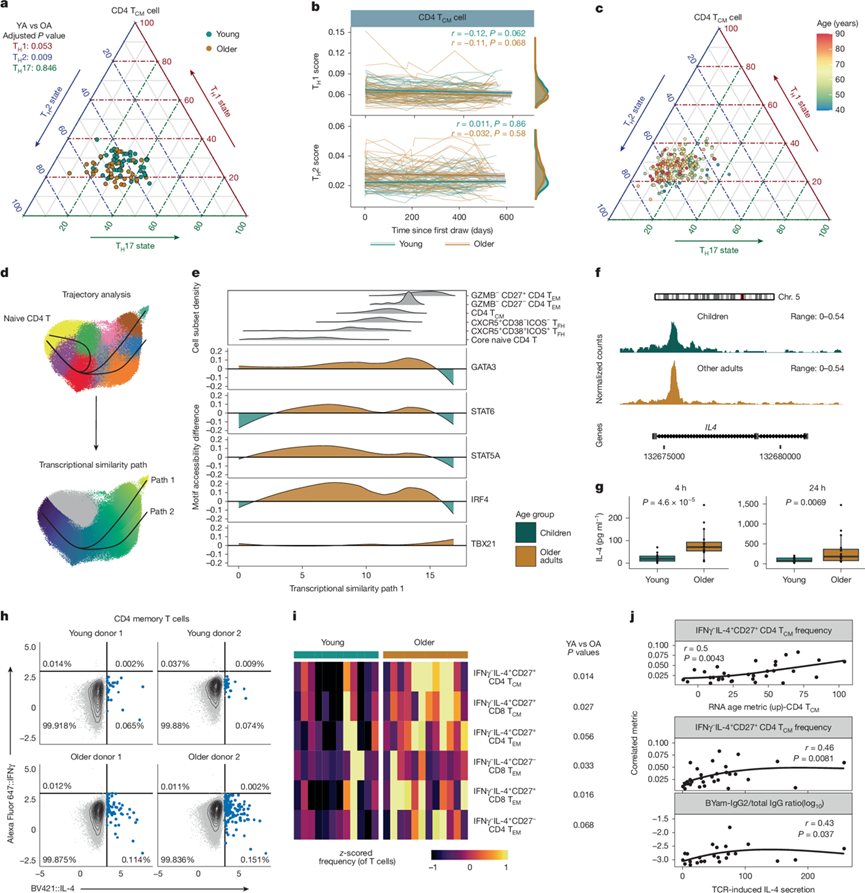
图5. 随着年龄增长,记忆 T 细胞中 TH2 样状态发生改变并逐渐积累。
(a) 年轻成人和老年人CD4 TCM细胞中 TH1、TH 2和 TH 17 状态评分的三角形图。(b) 年轻成人和老年人CD4 TCM细胞中TH1 和 TH2 状态评分随时间的变化。(c) CD4 TCM细胞中TH1、TH2 和 TH17 状态评分的三角形图。(d) 基于 T 细胞 TEA-seq 数据集的 CD4 T 细胞 RNA 轨迹分析。(e) 基于Chromvar分析,对来自儿童和老年人CD4 T细胞的scATAC-seq数据进行分析。(f) 染色质可及性轨迹。(g) T细胞分泌的IL-4浓度。(h)流式细胞分析。(i) IFNγ- IL-4+记忆性T细胞频率的热图。(j) IFNγ-IL-4+ CD4 T CM细胞与CD4 TCM细胞中的RAM以及TCR刺激后IL-4分泌与IFNγ-IL-4+ CD4 TCM细胞频率的Spearman相关性。
+ + + + + + + + + + +
结 论
本研究利用scRNA-seq、蛋白质组学和流式细胞技术,对300多名健康成年人(25至90岁)的外周免疫进行了分析。其中96名成年人接受了为期两年的季节性流感疫苗接种随访。由此,构建了一个包含超过1600万个外周血单核细胞的scRNA-seq据集,其中包含来自我们人类免疫健康图谱的71个免疫细胞亚群。该数据集使后续相关研究人员能够探究免疫细胞组成和状态如何随年龄、慢性病毒感染和疫苗接种而变化。基于这些数据,本研究证实了T细胞亚群中存在显著的非线性转录重编程,且该过程并非由全身炎症或慢性巨细胞病毒感染驱动。这种与年龄相关的重编程导致记忆性T细胞中功能性TH2细胞偏向,这与流感疫苗中高浓度增强抗原引起的B细胞反应失调有关。总的来说,这项研究揭示了高龄之前发生的免疫衰老过程的独特特征,并为与年龄相关的免疫调节提供了新的靶点。
+ + + + +



